四川桤木7×7完全双列杂交种实、种苗性状的遗传效应分析
2018-06-23郭洪英杨汉波王泽亮李佳蔓肖兴翠康向阳
郭洪英 杨汉波 陈 炙 王泽亮 黄 振 李佳蔓 肖兴翠 康向阳
(1.林木分子设计育种高精尖创新中心/林木育种国家工程实验室/林木花卉遗传育种教育部重点实验室/城乡生态环境北京实验室,北京林业大学生物科学与技术学院,北京 100083; 2.四川省林业科学研究院,成都 610081)
四川桤木(AlnuscremastogyneBurk.)属桦木科(Betulaceae)桤木属(Alnus),是国产桤木属11个种中最为重要的一个特有种[1]。四川桤木能防止河水对堤岸的侵蚀,是重要的生态防护树种;其根系具有根瘤,能固定空气中游离的氮素增加土壤肥力;生长快,木材用途广,被列为优质造纸材用树种,是营造短周期工业用材原料林的重要树种之一[1~2]。总之,四川桤木是集速生用材、造纸材和生态防护等为一体的多功能树种,具有重要的生态和经济价值[1~3]。鉴于四川桤木重要的生态和经济价值,20世纪60年代以来,在湘、鄂、赣、浙等长江中下游地区广泛引种栽培[4]。陈益泰等[5]调查研究了四川盆地及周边21个不同产地天然林或人工林桤木的表型特征,并对桤木自然分布区进行了初步的划分。陈炳星等[6]就桤木商用种植的生产潜力、材性性状及制浆造纸研究现状进行了综合评述,指出开发利用桤木木浆具有广阔的前景。王军辉等[4,7]对桤木优良种源/家系遗传变异进行了研究,指出树高、材积和材性性状在种源间差异极显著,种源×环境的互作显著。周永丽等[8]对四川原产区的四川桤木材性进行了研究,发现四川桤木材性与生长性状可进行独立选择,并选出优良种源金堂-1,可进行优质制浆林定向培育推广试验。
完全双列杂交设计(Complete diallel cross design)是指一组亲本间进行所有可能的杂交[9]。完全双列杂交设计广泛应用于评估植物杂交育种配合力、杂种优势以及群体遗传结构[10~13]。配合力作为选配亲本的重要指标在林木遗传改良实践中发挥着十分重要的作用[14~16]。通过配合力的研究,可以估算亲本无性系的育种值,可为优良亲本选配提供依据,同时可揭示性状的遗传控制方式,用以科学指导有效的遗传改良方案和提高育种效率[17]。在马尾松(Pinusmassoniana)、杉木(Cunninghamialanceolata)、湿地松(Pinuselliottii)等重要用材树种的遗传改良进程中,配合力的估算发挥了重要的作用[17~19]。本研究采用Griffing完全双列杂交法[20~22],通过对四川桤木优树亲本7×7完全双列杂交种实及种苗性状的分析,估算四川桤木亲本的遗传参数,为四川桤木杂交亲本的选配及优良杂交子代苗期选择提供理论依据。
1 材料与方法
1.1 供试材料
选择7个优树作为杂交亲本(表1),2016年在四川省林业科学研究院唐昌基地按照完全双列交配设计(表2)进行控制授粉,每个杂交组合套4个袋,每个袋中5~15个雌花,人工授粉后套袋,待成熟后收获。
1.2 果实和种子性状的测定
根据杂交组合套袋将每个杂交组合分为4组,分别收获果实,测定4组内每个果实的鲜重、干重和单果出种数,分别用游标卡尺测量每个果实的果长、果宽和果柄长。每个杂交组合随机抽取900粒种子,分为3等份,每份300粒,分别混合称重,进而推算出千粒重,并利用体式显微镜分别对每份种子进行拍照,采用Image-pro-plus 6.0(IPP6.0)图像软件处理,并计算种子的表面积、种长和种宽。

表1 亲本来源
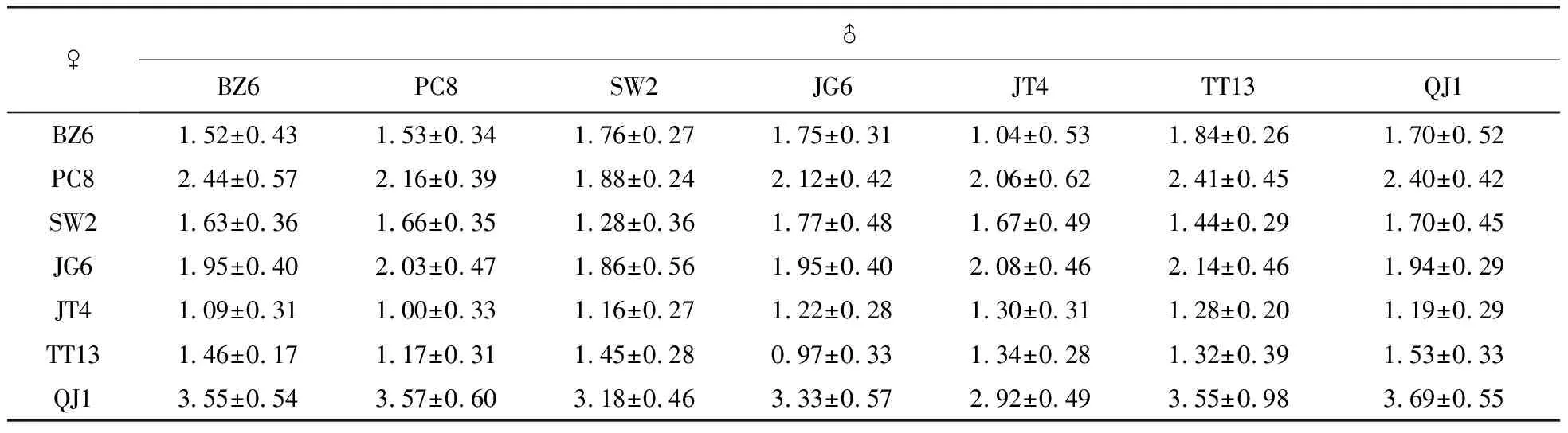
