国外引进甘蔗栽培品种的光合气体交换参数遗传差异与聚类分析
2018-06-29李纯佳覃伟徐超华刘洪博毛钧陆鑫
李纯佳,覃伟,徐超华,刘洪博,毛钧,陆鑫
国外引进甘蔗栽培品种的光合气体交换参数遗传差异与聚类分析
李纯佳,覃伟,徐超华,刘洪博,毛钧,陆鑫
(云南省农业科学院甘蔗研究所/云南省甘蔗遗传改良重点实验室,云南开远 661699)
【目的】探明国外引进甘蔗栽培品种光合气体交换参数遗传变异特征,筛选优良基因型,并为甘蔗高光效育种技术优化提供参考依据。【方法】利用LI-6400便携式光合仪,于大伸长期对50份国外引进甘蔗栽培品种顶端全展叶6项光合气体交换参数进行测定,包括净光速率()、气孔导度(gs)、胞间CO2浓度(i)、蒸腾速率()、固有水分利用效率(WUEintr)、瞬时水分利用效率(WUEinst)。通过方差分析、广义遗传力计算、相关分析以及主成分分析明确气体交换参数变异特征,并通过聚类和判别分析筛选优异基因型。【结果】所有光合气体交换参数在参试基因型间差异均达到极显著水平,变异程度依次为gs>>>i>WUEintr>WUEinst。各项参数广义遗传力较高,除WUEinst为58.8%外,其余参数均达70%以上。除WUEinst与间相关性不显著外,其他气体交换参数间相关性均达显著水平。气孔导度同其他参数具有较强的非线性相关,体现了其对气体交换的重要调控作用。主成分分析共提取两项公因子,可分别解释为“碳同化性能”和“水分利用效率”,二者在基因型间的变化彼此独立,表明兼具高碳同化性能和高水分利用效率材料的筛选是可能的。通过聚类分析最终筛选出B4362、B51-410、US67-22、BH10-12、C323-87、Co685等6个碳同化能力突出、且同时具有极佳水分利用效率的优异基因型。【结论】国外引进甘蔗栽培品种中蕴含丰富的气体交换参数变异,遗传差异是该变异产生的主要原因。鉴定筛选到6份兼具高碳同化性能和高水分利用效率的优异材料,为甘蔗高光效育种提供了可靠的种质资源和优化建议。
甘蔗;国外引进栽培品种;光合气体交换参数;遗传差异;聚类筛选
0 引言
【研究意义】甘蔗(spp.)是我国和世界首要糖料作物,并在生物能源领域发挥着愈发重要的作用。作为世界上最高产的作物之一,强大的光合能力是甘蔗的显著生理特征[1-2]。光合作用是植物生物量的来源,对作物产量和品质均有重要影响[3]。因此,高光效育种被视为作物遗传改良的重要内容和研究方向。我国甘蔗育种起步较晚,为拓宽自育品种遗传基础、提高育种效率,陆续引进了大批国外栽培品种。这些材料蕴含丰富的遗传变异,对其开展光合气体交换参数遗传变异特征研究,可为我国甘蔗高光效育种亲本选择和技术优化提供参考依据。【前人研究进展】光合效能是复杂的多因素综合性状,叶绿素荧光[4]、光合放氧[5]、气体交换[6]等生理参数均可反映叶片光合效率。其中,气体交换参数凭借实时无损、直观准确的测量技术优势,在高等植物光合生理研究中应用广泛[6]。甘蔗高光效育种实践中,气体交换参数具有重要作用。首先,气体交换参数可用于甘蔗优异种质资源筛选。罗俊等[7]发现甘蔗光合气体交换参数在不同品种间存在较大差异,其中的3个参数可用于品种光合性能的聚类筛选。徐超华等[8]分析了甘蔗近缘属野生种质资源蔗茅()的光合气体交换数据,发现材料间差异较大,并从中筛选出了具有较高光合效率和较高水分利用效率的群体。其次,气体交换参数还可用于分析甘蔗光合特性遗传规律。吕建林等[9]发现各项气体交换参数在甘蔗组合间和组合内均存在高度分离,从而证明在甘蔗F1代材料中进行高光效筛选是可行的,并认为净光速率、气孔导度和蒸腾效率等3个气体交换性状的遗传差异主要由非加性效应引起。罗俊等[10]在实生苗中发现甘蔗气体交换性状受组合特殊配合力影响大于亲本一般配合力。【本研究切入点】光合性能的遗传变异是高光效育种的基础。