转AtTGA4小麦田间耐低磷胁迫的功能分析
2018-06-29凌炳琦柏星轩周永斌王春霄徐兆师马有志陈明张小红
凌炳琦,柏星轩,周永斌,王春霄,徐兆师,马有志,陈明,张小红
转小麦田间耐低磷胁迫的功能分析
凌炳琦1,柏星轩2,周永斌2,王春霄2,徐兆师2,马有志2,陈明2,张小红1
(1西北农林科技大学生命科学学院,陕西杨凌 712100;2中国农业科学院作物科学研究所,北京 100081)
【目的】将bZIP类转录因子基因转化小麦创制耐低磷转基因小麦新材料,同时分析提高小麦抗逆性的生理机制,为小麦耐低磷胁迫分子育种奠定基础。【方法】采用最小表达框基因枪转化法将和筛选标记基因共转化受体小麦石4056,通过PCR检测筛选出无并能稳定遗传的转基因小麦新株系。基于试验地土壤养分含量状况施用不同水平的磷肥,形成一定程度的正常和低磷营养胁迫,对转基因小麦新株系进行低磷胁迫耐受性试验。在开花期进行了光系统Ⅱ原初光能转化效率(light efficiency of the light system Ⅱ,Fv/Fm),叶绿素相对含量(soil and plant analyzer development readings,SPAD值)和气冠温差(canopy temperature depression,CTD)等生理指标的测定,在成熟期进行了株高、分蘖数、穗粒数等农艺性状的调查,并在小麦收获后进行了产量及不同组分(根、茎、叶、籽粒)磷浓度和磷吸收、残留总量的测定和统计。【结果】PCR分析结果证明,已在石4056小麦中稳定遗传至T4代,共获得4个稳定转基因株系。根据土壤养分含量测定结果,在正常条件地块施加812.39 kg·hm-2的过磷酸钙,低磷处理地块不施磷肥。产量及农艺性状统计结果显示,转基因株系L1和L2在正常和低磷胁迫条件下的产量相对于受体对照小麦显著增加,正常条件下产量增幅为5.3%—8.6%,低磷胁迫下产量增幅为4.4%—7.7%。在低磷胁迫条件下,过表达的转基因小麦种子千粒重显著比受体显著增加。开花期田间生理指标测定结果显示,转基因株系L1和L2在低磷条件下的Fv/Fm和CTD明显优于受体,而SPAD值没有明显差异。田间调查时发现,低磷条件下受体比转基因材料提早结束灌浆,表现在穗子提早变黄。成熟末期磷含量测定结果显示,转基因株系L1和L2在低磷条件下茎杆磷浓度比受体显著提高,在其他组织中则无显著差异。2个转基因株系在低磷条件下茎、叶和籽粒吸收、残留的总磷含量都要高于受体,地上部总磷含量增幅达6.38%—17.47%。转基因材料表达量分析结果显示,目标基因在株系L2中的表达量较株系L1中的低,是株系L1的0.69倍。【结论】在低磷胁迫条件下可以显著提高转基因小麦对磷元素的吸收及运输,提高转基因小麦的产量,进而提高转基因小麦对低磷胁迫的耐性。
转基因小麦;;低磷营养胁迫;产量;生理指标
0 引言
【研究意义】磷是植物体生长发育所需的大量元素之一,它在膜系统的构成、物质和能量代谢、遗传等生理生化过程中扮演着极其重要的角色[1]。研究表明,耕地土壤中的磷主要以无机磷、有机磷及其他金属螯合磷的形式存在;一般情况下,植物以吸收无机磷为主,而无机磷当中可被直接吸收利用的有效磷却占总磷含量的不足0.5%[2-3]。如今植物当季从土壤中吸收的磷大多来源于人为施加的磷肥,而磷肥在土壤中很容易被吸附和固定,并且耕地中的可溶性磷易随着灌溉和降雨流失,这不仅导致了磷肥的当季利用率低下,也造成耕地土壤无效磷库的逐年增长,更引发了许多环境问题[4-5]。小麦是中国第二大粮食粮作物,在种植过程中普遍面临着土壤有效磷不足的问题;随着人口的不断增长、生活质量需求的不断提高和不断缩小的耕地面积,培育稳产、高产的耐低磷胁迫小麦迫在眉睫[6-7]。因而发掘新的耐低磷胁迫基因,通过转基因技术创制耐低磷胁迫小麦具有重要的实际意义。【前人研究进展】目前,在拟南芥、烟草、水稻等植物低磷逆境胁迫研究中发现了许多起重要作用的转录因子。其中WANG等[8]和Rubio等[9]在拟南芥中发现了一个以AtPHR1蛋白为中心的磷信号调控网络,AtPHR1蛋白属于MYB-CC家族转录因子,能在低磷条件下调控其下游基因的表达(如磷转运蛋白基因等),增强拟南芥对磷及其他元素的吸收,提高其对低磷的耐受性,该基因还能通过增加单穗的结实率从而促进转基因小麦的增产。