黄野螟的羽化和生殖行为节律
2018-06-29王忠谢伟忠朱诚棋陆雪雷曹春雷温秀军
王忠,谢伟忠,朱诚棋,陆雪雷,曹春雷,温秀军
(1.广东省林业有害生物防治检疫管理办公室,广东 广州 510173; 2.华南农业大学林学院林学与风景园林学院,广东 广州 510642)
土沉香Aquilariasinensis(Lour.) Gilg为瑞香科Thymelaeaceae沉香属Aquilaria植物,又名白木香、女儿香、崖香和蜜香等。主产于海南、广东、广西、福建、台湾等低纬度地区[1]。土沉香不仅是传统名贵的天然香料[2],还是唯一能够生产中药沉香的药用植物[3],现代医学研究表明,中药沉香具有温和止呕,行气止痛,纳气平喘的功效,可用于治疗胸腹、胀闷疼痛,肾虚气逆喘急等疾病。
黄野螟HeortiavitessoidesMoore属鳞翅目Lepidoptera草螟科Crambidae齿螟亚科Odontiinae,国内主要分布于广东、云南、福建、广西、海南、台湾、香港等地,国外主要分布于印度、尼泊尔、斯里兰卡、斐济等国家以及东南亚、东印度群岛及新赫布里底群岛等地[4],是土沉香的主要害虫。幼虫蚕食土沉香叶片导致叶柄脱落,暴发时将受害区内的土沉香全部吃光,造成树势下降甚至死亡,严重影响土沉香正常生长[5]。
昆虫性信息素用于害虫的大量诱捕、监测和干扰交配[6],目前已经在多个国家和地区推行。目前对于黄野螟成虫生殖行为学的研究较少,仅对其危害情况、生物特征有一些报道[7-8]。本文通过对黄野螟羽化、性行为和寿命进行研究,以期为该虫性信息素的提取和鉴定提供科学依据,更好地服务于该虫种群监测和防控工作。
1 材料与方法
1.1 供试虫源 黄野螟老熟幼虫采自广州市白云区太和镇和广州天麓湖森林公园,将幼虫分装入养虫箱(30 cm×40 cm×60 cm),移入暗室(温度25±1 ℃,光周期L∶D =14 h∶10 h,光期06∶00—20∶00,相对湿度RH=75%~80%),观察化蛹及羽化情况。成虫羽化后0~24 h视为1日龄,>24~48 h为2日龄,依此类推。
1.2 试验方法
1.2.1 黄野螟羽化节律观察 黄野螟老熟幼虫化蛹后,每天定时观察蛹的羽化状况。当第一头蛹开始羽化,则全天24 h观察黄野螟蛹的羽化状况,每隔2 h记录1次羽化的雌雄虫数,直到所有蛹全部羽化。观察羽化过程及羽化时间,计算成虫的雌雄比例。用加权平均法计算雌雄蛹羽化的时间差[9]:
M差异(d)=雌虫羽化M♀-雄虫羽化M♂
M=(X1×1+X2×2+…+Xn×n)/(X1+X2+…+Xn)
Xn为第n d(自羽化初日算起)的羽化数量。
1.2.2 黄野螟生殖行为观察 将同日羽化的雌雄成虫放入密闭养虫笼(高13 cm,上口直径17 cm,下口直径12 cm),笼内放入土沉香枝条以及蘸有5 %蜂蜜水的棉球,每日更换棉球,全天24 h每隔30 min观察求偶行为,记录不同日龄雌虫求偶起始时间、求偶持续时间及求偶次数。求偶行为及求偶次数以雌虫腹部末端伸出性腺体为标准,全天观察黄野螟的交尾行为,记录交尾的起始时间和持续时间,数据分析时按照每小时的交尾率进行统计,设3组重复,共观察600对。