国外甘蔗优良品种的引进和利用,极大丰富了我国甘蔗杂交育种基因库,对我国自育甘蔗品种产量、糖分、抗性的提升均做出了重要贡献。但其在高光效甘蔗新品种培育方面的潜力尚未得到充分利用。要深入挖掘引进品种的高光效育种潜力准确高效的光合气体交换特性分析是十分必要的工作。目前我国对国外引进甘蔗品种的评价多集中在生产特性[11-12]、表型[13-14]和分子遗传多样性[15-16]等方面,研究已从不同角度和水平揭示了国外引进甘蔗栽培品种中蕴含的丰富遗传变异,但这些变异在光合生理、尤其是气体交换特性上的具体体现仍是有待研究的问题。【拟解决的关键问题】本研究随机选取了国家甘蔗种质资源圃内保育的50份国外引进甘蔗栽培品种,对其光合气体交换参数进行了测量和分析,旨在探明其气体交换特性差异,筛选可用于高光效育种的优异种质资源,并为更大规模的高光效资源鉴定和品种选育的技术优化提供依据。
1 材料与方法
1.1 供试材料
试验材料为50份国外引进甘蔗栽培品种无性系(表1),从国家甘蔗种质资源圃中随机选取,由国家农作物种质资源共享平台提供。
1.2 试验设计
试验于云南省农业科学院甘蔗研究所国家甘蔗种质资源圃内进行。试验地位于北纬23.7°,东经103.25°,海拔约1 052 m,属低纬度亚热带高原季风气候,全年以晴天为主,日温差较大,年温差较小。年平均气温20°C左右,光照充足,年日照时数2 385 h,无霜期341 d。试验地土壤为砂质红壤,有机质含量21.2 g·kg-1,pH 7.8,全氮、全磷、全钾含量分别为0.111、0.149、1.200 g·kg-1,其中有效氮、有效磷、有效钾含量分别为67、18.6、166 mg·kg-1。
试验整体采用单因素完全随机设计,种植行长3 m,行距1.2 m,株距约0.5 m。于2017年3月初,选取健康完整的中上部单芽茎段下种,种植密度90 000芽/hm2。田间管理措施参照常规甘蔗高产栽培技术实行。试验期间各材料均健康生长,未见病虫害发生。
1.3 测定项目及方法
2017年7月至9月,甘蔗进入大伸长期。每一无性系选取长势具有代表性、均匀一致的植株3株,挂牌标识其+1叶进行测量,共测量5次。采用LI-6400型便携式光合仪(Li-Cor,USA)测定光合气体交换参数。于天气晴朗或少云的上午,待叶片经自然光诱导充分后进行测量,时间为9:30—11:30。以光合仪叶室固定叶片,5—10 min数值稳定后进行记录。
光合仪设定参数:气体流量500 μmol·mol-1,CO2浓度400 μmol·mol-1(由小钢瓶供给),叶室温度30°C,光强1 400 μmol·m-2·s-1(使用叶室LED红蓝光源)。

表1 参试国外引进甘蔗栽培品种信息
测定光合气体交换参数:净光速率(photosynthesis,),μmol·m-2·s-1;气孔导度(stomatal conductance,gs),mol·m-2·s-1;胞间CO2浓度(intercellular CO2concentration,i),μmol mol-1;蒸腾速率(transpiration rate,),mmol·m-2·s-1;叶片固有水分利用效率(intrinsic water use efficiency,WUEintr),μmol·mol-1;瞬时水分利用效率(instantanous water use efficiency,WUEinst),μmol·mol-1。其中,WUEintr=/gs,WUEinst=/E。
1.4 数据分析
使用SPSS 19.0软件进行方差分析、相关分析、主成分分析、判别分析。广义遗传力计算参照杜荣骞[17]方法。使用Origin 2017 Pro软件绘制拟合曲线。使用HemI软件[18]绘制热图,并进行聚类分析。原始数据标准化为Z分数进行主成分分析、判别分析、聚类热图绘制。
2 结果
2.1 光合气体交换参数遗传差异分析