Devaiah等[10]在拟南芥中发现了WRKY型转录因子AtWRKY75,它的表达受低磷胁迫诱导,并且其RNAi突变体对低磷的敏感度更高,表现出提早的花青素积累。Sano等[11]发现,给受到低磷胁迫的烟草BY2细胞补充磷元素能够诱导(bZIP类)的表达。Yi等[12]在水稻中发现了(表达bHLH型转录因子),在水稻低磷处理条件下,过表达能提高水稻对磷的吸收,表现在转基因植株根系和茎生物量的增加,进而提升转基因水稻的低磷耐受性。Zhang等[13]发现大豆在低磷胁迫条件下能诱导的表达,其表达产物通过与GmSPX1互作,对大豆应答低磷胁迫中起负调控作用。基于前人的研究,表明不同类型的转录因子在植物对抗低磷非生物胁迫中发挥着重要的作用。【本研究切入点】拟南芥AtTGA4属于bZIP转录因子家族成员,Zhong等[14]研究发现,在拟南芥中能被干旱和低氮胁迫诱导表达,该基因的过表达可以显著提高转基因拟南芥的抗旱性,但是还未有任何与植物低磷胁迫响应相关的研究报导。而本研究将通过基因枪介导法转化至常规小麦品种石4056中,通过PCR检测获得4个稳定转基因小麦株系,用以进行田间低磷胁迫试验。【拟解决的关键问题】本研究通过试验鉴定转小麦的低磷耐受性,创制耐低磷胁迫转基因小麦新材料,分析提高石4056小麦低磷耐受性的生理机制,为小麦耐低磷胁迫分子育种奠定基础。
1 材料与方法
1.1 材料及种植条件
采用最小表达框的基因枪转化法,将和分别与只含有启动子(Ubiquitin)、目的基因、终止子(Nos)的DNA片段连接,通过轰击石4056小麦的愈伤组织,共获得7株T0代阳性植株。后经逐年的筛选、鉴定和繁育,共获得4个稳定遗传的T3代转基因株系供试,依次命名为AtTGA4-L1、AtTGA4-L2、AtTGA4-L3和AtTGA4-L4。
试验地位于北京市顺义区后桑园村中国农业科学院顺义试验基地(40º18′N,116º28′E)。2015年10月秋播前,以S型土样采集办法在不同营养地块均匀的采集6份土样,用以测定土壤中碱解氮、有效磷和速效钾含量,后根据《北京市土壤养分指标评定规则》和测定结果进行施肥[15]。供试材料采用完全随机区组的种植方式,通过自动播种机按照3.14×106·hm-2基本苗进行播种,每个种植小区6 m2(1.5 m×4 m),每个营养地块3个生物学重复。播种后的日常田间管理由试验基地相关责任人负责。
1.2 转基因小麦的常规PCR检测
采用GMO作物提取检测试剂盒(TIANGEN)提取转基因小麦全基因组DNA(于拔节期从每个转基因株系中随机选取10株对其倒二叶进行全基因组DNA的提取),并根据和Nos终止子序列、和Nos终止子序列通过DNAman软件设计引物对其进行常规PCR检测。
的检测引物:正向引物为5′-TCAAGCT GATCATCTGAGACAT-3′;反向引物为5′-ATAAAAA CCCATCTCATAAATAACG-3′,扩增片段为300 bp。PCR反应体系为10 μL 2×Taq PCR StarMix with Loading Dye(GenStar)、正向引物和反向引物各0.8 μL(10 μmol·L-1)、200—600 ng的DNA模板,补加蒸馏水至20 μL。PCR反应程序采用Touchdown的方法:95℃10 min;95℃30 s,58℃30 s(从第二个循环开始,每一轮循环降低0.5℃),72℃30 s,20个循环;95℃30 s,52℃30 s,72℃30 s,20个循环;72℃10 min,4℃保存。
的检测引物:正向引物为5′-CGATTCCCAGT CATCAAGTG-3′;反向引物为5′-ATTCAGCGACTT CTGGCA-3′,扩增片段大小为380 bp。PCR反应体系为10 μL 2×Taq PCR StarMix with Loading Dye(GenStar)、正向引物和反向引物各0.5 μL(10 μmol·L-1)、200—600 ng的DNA模板,补加蒸馏水至20 μL。PCR反应程序为95℃10 min;95℃30 s,56℃30 s,72℃30 s,25个循环;72℃10 min,4℃保存。