观察交尾次数。将交尾后的雌、雄成虫取出,分别放入已有5头未交配健康异性的密闭养虫笼中,全天观察并记录是否有交尾行为及交尾次数。实验重复10次。
1.2.3 黄野螟产卵行为 将交尾后的雌蛾置于稍大的养虫笼(高30 cm,直径20 cm)中,笼内放入新鲜的土沉香枝条,并以5 %的蜂蜜水喂养,逐日更换新鲜蜂蜜水棉球,暗期中每隔30 min观察雌虫的产卵情况,并记录雌蛾的产卵方式、产卵时间及产卵量。
1.2.4 黄野螟的成虫寿命 将同日羽化的成虫30头分别置于3个养虫笼(60 cm×60 cm×70 cm)中,笼内各放入一棵土沉香幼苗,每日更换5%的蜂蜜水棉球,设置3个处理:未交尾的雄虫,未交尾的雌虫,雌雄成虫混合。每日暗期结束后观察3个养虫笼中黄野螟的成活情况。
1.3 数据统计方法 采用Microsoft Excel 2013软件处理数据并制图,SPSS 22统计软件对数据进行分析处理,用Duncan氏新复极差法检验差异性。
2 结果与分析
2.1 成虫的羽化节律
2.2.1 成虫的羽化行为 黄野螟老熟幼虫初化蛹时,蛹体呈淡黄色,具有光泽,在羽化前,蛹体颜色变深,渐变为深褐色。羽化时,蛹体不断扭动腹部,并用头部顶破蛹壳,约3~4 min成虫蜕出蛹壳,初羽化的成虫翅形皱缩,潮湿柔软,静止片刻后四处爬行,并寻找较高处静伏以利翅的伸展,2~3 min后,翅逐渐伸展,双翅合拢成屋脊状立于背上,再经过大约5 min,前后翅完全合拢,则羽化完成。
2.2.2 成虫羽化的时辰节律 从全天24 h的羽化节律来看,羽化行为全天可见,但羽化行为集中出现在20∶00~22∶00,即进入暗期后的2 h,此时段的羽化数占全天羽化的22.6 %。雌雄蛹羽化的高峰期同时出现(图1)。
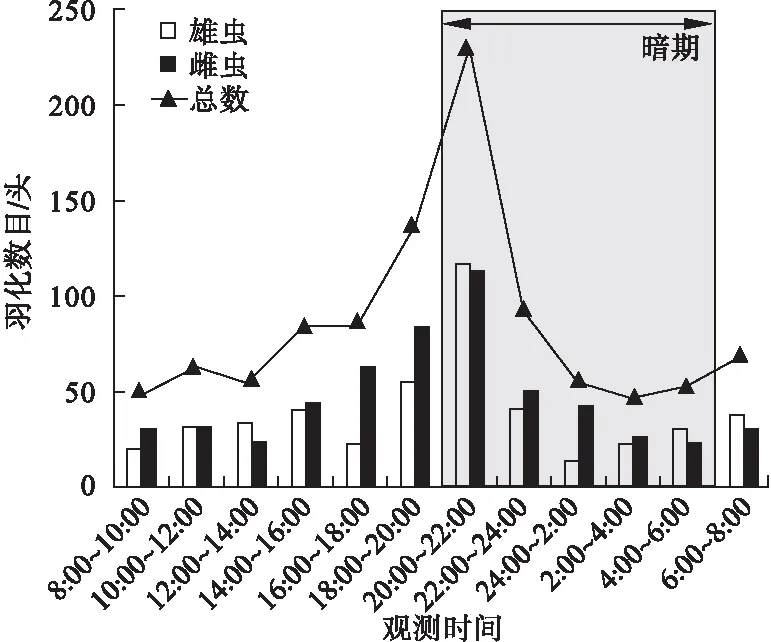
图1 黄野螟的时羽化节律
2.2.3 成虫的日羽化节律 黄野螟的羽化期约为10 d,共观察蛹数1 255头,累计羽化成虫1 044头,羽化率为83.2 %。其中雌虫553头,雄虫491头,雌雄比为1.12∶1,实验室内黄野螟的羽化进度呈现明显的单峰型,这与野外调查时成虫密集羽化状况相同,开始羽化第15~16 d羽化数量较大,雄蛾先于雌蛾羽化,通过公式计算出,雄蛾比雌蛾早羽化0.82 d(图2)。

图2 雌雄成虫性比和羽化时间差异(2014年)
2.3 成虫生殖行为及交尾节律 雌虫的求偶仅发现在暗期,求偶行为非持续,求偶时间约为20 min,求偶时,雌虫会在笼壁上静伏,两翅轻微振动,触角前伸;稍后,雌虫的腹部末端轻微抬起,使产卵器和性腺暴露于空气中,受召唤的雄蛾会飞向雌蛾,并在雌蛾周围爬行,雄蛾处于高度兴奋状态,触角振动频率升高,张开抱握器试图与雌虫交尾,雌雄交尾姿势呈“一”字形。雌雄成虫交尾时静止不动,当交尾结束,雄虫飞离交尾处,雌虫活跃性也降低(图3)。试验中未观察到雌雄成虫存在多次交尾现象。