50份国外引进甘蔗栽培品种光合气体交换参数基因型间差异如表2所示。各材料平均为27.3 μmol·m-2·s-1,最高的无性系VMC87-95(36.2 μmol·m-2·s-1)与最低的CAC113(15.9 μmol·m-2·s-1)间存在2.27倍差异;gs均值为0.229 mol·m-2·s-1,最高为MY5465(0.367 mol·m-2·s-1),最低为CAC113(0.117 mol·m-2·s-1),二者间差异达3.14倍;i均值为144 μmol·mol-1,最高的MY5465(190 μmol·mol-1)与最低的FR93- 264(103 μmol·mol-1)间差异为1.85倍;均值为3.98 mmol·m-2·s-1,最高为Co330(5.42 mmol·m-2·s-1),最低为CAC113(2.63 mmol·m-2·s-1),差异达2.06倍;WUEintr均值为126 μmol·mol-1,最高的FR93-264(157 μmol·mol-1)与最低的MY5465(95 μmol·mol-1)间存在1.65倍差异;WUEinst均值为6.86 μmol·mol-1,最高为C2858-74(7.66 μmol·mol-1),与最低的CAC113(5.65 μmol·mol-1)间差异为1.36倍。
倍数差异和变异系数均显示gs为变异最剧烈的参数,其次为,再次为,其后为i和WUEintr,WUEinst变异程度最小。方差分析结果显示,所有光合气体交换参数在参试基因型间差异均达到极显著水平(<0.001)。总体而言,各参数广义遗传力较高,其中最高为,达74.8%;其后依次为gs、i、、WUEintr,均达70%以上;WUEinst最低,亦达58.8%。
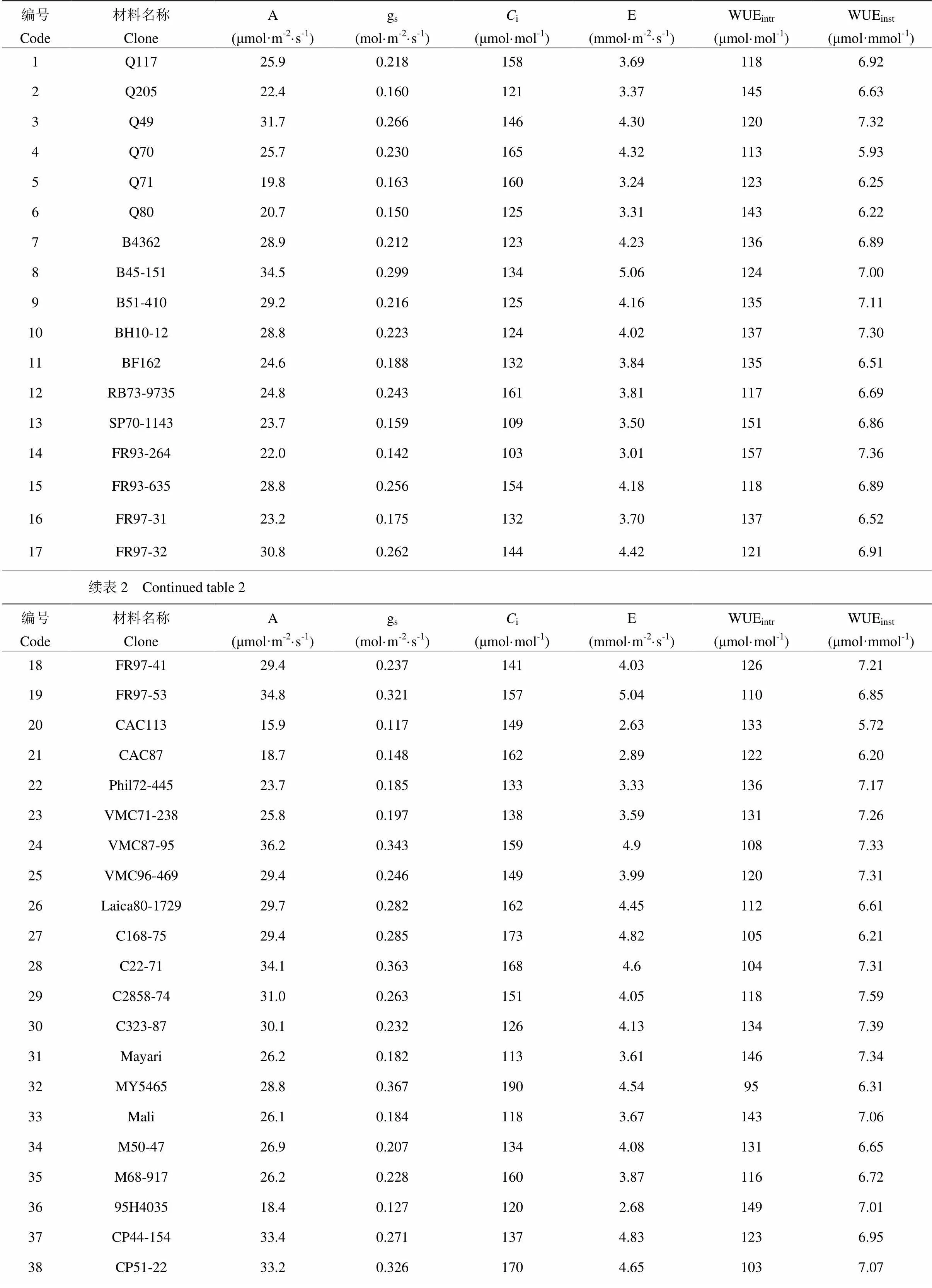
表2 国外引进甘蔗栽培品种间气体交换参数遗传差异
***,在<0.001水平上达到极显著差异 ***, statistically significant at<0.001
2.2 光合气体交换参数间相关性
虽然WUEinst为与的比值,但其与的相关系数仅为0.008,相关性不显著。除此以外,其他光合气体交换参数间相关性均达到极显著水平(表3)。
除WUEinst外,gs与其他气体交换参数间的相关性均较强。其中,与、i、呈正相关,与WUEintr呈负相关。这些相关关系均为非线性,使用指数函数和幂函数衍生的超越方程可较好描述其函数关系(图1-a—图1-d)。与间同样存在较强的非线性正相关关系,可以幂函数方程= 167.360.12- 170.16(2=0.85)拟合(图1-e)。WUEintr与、gs、i、均呈负相关,其中,与i的相关性极强,使用线性方程= -0.68+ 222.92进行拟合,2可达0.90(图1-f)。虽然同为水分利用效率,WUEinst与WUEintr相关系数较低,相关性较弱。
2.3 光合气体交换参数主成分分析
表4表明,以特征根≥1且积累贡献率≥85%为判别条件,共可提取两项主成分因子。PC1特征根达3.459,对总体方差的贡献率高达57.651%;PC2特征根为1.907,所含方差占总方差的31.777%。两项主成分因子共解释了89.427%的总体变异,表明主成分因子提取是有效的,包含了本试验国外引进甘蔗栽培品种光合气体交换数据的主要信息。

图1 气体交换参数间相关关系

表3 光合气体交换参数间相关系数
表中数字为皮尔森相关系数,相关显著性双侧检验。**,在<0.01水平达到极显著相关;ns,相关性不显著
Pearson’s correlation coefficients were shown with numbers in above table. Significance was two-tailed tested. **, statistically significant at<0.01; ns, not significant