扩增产物经1%浓度的琼脂糖凝胶检测并拍照。
1.3 转基因小麦产量及部分农艺性状分析
虽然产量及其农艺性状受田间多种因素的影响,但其能直观的反应出供试材料在实际农业生产环境下的产出状况[16-19]。因此,在收获期进行产量及以下农艺性状的统计和分析:产量(kg·hm-2)由小区产量折算;分穗数(×104/hm2)则通过在小区长势均匀的样段中随机选取2段1 m行长的样段进行全分蘖数的统计,折算成小区实际分蘖数,进而换算;穗粒数则是在小区长势均匀的样段中随机取完整的20个穗子进行穗粒数清点而得出。
1.4 转基因小麦开花期部分生理指标的测定
1.4.1 PSⅡ原初光能转化效率(Fv/Fm) 于晴天的14:00时在小区中随机选取3棵单株,用捷克产的FluorPen手持叶绿素荧光仪进行原初光能转化效率的测定。
1.4.2 叶绿素相对含量(SPAD) 于晴天在小区中随机选取5棵单株,用产自日本的SPAD-502型叶绿素仪对其旗叶的叶基、叶片中部和叶尖进行叶绿素相对含量的测定,取平均值。
1.4.3 气冠温差(CTD) 于晴天的13:00至14:00时段,用德国产的Optris红外测温仪对供试材料小区的两端和中部进行冠层温度和大气温度的测量,通过计算得出气冠温差。
1.5 转基因小麦不同组分磷含量的测定
为了比较转基因材料与受体的磷吸收、利用能力,在小麦成熟末期对供试材料根、茎、叶和籽粒的磷浓度和吸收、残留总量进行了测定和统计分析。
采用浓H2SO4-H2O2消煮法结合全自动流动分析仪测定供试材料不同组分磷浓度:小麦成熟后,从小区中选取10株长势一致的小麦进行根、茎、叶、籽粒的分离,用蒸馏水洗净后,烘干、称重,并用FW100型高速万能粉碎机(天津)进行充分粉碎;每份粉碎样品称取3份0.5000 g进行H2SO4-H2O2消煮,最后稀释到合适的浓度并通过全自动流动分析仪进行全磷含量的测定[20]。由于根系无法完整的取样,只能对成熟末期供试材料地上部各组分吸收、残留总磷含量进行统计和分析。
1.6 转基因小麦AtTGA4的定量表达分析
取正常营养条件下的转基因株系苗期叶片,采用DP432植物总RNA提取试剂盒(TIANGEN,北京)提取总RNA,反转录采用全式金公司的RNA反转录试剂盒,采用qPCR Master Mix(SYBR Green)试剂盒进行Real-time PCR分析。通过NCBI设计的正向引物为5′-TGGAGGCTTTGGTCAGCTTT-3′,反向引物为5′-ACGTTGGTTCACGTTGCCTA-3′;小麦内参所用正向引物为5′-TGCTATCCTTCGTTTGGAC CTT-3′,反向引物为5′-AGCGGTTGTTGTGAGGGAG T-3′[21]。
2 结果
2.1 基因枪法线性转化片段
和线性转化片段如图1所示,由于转化片段中不含有任何载体骨架序列,最大程度地减少了由于载体骨架序列可能带来的安全风险;此外,由于和分别单独构制在2个线性转化片段中,为日后在转基因材料后代中筛选筛除的转基因株系提供了可能性。
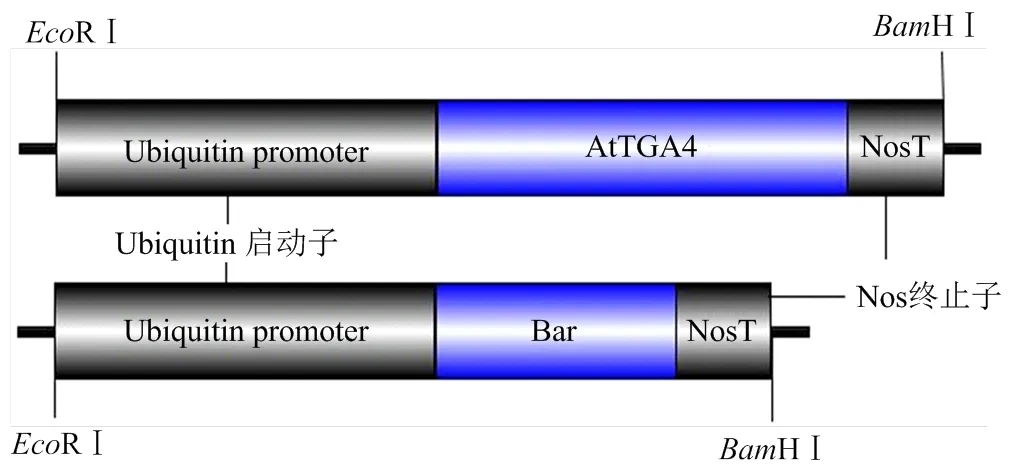
图1 线性转化片段示意图
2.2 转基因小麦的PCR检测
T4代转基因小麦PCR检测结果表明,已成功的整合受体石4056小麦基因组中,并稳定的遗传至T4代(图2);并没有检测到,说明在后代中发生了分离而被人为筛除,该转基因材料中只含有目的基因,而不含有筛选标记基因。
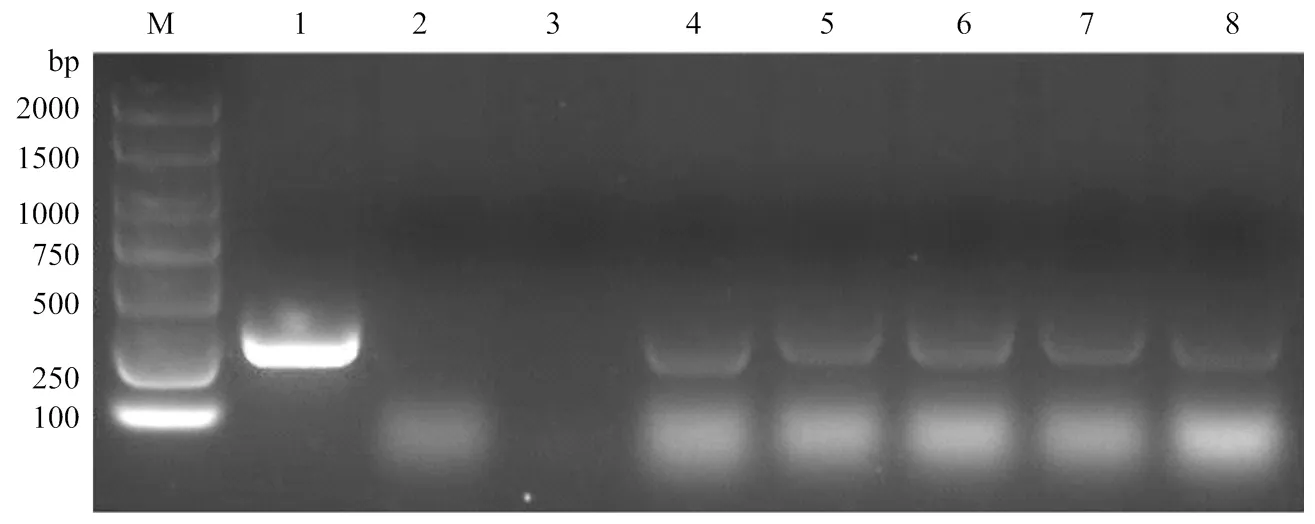
M:DNA Marker;1:阳性质粒对照;2:阴性受体对照;3:空白对照;4和5:株系L1;6—8:株系L2
2.3 试验地土壤养分含量测定结果及实际施肥情况
试验地土壤养分含量测定结果显示,正常和低磷处理土壤中有效氮、磷、钾养分的含量都比较一致。其中,正常和低磷处理中速效钾含量分别为256.39和254.54 mg·kg-1;碱解氮的含量则都偏高,分别为91.68和83.08 mg·kg-1,有效磷的含量则都偏低,分别为34.59和32.64 mg·kg-1。因此,在播种前分别施与适当的氮肥(尿素)和磷肥(过磷酸钙),实际施肥情况如下:正常处理地块施加过磷酸钙(812.39 kg·hm-2)和尿素(97.57 kg·hm-2);而低磷处理地块则只施加尿素(97.57 kg·hm-2)。