图3 黄野螟成虫交尾
对实验室600头成熟处女雌蛾的求偶现象观察发现,黄野螟雌成虫羽化次日至第5天均有求偶行为,但在第6天没有观察到其求偶行为。这表明黄野螟雌成虫羽化次日生殖器官已发育成熟。雌虫在2日龄与3日龄时出现求偶行为高峰,求偶行为占整个观察过程中出现次数的60.8 %,其中2日龄占33.3%,3日龄占27.5%(图4)。室内实验还观察到日龄较大(5~6 d)的雌蛾求偶时间提前。

图4 不同日龄黄野螟雌成虫的求偶次数
处女雌蛾在进入暗期30 min始至暗期结束,均有求偶行为出现,其求偶行为多发生在20∶00~21∶00,显著大于暗期其他时间(P<0.05)。在此时间内出现的求偶次数占成虫期总求偶行为次数的60.2%,之后求偶行为逐渐减少(图5)。
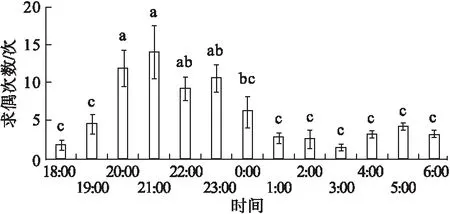
图5 黄野螟雌成虫不同时间的求偶规律
2.5 黄野螟的成虫寿命 成虫的寿命与其交尾情况相关(表1),在未交尾情况下,雌虫平均寿命为(8.83 ± 0.64)d,最长寿命为11 d,雄虫平均寿命为(8.24±0.76)d,最长寿命为9 d;交尾后雌虫平均寿命为(6.32±0.55)d,最长寿命降低至8 d,雄蛾平均寿命为(5.75±0.26)d,最长寿命降低至7 d。由此可见,未交尾雌蛾寿命长于雄蛾,且未交尾雌雄成虫的寿命显著长于交尾过后的成虫(P<0.05);雌虫寿命长于雄虫,但差异不显著(P>0.05)。

表1 不同处理成虫的寿命
注:表中数据为平均值±标准误,同一列中小写字母代表差异显著(P<0.05),大写字母代表差异极显著(P<0.01)。
2.6 黄野螟成虫产卵量和幼虫孵化率 雌虫交尾结束第2天开始产卵,产卵行为发生在夜间,持续约20 min。产卵时,雌蛾静止在叶片背面,腹部弯曲,伸出产卵器寻找合适的位置产卵,卵为聚产,雌虫平均产1~2块,产卵期间雌虫需要补充营养。黄野螟的卵产于土沉香叶片背面靠近叶尖部分,卵块有重叠,直径为2.38±0.56 mm,初产的卵块为淡黄色,扁圆形,卵粒粘结呈鳞片状,近孵化时呈黑褐色。室内观察卵的孵化率为94.35 %±3.02 %,孵化时间多发生在进入暗期的2 h以后。

图6 黄野螟的卵块和幼虫
3 讨论
黄野螟羽化行为呈现明显的单峰型,即羽化比较集中,这有利于对成虫开展性信息素防治,且化蛹时间较为一致,化蛹场所多于树下泥土中,便于深翻集中清理。该虫羽化和生殖行为受昼夜节律的影响,其羽化、求偶、交尾行为具有明显的阶段性,这与许多螟蛾科的害虫具有相似性,如油松球果螟DioryctriamendacellaStaudinger[10]、印度谷螟PlodiainterpunctellaHübner[11]、豆野螟MarucavitrataFabricius[12]、二化螟ChilosuppressalisWalker[13]等。