表4 光合气体交换参数主成分因子得分及载荷
主成分因子PC1的载荷主要来自、gs、,且均为正向(表4)。值为叶片CO2同化速率的直接体现,gs、为叶片固碳性能的重要组成部分和表征因子,与具有极显著的正相关关系(表3,图1-a—图1-e)。可见,PC1因子可称为“碳同化性能”。主成分因子PC2主要由WUEinst和WUEintr提供正向载荷,其负向载荷主要由i承担(表4)。i与WUEintr间存在极显著的线性负相关关系(表2,图1-f),为WUEintr的负向指征。因此,PC2因子可称为“水分利用效率”。
根据各基因型的主成分因子得分分析因子间相关性,发现PC1与PC2间无相关关系,彼此独立(2= 0.02,= 0.85)(图2)。这为同时具有优异碳同化性能和水分利用效率的优异材料筛选提供了可能。

图2 主成分因子间关系
2.4 聚类与判别分析
R型聚类结果显示,6项气体交换参数中,、gs、i、等固碳相关参数聚成一组,WUEintr、WUEinst等两项水分利用效率参数聚成一组(图3)。Q型聚类中,50个基因型总体划分为两个群。其中一个群包含I、II 2个组,另一个群由III、IV、V 3个组组成。热图中,两个群特征差异明显,其中一个群固碳相关指标水平较弱,水分利用效率较高,另一个群则相反(图3)。总体而言,聚类结果与主成分因子分析结果较为一致。
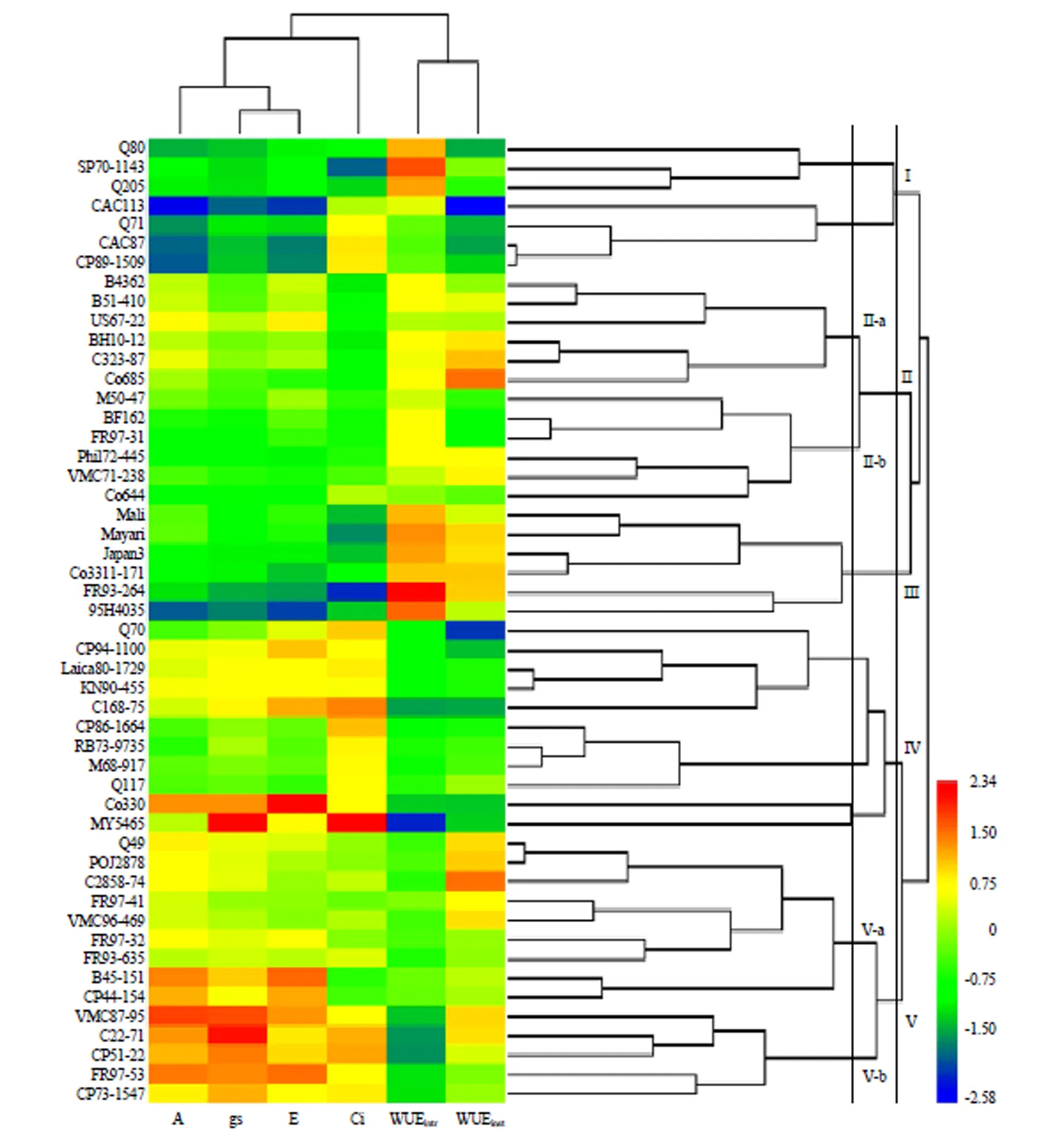
图3 国外引进甘蔗栽培品种气体交换参数聚类热图
以上述聚类结果作为先验概率,使用步进法对Q型聚类结果进行贝叶斯判别分析。经Wilk’s Lambda检验,gs、、WUEintr、WUEinst等4项参数作为引入变量进入判别模型,其在各组分类判别函数中的系数如表5所示。上述模型判别结果显示分组正判率为100%,说明图3所示聚类结果是可靠的。
各基因型的聚类分组的气体交换特征如表6所示。I组包含7个基因型,、gs、、WUEinst均为5组中最低,i和WUEintr为中等水平,表明该组材料固碳性能和水分利用高效率均较差,属气体交换性能最差的类型;II组包含11个基因型,其中6项气体交换性状均属中等水平,表明该组材料为固碳性能和水分利用效率较为均衡的类型;III组包含6个基因型,和均较低,WUEintr和WUEinst为各组最高,可视为水分利用效率的极端材料;IV组包含11个基因型,具有中等水平的和较高的gs、,但其i过高,WUEintr和WUEinst均较属最低水平,为气体交换性能较差的类型;V组包含16个基因型,其、gs和均为最高水平,i和WUEinst较高,WUEintr较低,综合认为该组材料为固碳性能的极端类型,同时具一定水平的水分利用效率。

表5 各组判别函数系数
II组和V组具有进一步筛选的价值,且所含材料较多,可各自划分出II-a、II-b、V-a、V-b等4个亚组(图3,表6)。其中,II-a亚组包含B4362、B51-410、US67-22、BH10-12、C323-87、Co685等6个基因型,总体表现为高,i低,gs和适中,WUEintr和WUEinst为4个亚组中最高,同时具有优异的固碳性能和水分利用效率,为本研究中气体交换特征较为理想的一组材料。

表6 聚类分组气体交换特征
表中数字为各组均值,使用Duncan法进行多重比较,不同字母表示组间差异在<0.05上达到显著水平
Mean values of each group were shown in the table. Multiple comparison was conducted using Duncan’s method. Different letters mean statistical significance between groups at0.05