2.4 灌浆期田间表型
研究发现的2个转基因株系L1和L2在灌浆期相对于受体石4056表现出一定的生长优势。其中,株系L1的表型尤其明显(图3),在正常营养条件下,株系L1与受体长势基本一致;而在低磷处理条件下,受体明显比株系L1提早结束灌浆,主穗普遍提早变黄。

图3 灌浆期田间表型
2.5 低磷对转基因小麦产量及部分农艺性状的影响
从产量及部分农艺性状的统计结果(表1)可以看出,2个转基因株系在正常和低磷处理条件下的产量都显著高于受体,增幅在4.45%—8.60%;低磷对供试材料的株高没有明显的影响,但是普遍都降低了植株的分蘖和单穗结实率,表现在低磷处理条件下,供试材料的分蘖数和穗粒数都较正常营养条件下的低,分蘖减少9.84%—15.24%,穗粒数减少10.07%—13.38%;相反的,磷养分的缺少反而促进了供试材料籽粒重量的增加,表现在低磷胁迫条件下供试材料的千粒重都较正常条件下的要高,并且转基因株系L1和L2在低磷条件下的千粒重显著比受体提高8.10%—9.91%。该统计结果表明,低磷胁迫虽然促进了供试材料籽粒重量的增加,但由于普遍降低了其分蘖和单穗结实率,以致产量的下降;但由于2个转基因株系在不同处理条件下的分蘖和穗粒数与受体相当,籽粒重量比受体高,因而导致转基因材料在正常和低磷胁迫条件下的产量都要显著高于受体。
2.6 低磷对转基因小麦开花期部分生理指标的影响
如图4田间部分生理指标测定结果所示,在开花期,供试材料在低磷处理条件下的Fv/Fm都比正常条件下要低,降幅在8.40%—11.40%,正常营养条件下供试材料的Fv/Fm无明显差别;而低磷处理条件下,2个转基因株系L1和L2的光合速率都显著强于受体。开花期气冠温差测定结果表明,低磷条件下,2个转基因株系的CTD值都显著高于受体,表明转基因株系开花期体内水分代谢强度都要显著强于受体。然而开花期SPAD的测定结果却显示低磷胁迫对受体和转基因株系的叶绿素相对含量都没有表现出很明显的负面影响作用,受体和转基因株系之间的叶绿素相对含量也没有明显的差别。

表1 产量和部分农艺性状统计分析
*在0.01<<0.05时差异显著,**在<0.01时差异极显著。下同
* means significant difference at 0.01<<0.05, ** means extremely significant difference at<0.01. The same as below
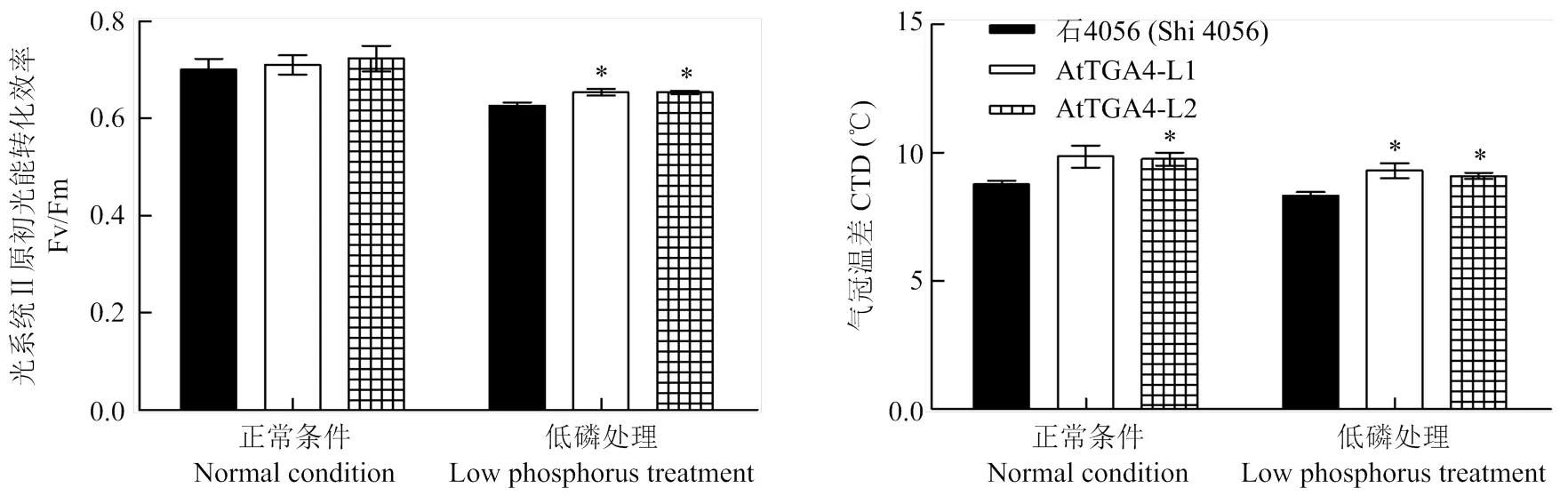
转基因株系与受体在相应营养条件下的显著性分析,*在0.01<P<0.05时差异显著。下同
2.7 低磷对转基因小麦不同组分磷含量的影响
成熟末期,供试材料不同组织部位磷浓度测定结果如图5所示,受低磷条件的影响,供试材料在低磷胁迫条件下的各组分磷浓度都有不同程度的下降。在低磷处理条件下,2个转基因株系的茎杆磷浓度都要显著高于受体,L1和L2株系的茎杆磷浓度是受体的1.37—1.52倍,株系L1的叶片磷含量和L2的籽粒磷浓度也较受体显著提高;正常条件下的则都无显著差异。
虽然转基因材料不同组分的磷浓度在除了茎杆以外的其他组织中相对于受体而言没有普遍的差异,但是成熟末期地上部各部位的磷吸收、残留总量统计结果(表2)却显示,2个转基因材料在茎杆、叶片中的磷残留总量在低磷胁迫条件下显著的高于受体,籽粒吸收的总磷含量在低磷条件下也显著的比受体高,而2个转基因材料在低磷营养胁迫条件下的茎杆、叶片和籽粒干重未发现有显著差异,这可能与作物开花后叶片、茎杆等组织中磷养分的再利用与分配有关[22-23]。结果表明,转基因小麦材料在低磷胁迫条件下地上部各部位对磷的保有能力都要显著高于受体石4056;低磷条件下,转基因小麦地上部整体总磷含量较受体增幅达6.38%—17.47%。
2.8 转基因材料AtTGA4的定量分析