同时笔者观察到在黄野螟种群中,雄蛾的羽化早于雌蛾0.82 d,在茶毛虫Euproctispseudoconspersa[14]、六星黑点豹蠹蛾Zeuzeraleuconotum[15]和栎黄枯叶蛾TrabalavishnougigantinaYang[16]均有类似的报道,利用性信息素进行诱杀成虫主要针对雄蛾,利用黄野螟中雄性成虫先羽化的规律,可以对雄蛾进行大量诱杀,降低雌雄成虫的交尾率,达到控制其种群数量的目的。
此外,实验中还观察到日龄较大(5~6 d)的雌虫求偶相比日龄较小(2~ 4 d)的雌虫求偶起始时间提前,这种雌蛾的行为同样表现在烟夜蛾helicoverpaassulta[17]、小地老虎Agrotisypsilon[18]和芦笋木蠹蛾IsocerassibiricaAlpheraky[19]中。推测日龄较大雌虫提早求偶具有一定好处,可能避免与日龄较小的雌虫产生交尾竞争,同时延长暗期交尾时间,提高交配的成功率[20]。
昆虫的性行为反作用于昆虫的寿命。Callado-Galindo指出,雌虫能够通过性行为从雄虫体内获得营养物质(精液、附腺物质等),从而延长雌虫的寿命[21]。这些雄虫生殖腺体的分泌物能够刺激雄虫生殖细胞的成熟以及帮助雌虫的生殖行为[22]。本实验中交尾后的雌雄成虫寿命显著小于未交尾的雌雄成虫(P<0.05),这并不符合Callado-Galindo提出的观点,说明黄野螟雌雄成虫活动主要是完成生殖寿命,且完成生殖行为时消耗了大量能量[23],这种交尾使寿命变短的现象出现在竹笋基夜蛾Kumasiakumaso[24]和褛裳夜蛾Catocalaremissa[25]中。
在此次实验中,室外观察和室内研究结果基本一致,但室内暗室实验中雌雄成虫交尾率不高,可能由于室内温度、湿度以及光周期条件不适宜,养虫盒空间小等原因,需进一步进行实验验证。
参考文献:
[1] 李林海,寿海洋,马清温.土沉香(瑞香科)的地理分布研究[J].安徽农业科学,2012,40(17):9254-9256.
[2] BARDEN A,ANAK N.A,MULLIKEN T.,SONG M.Heart of the matter:agarwood use and trade and CITIS implementation forAquilariamalaccensis[M].Cambridge:Traffic International,2000:24-26.
[3] 梅全喜,李汉超,汪科元,等.南药沉香的药用历史与产地考证[J].今日药学,2011,21(1):3-5.
[4] KALITA J.HeortiavitessoidesMoore (Lepidoptera:Pyralidae) -A serious pest of agarwood plant (AquilariamalaccensisLamk.) [J].Geobios,2002,29(1):13-16.
[5] 陈志云,李东文,王玲,等.土沉香黄野螟生物学特性研究[J].中国植保导刊,2011,31(11):10-14.