3 讨论
3.1 国外引进甘蔗栽培品种光合气体交换参数遗传变异
光合作用为作物生长和产量形成提供必要的物质基础。作为主要光合器官,叶片的固碳作用为高等植物积累了近90%的生物量[2-3]。叶片光合能力的遗传改良,已成为作物高光效育种的热点方向之一[3]。本文对50份国外引进甘蔗栽培品种的叶片气体交换参数进行了分析,发现各项参数中存在巨大变异,且广义遗传力较高(表2)。这表明遗传因素,而非环境饰变,是国外引进甘蔗品种叶片气体交换参数变异产生的主要原因。
前人利用国外引进栽培品种开展了甘蔗叶片光合效率研究[7,10],也显示了气体交换参数在不同材料间存在显著差异,但未解释其产生的原因。Jackson[19]和LI等[20]此前的工作表明遗传差异对甘蔗及近缘属植物叶片光合水平差异的形成具有决定性作用,但所用无性系群体由多个种属材料混合而成,遗传背景复杂,天然具有较高的异质性。本研究在遗传背景相对狭窄的国外引进栽培品种无性系群体中,揭示了由其内在遗传差异导致的气体交换参数的巨大变异。原种和近缘属资源的杂交利用可拓宽甘蔗种质基础,但作为高代材料,现有国外引进甘蔗栽培品种亦可为我国甘蔗高光效育种提供较为丰富的遗传资源,种质创新与品种选育实践中应予关注。
3.2 优异基因型筛选及其气体交换特征
优异资源的鉴定与筛选是开展甘蔗高光效品种选育的基础与前提。从气体交换角度来看,光合作用即为一个“以散失自身水分为代价吸收外界CO2”的过程[21]。前人的报道[7-8,22]和本文主成分分析结果(表4)也显示,CO2固定和水分散失是叶片光合气体交换的两大主题。因此,高光效优异材料的筛选,不应一味追求光合速率,而应注重碳同化性能与水分利用效率的平衡。借助聚类分析和判别分析,笔者筛选和鉴定出了可供甘蔗高光效育种优先选择的6份材料,分别为B4362、B51-410、US67-22、BH10-12、C323-87,以及Co685。这一组材料的气体交换特征在于固碳性能优异,同时水分利用效率极佳(表6),在较高水平上取得了二者间平衡。
由于碳同化性能与水分利用效率可各自独立在基因型间变化(图2),二者间的平衡是可以达成的。但更多情况下,植物固碳能力和水分利用效率间的矛盾难以调和[23]。本文和其他大量研究[8,19-20]结果表明,光合速率与固有水分利用效率间存在中等以上强度(>0.5)的显著负相关关系(表2),即为该矛盾的具体体现。本研究中部分材料取得了二者的平衡,应为人工选择压力间接作用的结果。参试材料虽为不同国家的不同育种机构育成,但均由常规选择方法选育,无一为专门的高光效育种计划的产物。甘蔗的常规选育是基于表型的,而部分表型性状,尤其是叶部表型,与其光合能力紧密相关。如叶片厚度可决定单位面积叶片的Rubisco酶含量,从而影响光合速率[1-2];而叶型则与耐旱性有关,叶片长宽比与水分利用效率具有一定正相关关系[2]。相关表型性状的综合考量,可间接完成作物叶片固碳能力和水分利用效率的微妙调和与平衡[24-25]。以上6个优异基因型固碳能力与水分利用效率在较高水平达成平衡,即为表型选择压力间接塑造甘蔗叶片气体交换生理特征的典型体现。
3.3 甘蔗大规模高光效种质资源筛选方案优化
气体交换数据是目前评价叶片光合能力的主流标准[6],但其测量结果易受环境影响。云层遮盖、日光角度、植株自身和邻株的遮光、空气流动等难以避免的扰动,都将通过气孔行为对各气体交换参数数值产生影响[3,22]。广义遗传力可反映环境方差在总体方差中的比例[17],因此可作为气体交换数据质量的衡量指标[20]。为取得更高的广义遗传力,往往需要提高测量强度[17]。这使得气体交换测量成为一项劳动密集工作,对大规模的高光效材料筛选犹是如此。