为了解释2个转基因株系在应对低磷胁迫时的表现差异,通过qRT-PCR分析,结果(图6)显示,以株系L1中的表达量为标准,株系L2中目标基因的表达量要比株系L1的低,是株系L1中的0.69倍。
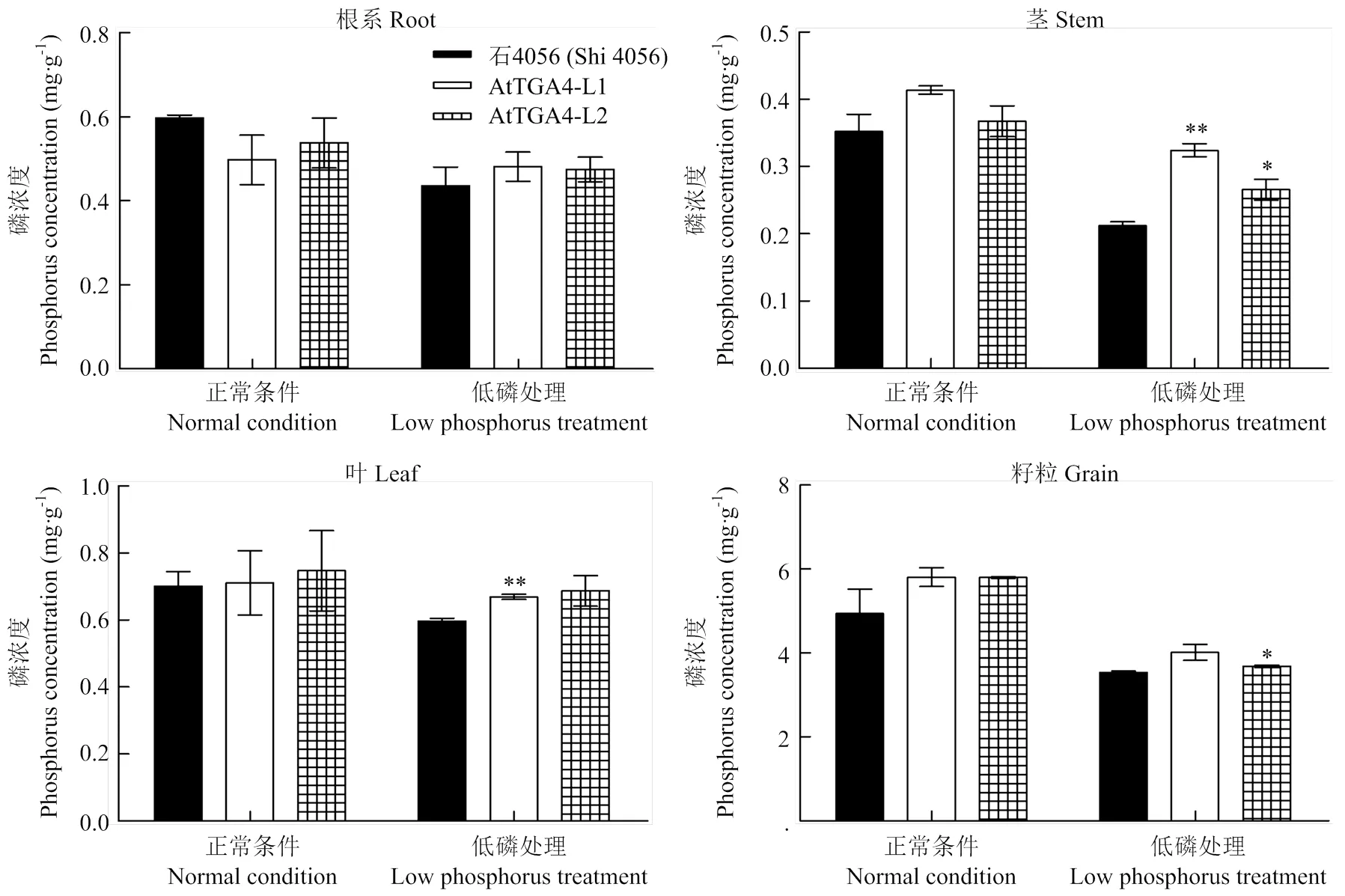
转基因株系与受体在相应营养条件下的显著性分析。**在P<0.01时差异显著
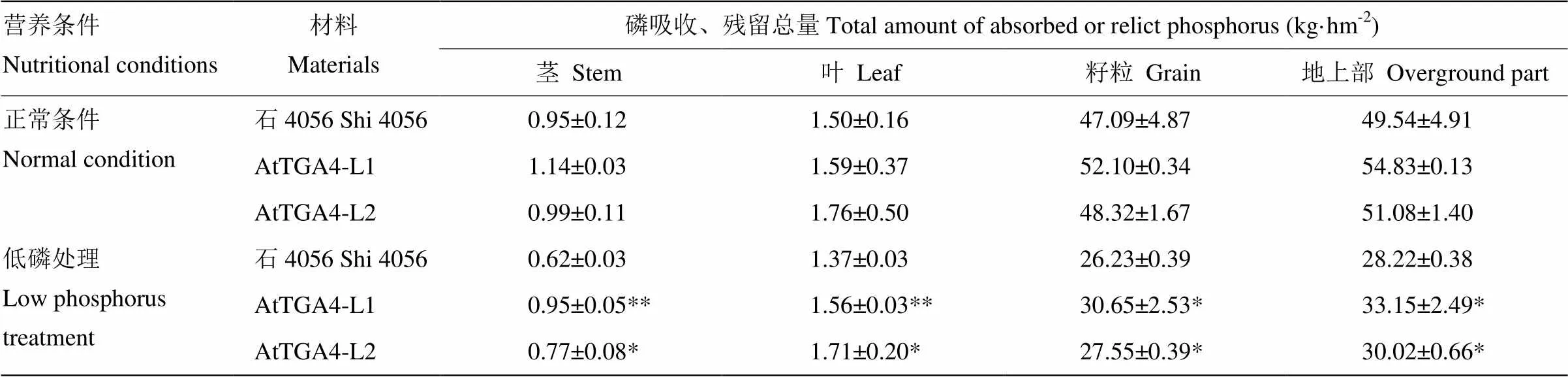
表2 各组分磷吸收、残留总量统计结果
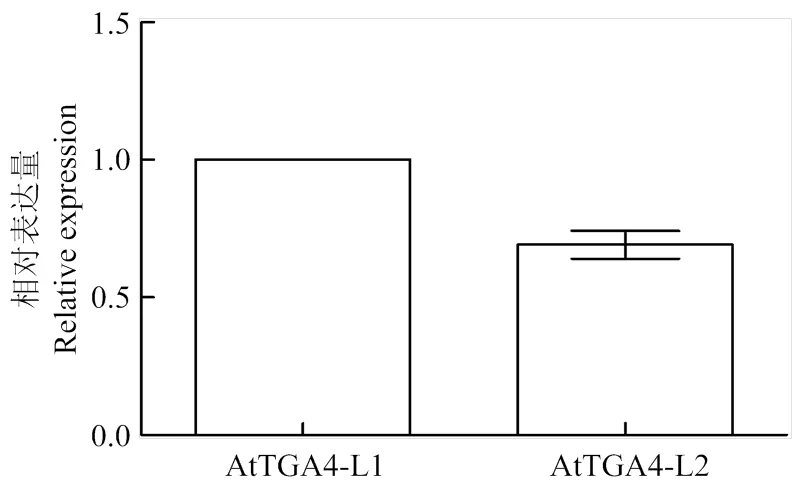
图6 转基因材料中AtTGA4的qRT-PCR分析
3 讨论
AtTGA4转录因子是bZIP转录因子家族成员之一,其亮氨酸拉链区能够特异的识别并结合TGACG调控序列,对靶基因进行转录激活/抑制的调节,参与植物抵御众多生物/非生物逆境胁迫的反应[24-26]。已有研究表明,不同物种、同一物种的不同基因型间对磷吸收和利用效率存在着遗传差异[8]。小麦属于异源六倍体植物,基因组庞大,遗传背景相对于玉米、水稻等其他作物要复杂得多[27]。在拟南芥中已经发现了一个以PHR为中心的调控网络,在水稻、小麦等植物中也发现了其同源基因。拟南芥AtPHR1能够识别并结合到其下游基因启动子的P1BS顺式作用元件上,调控其表达,参与植物应对低磷营养胁迫,而有研究表明,大多数磷饥饿响应基因的启动子区都拥有P1BS结构域[9,28-29]。植物一方面在不同的生长阶段对营养的需求有很大的不同,另一方面由于体内代谢的需要,养分在植物体内的分配与再利用也随着生长阶段的变化而在不断的调整中[30-31]。但是到了生殖生长后期,绝大多数一年生植物都会调动营养器官中的养分向生殖器官进行转运和再利用,以繁衍后代[32-33]。小麦与同源,研究发现,在小麦中过表达能够增强转基因小麦在低磷营养条件下对磷养分的吸收,并主要通过穗粒数的增加而致使增产,但是吸收的总磷量转化效率(磷收获指数)并没有提升[8];同样的,这种磷养分吸收的增加在和的过表达株系低磷胁迫研究中也出现过[34-35]。在本研究中,过表达转基因小麦株系在田间低磷营养胁迫条件下主要通过籽粒重量的增加而导致增产,这可能与转基因株系开花期较强的养分吸收和延长的灌浆期有关;此外,成熟期转基因株系低磷胁迫条件下地上部各部位磷吸收、残留总量都显著比受体要高,表明增强了小麦在低磷条件下对磷的吸收和磷养分由根系向地上部运输的强度,这与Nilsson[34]和Zhou[35]等关于PHR1类蛋白的研究结果相似。但是,在小麦应对低磷营养胁迫中发挥的作用又与不尽相同,在本研究中转基因小麦材料在正常和低磷营养条件下的分蘖和穗粒数都与受体无明显差异,反而是千粒重相对于受体得到了明显的增加。目前还没有数据证明与有关联。
土壤中磷养分的缺乏会降低植株叶片光合速率及各项叶绿素荧光参数(Fv/Fm,最大荧光等)[36],与本研究开花期Fv/Fm测定结果相同,供试材料低磷条件下的Fv/Fm都低于正常营养条件下的,但转基因株系在低磷条件下的Fv/Fm仍旧显著高于受体;而根系对土壤中养分的吸收、叶片光合作用对CO2的同化作用是植物体正常生长发育的基础,直接或间接的影响了作物产量的形成[37]。在光合作用中,光合色素对光能的吸收、转换和传递起着至关重要的作用,但是光合色素含量的高低与光合速率并无偶联关系[38]。本研究中,供试材料在开花期低磷条件下的SPAD与正常营养条件下的相当,同一营养条件下受体与转基因株系间的SPAD也无明显差异;此外,虽然CTD通常在评价植物抗旱强弱方面作为一个关键性指标,其往往反映的是植物从土壤中吸收有限水分进行体内各种生理代谢的能力,但是在土壤水分供应充足的情况下,通过测定CTD也能反映出不同植物、不同品系间体内生理代谢的强弱差异[39-40],在本研究中,转基因株系在开花期两种营养条件下的CTD都普遍高于受体,表明转基因株系在开花期午间时段的体内生理代谢水平要比受体强,有利于光合作用的碳同化。