[6] WILLSON H R,SEMEL M,TEBCHERANY M,et al.Evaluation of sex attractant and blacklight traps for monitoring black cutworm and variegated cutworm [J].Econ.Entomol,1981,74(5):517-519.
[7] 苏跃平.白木香黄野螟生物学特性[J].中药材,1994(12):7-9.
[8] 乔海莉,陆鹏飞,陈君,等.黄野螟生物学特性及发生规律研究[J].应用昆虫学报,2013,50(5):1244-1252.
[9] 王永模,戈峰,刘向辉,等.茶毛虫成虫的行为习性观察[J].应用昆虫学报,2003,40(4):343-345.
[10]辛海萍,张金桐,宗世祥,等.油松球果螟羽化节律和成虫生殖行为观察[J].山西农业大学学报(自然科学版),2012,32(1):12-17.
[11]NORDLUND D A,EUGENEBRADY U.The calling behavior of femalePlodiainterpunctellaHübner (Lepidoptera:Pyralidae) under two light regimes [J].Environmental Entomology,1974,3(5):793-796.
[12]LU P,QIAO H,WANG X,et al.Adult behavior and circadian rhythm of sex pheromone production and release of the legume pod borer,MarucavitrataFabricius (Lepidoptera:Pyralidae)[J].Acta Entomologica Sinica,2007,50(4):335-342.
[13]KANNO H.Mating behaviour of the rice stem borer moth,ChilosuppressalisWalker (Lepidoptera:Pyralidae):V.Critical illumination intensity for female calling and male sexual response under various temperatures [J].Applied Entomology & Zoology,1981,16:179-185.
[14]王永模,戈峰,刘向辉,等.茶毛虫成虫的行为习性观察[J].应用昆虫学报,2003,40(4):343-345.
[15]刘金龙,宗世祥,张金桐,等.六星黑点豹蠹蛾成虫生殖行为特征与性趋向[J].生态学报,2011,31(17):4919-4927.
[16]刘永华,阎雄飞,章一巧,等.栎黄枯叶蛾羽化及生殖行为研究[J].应用昆虫学报,2013,50(5):1253-1259.
[17]KAMIMURA M,Tatsuki S.Diel rhythms of calling behavior and pheromone production of oriental tobacco budworm moth,Helicoverpaassulta,(Lepidoptera:Noctuidae) [J].Journal of Chemical Ecology,1993,19(12):2953-2963.
[18]XIANG Y Y,Yang M F,Li Z Z.Calling behavior and rhythms of sex pheromone production in the black cutworm moth in China [J].Journal of Insect Behavior,2010,23(1):35-44.
[19]LIU H,ZHAO W,YANG M,et al.Diel rhythms of sexual behavior and pheromone titers inIsocerassibiricaAlpheraky (Lepidoptera:Cossidae) [J].Archives of Insect Biochemistry & Physiology,2013,84(1):15-26.
[20]DELISLE J.Effect of male and female age on the mating success of the obliquebanded leafrollerChoristoneurarosaceana(Lepidoptera:Tortricidae) under different ecological conditions [J].Journal of Insect Behavior,1995,8(6):781-799.
[21]GILLOTT C.Male accessory gland secretions:modulators of female reproductive physiology and behavior [J].Annual Review of Entomology,2003,48(48):163-184.
[22]XU J,WANG Q.Seminal fluid reduces female longevity and stimulates egg production and sperm trigger oviposition in a moth [J].Journal of Insect Physiology,2011,57(3):385-390.
[23]向玉勇,杨茂发,李子忠.交配对小地老虎成虫寿命和繁殖的影响[J].四川动物,2010,29(1):85-86.
[24]舒金平,滕莹,张爱良,等.竹笋基夜蛾的求偶及交配行为[J].应用生态学报,2012,23(12):3421-3428.
[25]郑海霞,阎毅,张金桐,等.褛裳夜蛾成虫生物学与性趋向[J].林业科学,2014,50(12):87-93.