根据全生长期测量结果,LI等[20]预计甘蔗每个基因型约需至少20次测量才能令Hb达到70%。但本研究仅以每个基因型15次(3棵植株×5次测量/株)的测量强度便获得了这一水平的广义遗传力(表2),这可能与选择在大伸长期进行集中测量有关。这一时期,同一品种的各植株长势趋于一致,而不同品种的生长状态差异愈发凸显[1-2],测量结果能够充分体现遗传效应带来的差异,压缩环境方差所占比例。我们结合严格控制仪器测量参数以避免误差项过大,最终以较低的测量强度取得了较好的测量结果。
仅针对气孔导度开展筛选,亦被证明是有效的替代方案。这一方案的优势在于气孔导度直接或间接测量设备价格均远低于光合仪,且测量更为快捷便利。研究机构可负担多台,并以更快的单叶测量速度提高测量通量,从而满足大规模筛选的需求。气孔行为对叶片光合水平的关键影响,是该方案合理性的基础。以本文为例,气孔导度在所有气体交换参数中遗传变异最为强烈(表2),对其他参数具有显著调节作用(图1),同时还是“碳同化性能”主成分因子的主要载荷性状(表4)和聚类结果的判别性状(表5)。在水稻()[26]、小麦()[27]、马铃薯()[28]等作物的生理选育中,针对气孔导度的筛选已得到实际运用。甘蔗中,Basnayake[29]和Luo[30]已明确提出气孔导度可作为耐旱品种选育指标。鉴于适宜的气孔导度亦为本文6份优异材料气体交换的显著特征,笔者认为这一替代方案对甘蔗高光效种质筛选同样具有应用潜力。
4 结论
通过50份国外引进甘蔗栽培品种的叶片气体交换测量和分析发现,净光速率、气孔导度、胞间CO2浓度、蒸腾速率、固有水分利用效率和瞬时水分利用效率等6项参数存在极显著差异,材料间遗传差异是其产生的主要原因。各基因型间,碳同化性能与水分利用效率可以兼顾,B4362、B51-410、US67-22、BH10-12、C323-87、Co685等6份材料碳同化性能和水分利用效率俱佳,可供高光效育种利用。本文研究结果还为甘蔗高光效育种大规模气体交换测量提出了优化方案。
致谢:感谢澳大利亚联邦科学与工业研究组织(Commonwealth Scientific and Industrial Research Organization,CSIRO)高级研究员Phillip Jackson博士对本文的帮助。
[1] 李杨瑞. 现代甘蔗学. 北京: 中国农业出版社, 2010.
LI Y R.Beijing: China Agriculture Press, 2010. (in Chinese)
[2] MOORE P H, BOTHA F C.. John Wiley & Sons, 2014.
[3] EVANS J R. Improving photosynthesis., 2013, 162(4): 1780-1793.
[4] BAKER N R. Chlorophyll fluorescence: A probe of photosynthesis in vivo., 2008, 59(1): 89-113.
[5] WALKER D A, HILL R. The relation of oxygen evolution to carbon assimilation with isolated chloroplasts., 1967, 131(2): 330-338.
[6] STINZIANO J R, MORGAN P B, LYNCH D J, SAATHOFF A J, MCDERMITT D K, HANSON D T. The rapid A-Ci response: Photosynthesis in the phenomic era., 2017, 40(8): 1256-1262.
[7] 罗俊, 张华, 徐良年, 吕建林, 陈如凯. 甘蔗不同品种光合特性比较及其聚类分析. 中国农业科学, 2005, 38(8): 1562-1569.