此外,由于外源基因的插入位点、插入拷贝数以及受体自身转录翻译调控等原因,不同转基因株系在应对外界生物/非生物胁迫时会产生不同程度的响应。本研究中,转基因株系L1在正常和低磷营养条件下的亩产、籽粒/地上部磷吸收总量和不同组织的磷浓度表现都要普遍优于株系L2,这刚好与株系L1中较高的表达量相对应,表明株系L1在应对低磷胁迫时相对于株系L2在各方面变现出来的优越性可能与其较高的表达有关。本研究从表型、产量及农艺性状等方面初步分析了提高石4056小麦低磷胁迫耐受性的生理机制,相关的作用机理有待后续进一步的深入研究。
4 结论
在低磷胁迫条件下,转小麦产量及各项生理生化指标均有提升,显著提高了转基因小麦对低磷胁迫的耐性。
[1] Clarkson D T, Hanson J B. The mineral nutrition of higher plants., 1980, 31: 239-298.
[2] Schachtman D P, Reid R J, Ayling S M. Phosphorus uptake by plants: From soil to cell., 1998, 116: 447-453.
[3] 郭再华, 贺立源, 徐才国. 不同耐低磷水稻基因型秧苗对难溶性磷的吸收利用. 作物学报, 2005, 31(10): 1322-1327.
Guo Z H, He L Y, Xu C G. Uptake and use of sparingly soluble phosphorus by rice genotypes with different P-efficiency., 2005, 31(10): 1322-1327. (in Chinese)
[4] 李玉京, 李滨, 李继云, 李振声. 植物有效利用土壤磷特性的遗传学研究进展. 遗传, 1998, 20(3): 38-41.
Li Y J, Li B, Li J Y, Li Z S. Advances in genetic study on character of plant efficiently utilizing soil phosphorus., 1998, 20(3): 38-41. (in Chinese)
[5] 廖红, 严小龙. 菜豆根构型对低磷胁迫和适应性变化及其基因型差异. 植物学报, 2000, 42(2): 158-163.
Liao H, Yan X L. Adaptive changes and genotypic variation for root architecture of common bean in response to phosphorus deficiency., 2000, 42(2): 158-163. (in Chinese)
[6] Davies T G E, Ying J, Xu Q, Li Z S, Li J, Gordon-weeks R. Expression analysis of putative high-affinaty phosphate transporters in Chinese winter wheat., 2002(25): 1325-1340.
[7] Su J Y, Zheng Q, Li H W, Li B, Jing R L, Tong Y P, Li Z S. Detection of QTLs for phosphorus use efficiency in relation to agronomic performance of wheat grown under phosphorus sufficient and limited conditions., 2009(176): 824-836.
[8] Wang J, Sun J H, Miao J, Guo J K, Shi Z L, He M Q, Chen Y, Zhao X Q, Li B, Han F P, Tong Y P, Li Z S. A phosphate starvation response regulatoris involved in phosphate signalling and increases grain yield in wheat., 2013, 111: 1139-1153.