LUO J, ZHANG H, XU L N, LÜ J L, CHEN R K. Comparison and cluster analysis of photosynthetic character of different sugarcane varieties., 2005, 38(8): 1562-1569. (in Chinese)
[8] 徐超华, 李纯佳, 陆鑫, 刘新龙, 李旭娟, 毛钧, 林秀琴, 刘洪博, 字秋燕, 蔡青. 甘蔗近缘种蔗茅()光合气体交换特性的差异分析. 中国农业科学, 2016, 49(15): 2909-2920.
XU C H, LI C J, LU X, LIU X L, LI X J, MAO J, LIN X Q, LIU H B, ZI Q Y, CAI Q. Comparison of photosynthetic gas exchange parameters for sugarcane close relative species., 2016, 49(15): 2909-2920. (in Chinese)
[9] 吕建林, 陈如凯, 张木清, 廖建峰, 李才明. 甘蔗F1 代光合特性.甘蔗, 1998, 5(2): 1-6.
LÜ J L, CHEN R K, ZHANG M Q, LIAO J F, LI C M. Thecharacteristic of photosynthesis in sugarcane F1., 1998,5(2): 1-6. (in Chinese)
[10] 罗俊, 周会, 张木清, 陈如凯, 张华. 能源甘蔗主要经济和光合性状的遗传分析. 应用与环境生物学报, 2004, 10(3): 268-273.
LUO J, ZHOU H, ZHANG M Q, CHEN R K, ZHANG H. Genetic analysis of main economic and photosynthetic traits in energy sugarcane., 2004, 10(3): 268-273. (in Chinese)
[11] 马丽, 蔡青, 应雄美, 刘新龙, 陆鑫, 毛钧. 29个国外甘蔗种质3年宿根性的产质量评价. 中国糖料, 2010(1): 42-43.
MA L, CAI Q, YING X M, LIU X L, LU X, MAO J. Evaluation on yield and sucrose content to foreign ratoon sugarcanes., 2010(1): 42-43. (in Chinese)
[12] 刘家勇, 范源洪, 杨洪昌, 赵俊, 赵培方, 吴才文, 陈学宽. 31个国外甘蔗引进种灰色多维综合评估. 西南农业学报, 2006, 19(4): 683-687.
LIU J Y, FAN Y H, YANG H C, ZHAO J, ZHAO P F, WU C W, CHEN X K. Fuzzy multidimensional comprehensive evaluation method for 31 overseas varieties., 2006, 19(4): 683-687. (in Chinese)
[13] 昝逢刚, 吴才文, 赵培方, 赵俊, 夏红明, 杨昆, 姚丽, 覃伟, 刘家勇. 国外引进甘蔗种质表型性状的遗传多样性分析. 西南农业学报, 2014, 27(4): 1369-1373.
ZAN F G, WU C W, ZHAO P F, ZHAO J, XIA H M, YANG K, YAO L, QIN W, LIU J Y. Study on phenotypic diversity in exotic sugarcane germplasm., 2014, 27(4): 1369-1373. (in Chinese)
[14] 赵俊, 吴才文, 赵培方, 夏红明, 昝逢刚, 杨昆, 姚丽, 刘家勇. 引进甘蔗种质工艺与农艺性状的相关性及聚类分析. 湖南农业大学学报(自然科学版), 2012, 38(5): 476-481.
ZHAO J, WU C W, ZHAO P F, XIA H M, ZAN F G, YANG K, YAO L, LIU J Y. Principal component analysis and cluster analysis of agronomic and quality traits of exotic sugarcane germplasm.(), 2012, 38(5): 476-481. (in Chinese)
[15] 刘新龙, 李旭娟, 刘洪博, 马丽, 徐超华, 范源洪. 云南甘蔗常用亲本资源遗传多样性的 SSR 分析. 植物遗传资源学报, 2015, 16(6): 1214-1222.
LIU X L, LI X J, LIU H B, MA L, XU C H, FAN Y H. Genetic diversity analysis of Yunnan commonly-used parents by using SSR marker., 2015, 16(6): 1214-1222. (in Chinese)
[16] 昝逢刚, 应雄美, 吴才文, 赵培方, 陈学宽, 马丽, 苏火生, 刘家勇. 98份甘蔗种质资源遗传多样性的AFLP 分析. 中国农业科学, 2015, 48(5): 1002-1010.
Zan F G, YINIG X M, WU C W, ZHAO P F, CHEN X K, MA L, SU H S, LIU J Y. Genetic diversity analysis of 98 collections of sugarcane germplasm with AFLP markers., 2015, 48(5): 1002-1010. (in Chinese)
[17] 杜荣骞. 生物统计学(第四版). 北京: 高等教育出版社, 2014.
DU R J.. Beijing: Higher Education Press, 2014. (in Chinese)
[18] DENG W, WANG Y, LIU Z, CHENG H, XUE Y. HemI: A toolkit for illustrating heatmaps., 2014, 9(11): e111988.
[19] JACKSON P, BASNAYAKE J, INMAN-BAMBER G, LAKSHMANAN P, NATARAJAN S, STOKES C. Genetic variation in transpiration efficiency and relationships between whole plant and leaf gas exchange measurements inspp. and related germplasm., 2015, 67(3): 861-871.
[20] LI C, JACKSON P, LU X, XU C, CAI Q, BASNAYAKE J, LAKSHMANAN P, GHANNOUM O, FAN Y. Genotypic variation in transpiration efficiency due to differences in photosynthetic capacity among sugarcane-related clones., 2017, 68(9): 2377-2385.