[9] Rubio V, Linhares F, Solano R, Martin A C, Iglesias J, Leyva A, Paz-Ares J. A conserved MYB transcription fator involved in phophate starvation signaling both in vascular plants and in unicellular algae., 2001, 15: 2122-2133.
[10] Devaiah B N, Karthikeyan A S, Raghotthama K G.transcription factor is a modulator of phosphate acquisition and root development in., 2007, 143: 1789-1801.
[11] Sano T, Nagata T. The possible involvement of a phosphate- induced transcription factor encoded bygene from tobacco in ABA signaling pathways., 2002, 43: 12-20.
[12] Yi K K, Wu Z C, Zhou J, Du L M, Guo L B, Wu Y R, Wu P., a novel transcription factor involved in tolerance to phosphate starvation in rice., 2005, 138: 2087-2096.
[13] Zhang J Y, Zhou X, Xu Y, Yao M L, Xie F B, Gai J Y, Li Y, Yang S P. Soybeanis an important component of the response to phosphate deficiency for phosphorus homeostasis., 2016, 248: 82-91.
[14] Zhong L, Chen D D, Min D H, Li W W, Xu Z S, Zhou Y B, Li L C, Chen M, Ma Y Z., a bZIP transcription factor, confers drought resistance by enhancing nitrate transport and assimilation in., 2015, 457: 433-439.
[15] 王娟, 孙爱平, 王开营, 沈永平, 张延成, 王锦贵. 土壤样品采集的原则与方法. 现代农业科技, 2011, 21: 300-301.
Wang J, Sun A P, Wang K Y, Shen Y P, Zhang Y C, Wang J G. Principles and methods to collect soil samples., 2011, 21: 300-301. (in Chinese)
[16] 郭胜利, 党延辉, 刘守赞, 郝明德. 磷素吸附特性演变及其与土壤磷素形态、土壤有机碳含量的关系. 植物营养与肥料学报, 2005, 11(1): 33-39.
Guo S L, Dang Y H, Liu S Z, Hao M D. Changes in characterization of phosphorus sorption in relation to it’s forms and soil organic carbon., 2005, 11(1): 33-39. (in Chinese)
[17] 宇万太, 张璐, 沈善敏, 廉鸿志. 辽西褐土施肥及养分循环再利用中长期试验Ⅲ. 磷和氮在堆腐过程中的循环率及有机肥料中养分利用率. 应用生态学报, 2002, 11(13): 1407-1409.
Yu W T, Zhang L, Shen S M, Lian H Z. A long-term field trial on fertilization and on use of recycled nutrients in farming systems Ⅲ. Recycling rate of P and N through a composting cycle and recoveries of P and N in organic manure., 2002, 11(13): 1407-1409. (in Chinese)
[18] 李科江, 张素芳, 贾文竹, 宋平忠, 刘树庆, 霍习良, 王玉朵. 半干旱区长期施肥对作物产量和土壤肥力的影响. 植物营养与肥料学报, 1999, 5(1): 21-25.
Li K J, Zhang S F, Jia W Z, Song P Z, Liu S Q, Huo X L, Wang Y D. Effect of long-term fertilization on crop yield and soil fertility in semi-arid area., 1999, 5(1): 21-25. (in Chinese)
[19] 王旭刚, 李建民, 张春霞. 氮磷配施对旱地小麦产量和吸肥特性的影响. 西北农林科技大学学报(自然科学版), 2007, 35(2): 138-142.
Wang X G, Li J M, Zhang C X. Effects of fertilizer N, P on wheat yield and nutrients uptake characters in dryland of Loess Plateau., 2007, 35(2): 138-142. (in Chinese)
[20] 赵亚妮. 不同氮磷和种植模式下小麦的生长动态、产量及养分效率研究[D]. 雅安: 四川农业大学, 2016.
Zhao Y N. Wheat growth dynamic, yield and nutrient use efficiency associated with N and P in different planting patterns[D]. Yaan: Sichuan Agricultural University, 2016. (in Chinese)
[21] 胡梦芸, 李辉, 庞建周, 刘茜, 张颖君, 孙丽静. 过量表达蔗糖转运蛋白基因增强转基因小麦的耐寒性. 中国农业科学, 2015, 48(8): 1473-1483.
Hu M Y, Li H, Pang J Z, Liu Q, Zhang Y J, Sun L J. Overexpression of sucrose transporter () improves drought tolerance in transgenic wheat., 2015, 48(8): 1473-1483. (in Chinese)
[22] Vance C P, Uhde-Stone C, Allen D L. Phosphorus acquisition and use: Critical adaptation by plants for securing a nonrenewable resource., 2003, 157: 423-447.
[23] 柴彦君, 熊又升, 黄丽, 袁家富, 徐祥玉, 刘晔. 施氮对不同品种冬小麦氮素积累和转运的影响. 西北植物学报, 2010, 10: 2040-2046.
Chai Y J, Xiong Y S, Huang L, Yuan J F, Xu X Y, Liu Y. Effects of nitrogen application on nitrogen accumulation, distribution and translocation of different winter wheat varieties., 2010, 10: 2040-2046. (in Chinese)
[24] Jakoby M. bZIP transcription factors in., 2002, 7(3): 106-111.
[25] 田义, 张彩霞, 康国栋, 李武兴, 张利益, 丛佩华. 植物TGA转录因子研究进展. 中国农业科学, 2016, 49(4): 632-642.
Tian Y, Zhang C X, Kang G D, Li W X, Zhang L Y, Cong P H. Progress on TGA transcription factors in plant., 2016, 49(4): 632-642. (in Chinese)
[26] Katagiri F, Lam E, Chua N H. Two tobacco DNA-bind proteins with homology to the nuclear factor CREB., 1989, 340: 723-730.
[27] 喻修道, 徐兆师, 陈明, 李连成, 马有志. 小麦转基因技术研究及其应用. 中国农业科学, 2010, 43(8): 1539-1553.
Yu X D, Xu Z S, Chen M, Li L C, Ma Y Z. The progress and application of wheat transformation technology., 2010, 43(8): 1539-1553. (in Chinese)
[28] Muller R, Morant M, Jarmer H, Nilsson L, Nielsen T H. Genome-wide analysis of theleaf transcription reveals interaction of phosphate and sugar metabolism., 2007, 143: 156-171.