[21] FARQUHAR G D, EHLERINGER J R, HUBICK K T. Carbon isotope discrimination and photosynthesis., 1989, 40: 503-537.
[22] DRIEVER S M, LAWSON T, ANDRALOJC P J, RAINES C A, PARRY M A J. Natural variation in photosynthetic capacity, growth, and yield in 64 field-grown wheat genotypes., 2014, 65(17): 4959-4973.
[23] SINCLAIR T R. Is transpiration efficiency a viable plant trait in breeding for crop improvement?, 2012, 39(5): 359-365.
[24] FLOOD P J, HARBINSON J, AARTS M G. Natural genetic variation in plant photosynthesis., 2011, 16(6): 327-335.
[25] DEVI M J, SINCLAIR T R, VADEZ V. Genotypic variation in peanut for transpiration response to vapor pressure deficit., 2010, 50(1): 191-196.
[26] HORIE T, MATSUURA S, TAKAI T, KUWASAKI K, OHSUMI A, SHIRAIWA T. Genotypic difference in canopy diffusive conductance measured by a new remote-sensing method and its association with the difference in rice yield potential., 2006, 29(4): 653-660.
[27] ZHENG T C, ZHANG X K, YIN G H, WANG L N, HAN Y L, CHEN L, HUANG F, TANG J W, XIA X C, HE Z H. Genetic gains in grain yield, net photosynthesis and stomatal conductance achieved in Henan Province of China between 1981 and 2008., 2011, 122(3): 225-233.
[28] PRASHAR A, YILDIZ J, MCNICOL J W, BRYAN G J, JONES H G. Infra-red thermography for high throughput field phenotyping in., 2013, 8: e658166.
[29] BASNAYAKE J, JACKSON P A, INMAN-BAMBER N G, LAKSHMANAN P. Sugarcane for water-limited environments. Variation in stomatal conductance and its genetic correlation with crop productivity., 2015, 66(13): 3945-3958.
[30] LUO J, PAN Y B, XU L P, ZHANG Y Y, ZHANG H, CHEN R K, QUE Y X. Photosynthetic and canopy characteristics of different varieties at the early elongation stage and their relationships with the cane yield in sugarcane., 2014: 707095.
(责任编辑 杨鑫浩)
Genetic Variations and Cluster Analysis of Photosynthetic Gas Exchange Parameters in Exotic Sugarcane Cultivars
LI ChunJia, QIN Wei, XU ChaoHua, LIU HongBo, MAO Jun, LU Xin
(Sugarcane Research Institute, Yunnan Academy of Agricultural Sciences/Yunnan Key Laboratory of Sugarcane Genetic Improvement, Kaiyuan 661699, Yunnan)
【Objective】To identify genotypes with high photosynthesis capacity and to optimize methods for further screening, genetic variations of photosynthetic gas exchange parameters among exotic sugarcane cultivars was investigated. 【Method】Using a LI-6400 portable photosynthesis system, gas exchange measurements were conducted on the youngest fully-expanded leaves of 50 cultivars introduced from abroad at their grand-growth stage. Six parameters were measured, namely photosynthesis (), stomatal conductance (gs), intercellular CO2concentration (i), transpiration rate (), intrinsic water use efficiency (WUEintr), and instantaneous water use efficiency (WUEinst). Key statistics were determined including broad sense heritability, correlations among parameters. Besides, variation in responses among cultivars was characterized using principal component analysis, while cluster and discriminant analysis were conducted for elite screening. 【Result】 Significant genetic variation was found for every parameter, with the order (highest to lowest variation) being gs,,,i, WUEintr, to WUEinst. Broad heritability was high (>70%) for all parameters except WUEinst(which was 58.8%). There were significant correlations between all the parameters except that between WUEinstand. Stomatal conductance had strong nonlinear relationships with other parameters, consistent with its pivotal role in regulating leaf gas exchange. Two principal components were extracted through principal component analysis, which could be interpreted as being predominately related to "carbon assimilation" and "water use efficiency" respectively. These two components varied independently among the clones, indicating the possibility of screening for elites harboring both high carbon assimilation and high water use efficiency. Cluster analysis identified a group of six genotypes which had both high carbon assimilation and water use efficiency. 【Conclusion】 Significant variation due to genetic differences in gas exchange parameters exists among exotic sugarcane cultivars. An elite group of cultivars with both high carbon assimilation and water use efficiency were identified, including B4362, B51-410, US67-22, BH10-12, C323-87, and Co685. Methods to optimize large-scale screening in sugarcane breeding programs for favorable photosynthetic capacity were also determined and discussed.
spp.; exotic cultivars; photosynthetic gas exchange parameters; genetic variation; clustering-based screening
2017-11-29;
2018-03-06
云南省应用基础研究计划面上项目(2016FB067,2017FB054)、福建农林大学国家甘蔗工程技术研究中心开放课题基金(2016.2.1)、农作物种质资源保护项目(2017NWB017)、云南省农业科学院青年科技人才培养专项
李纯佳,E-mail:lcjzt@163.com。
陆鑫,E-mail:xinlu_ky@126.com
10.3864/j.issn.0578-1752.2018.12.006