[29] Nilsson L, Muller R, Nielsen T H. Dissecting the plant transcriptome and the regulatory responses to phosphate deprivation., 2010, 139: 129-143.
[30] Brant A N, Chen Y H Y. Patterns and mechanisms of nutrient resorption in plants., 2015, 34(5): 471-486.
[31] Freschet G T, Cornelissen J H C, Van Logtestijn R S P, Aerts R. Substantial nutrient resorption from leaves, stems and roots in a subarctic flora: what is the link with other resource economics traits?, 2010, 186(4): 879-889.
[32] Aerts R. Nutrient resorption from senescing leaves of perennials: are there general patterns?, 1996, 84(4): 597-608.
[33] Mao R, Song C C, Zhang X H, Wang X W, Zhang Z H. Response of leaf, sheath and stem nutrient resorption to 7 years of N addition in freshwater wetland of northeast China., 2013, 364(1): 385-394.
[34] Nilsson L, Muller R, Nielsen T H. Increased expression of the MYB-related transcription factor,, leads to enhanced phosphate uptake in., 2007, 30: 1499-1512.
[35] Zhou J, Jiao F C, Wu Z C, Li Y Y, Wang X M, He X W, Zhong W Q, Wu P.is involved in phosphate-starvation signaling and excessive phosphate accumulation in shoots of plants., 2008, 146: 1673-1686.
[36] Jacob J, Lawlor D W.photosynthetic electron transport does not limit photosynthetic capacity in phosphate-deficient sunflower and maize leaves., 1993, 16: 785-795.
[37] 许大全. 光合速率、光合效率与作物产量. 生物学通报, 1999, 34(8): 8-10.
Xu D Q. Photosynthetic rate, photosynthetic efficiency and crop yield., 1999, 34(8): 8-10. (in Chinese)
[38] 王菲, 曹翠玲. 磷水平对不同磷效率小麦叶绿素荧光参数的影响. 植物营养与肥料学报, 2010, 16(3): 758-762.
Wang F, Cao C L. Effects of phosphorus levels on chlorophyll fluorescence parameters of wheat (L.) with different phosphorus efficiencies., 2010, 16(3): 758-762. (in Chinese)
[39] 黄晓林, 李妍, 李国强. 冠层温度与作物水分状况关系研究进展. 安徽农业科学, 2009, 4: 1511-1515.
Huang X L, Li Y, Li G Q. Research advance in relationship between canopy temperature and crop water status., 2009, 4: 1511-1515. (in Chinese)
[40] 梁鸡保, 张元香, 王涛. 提高作物水分利用的研究. 北京农业, 2012, 27: 149.
Liang J B, Zhang Y X, Wang T. Study on improving water used of crop., 2012, 27:149. (in Chinese)
(责任编辑 李莉)
Functional Analysis ofTransgenic Wheat Tolerance to Low Phosphorus Stress in Field
Ling BingQi1, Bai XingXuan2, Zhou YongBin2, Wang ChunXiao2, Xu ZhaoShi2, Ma YouZhi2, Chen Ming2, Zhang XiaoHong1
(1College of Life Sciences, Northwest A & F University, Yangling 712100, Shaanxi;2Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081)
【Objective】In previous work, we had proven that a bZIP type transcription factor gene,can improve the drought resistance and low nitrogen tolerance of transgenic. In this study,was transformed into wheat to identify the tolerance of transgenic wheats to low phosphorus stress in field. At the same time, the physiological mechanism ofgene to improve the stress resistance of transgenic wheats was analyzed, which laid the foundation for the molecular breeding of wheat tolerant to low phosphorus stress. 【Method】Used particle bombardment method of minimal expression box to co-transformand marker geneinto wheat variety Shi4056. After transformation, we screened many transgenic wheat lines withoutgene and withgene through PCR assay. In field experiment, base on fertilizer nutrient content in soil we applied different levels of phosphate, which led to low phosphorus stress in soil.transgenic wheat lines were tested under low phosphorus. During the flowering stage some physiological indexes were identified such as light efficiency of the light system Ⅱ (Fv/Fm), the relative content of chlorophyll (SPAD) and crown the temperature difference (CTD), and other agronomic traits were investigated, during mature stage such as plants height, tiller number, grains per spike in mature period, and some values such as the grain yield and phosphorus concentration and phosphorus uptake in different components in wheat (root, stem, leaf and grain) were measured and data statistic analysis were completed. 【Result】PCR analysis showed thatgene had been stable heritability for T4generation in Shi4056 and four stable transgenic lines were obtained. According to the results of soil nutrient content, 812.39 kg·hm-2of superphosphate was applied to normal plots, and no phosphorus was applied in low phosphorus treatment plots. The statistical results of yield and agronomic characters showed that the grain yield oftransgenic lines L1 and L2 increased compared to wild type (WT) significantly under normal condition and low phosphorus stress. Under normal conditions the yield of transgenic wheat increase of 5.3%-8.6%, and under low phosphorus stress grain yield of transgenic wheat increased of 4.4%-7.7%. Thousand seed weight oftransgenic wheat increased significantly than WT. The results of physiological indexes assay in field showed that the Fv/Fm and CTD of transgenic lines L1 and L2 were significantly better than those of WT under the condition of low phosphorus, while SPAD had no significant difference. We found that under low phosphorus stress WT were in early grain filling than the transgenic wheat, and the ears turn yellow early in WT than that in transgenic wheat plants. The phosphorus content assay during the late mature stage showed that the phosphorus concentration in the stems of L1 and L2 of transgenic lines increased significantly compared with that of WT under the condition of low phosphorus, but there was no significant difference in other tissues. Under the condition of low phosphorus, the total phosphorus content in stems and leaves and grains of two transgenic lines was higher than that of WT, and the total phosphorus content in the aerial part increased by 6.38%-17.47%. qRT-PCR results oftransgenic wheat showed that the expression ofLine 2 (L1) was 0.69 times lower than that of Line 1 (L1). 【Conclusion】The above results showed that under the condition of low phosphorus stress,can improve the uptake and transportation of phosphorus in transgenic wheat plants, and increase the yield of transgenic plants, and further enhance the tolerance of transgenic wheat to low phosphorus stress.
transgenic wheat;; low phosphorus stress; yield; physiological index
2018-01-29;
2018-04-06
转基因生物重大专项(2016ZX08002-005)
凌炳琦,E-mail:370135413@qq.com。
张小红,E-mail:zhxh2493@126.com
10.3864/j.issn.0578-1752.2018.12.001