表2 四川桤木7×7完全双列杂交设计及各杂交组合获得的平均每个果穗鲜重
1.3 苗木生长调查
2016年3月,在四川省林业科学研究院唐昌基地进行营养杯育苗,试验采用完全随机区组设计,49个杂交组合,每个小区100杯,重复3次,定期浇灌。2017年8月,调查杂交子代的苗高和地径。
1.4 统计分析方法
采用SPSS 20.0统计软件对种实性状和种苗生长性状数据进行分析,对不同杂交组合间性状差异进行单因素方差分析。若遗传差异显著,则采用因子交配设计遗传分析软件WinNC2,按统计模型对一般配合力(GCA)、特殊配合力(SCA)、反交效应(REC)及遗传力进行估算:
yijklt=μ+Ei+Bij+Gk+Gl+Skl+GEik+GEil+SEikl+eijklt
(1)
式中:yijklt为第i个地点,第j个区组中第k个亲本与第l个亲本杂交后代的第t个个体的数量性状值;μ为总平均;Ei为第i个地点效应;Bij为第i个地点中第j个区组效应;Gk和Gl分别为第k个和第l个亲本的一般配合力;Skl为第k个与第l个亲本的特殊配合力;GEik和GEil为第i个地点与亲本一般配合力的交互效应;SEikl为第i个地点与亲本特殊配合力的交互效应;eijklt为随机误差[23]。
方差分析和配合力效应值估算采用固定模型,配合力方差分量和遗传力等遗传参数估算采用随机模型[24]。配合力稳定性则依据Kamble的稳定性比较法进行估算[25]。
遗传增益公式为:
(2)
式中:x为待评估杂交组合子代的平均值;X为所有测试子代的平均值[26]。
2 结果与分析
2.1 杂交组合间种实性状的差异显著性
通过对四川桤木完全双列杂交子代果实和种子性状的方差分析可以看出(表3),完全双列杂交子代的果实和种子性状、苗高、地径在不同组合间的差异均达到极显著水平。因此,各组合存在着显著的遗传差异,进行亲本间的选择是有效的。
分别对四川桤木各杂交组合的果实鲜重、种子千粒重、苗高和地径4个性状进行多重比较。结果表明,在果实鲜重上表现最优的组合是QJ1×QJ1和QJ1×PC8,其果实鲜重平均值分别为3.69 g和3.57 g(表4)。千粒重表现最优的组合为BZ6×TT13和QJ1×QJ1,分别为0.905和0.887 g,分别为最差组合的3.35和3.29倍。在果实鲜重和种子千粒重两性状上,自交组合QJ1×QJ1组合均处于较高的水平。49个杂交组合间的苗高和地径差异显著,苗高最优的组合是JT4×QJ1,其平均苗高为19.9 cm,为最差组合的4.7倍。地径最优的组合是BZ6×JG6和JT4×QJ1,平均地径分别为2.47和2.43 mm,分别为最差组合的2.6和2.5倍。7个自交组合杂交子代在苗高、地径上处于中下水平。
表3四川桤木种实和种苗各性状方差分析
Table3Varianceanalysisofthetraitsoffruits,seedsandseedlingsofA.cremastogyne

性状Traits自由度dfF值F valueP果实Fruit鲜重Fresh weight4850.543∗∗<0.001长度 Length4818.582∗∗<0.001宽度1 Width 1483.831∗∗<0.001宽度2 Width 24854.889∗∗<0.001果柄长 Carpopodium length4811.58∗∗<0.001种子Seed千粒重Thousand kernels weight4830.530∗∗<0.001面积 Area4838.672∗∗<0.001长度 Length4872.267∗∗<0.001宽度 Width4835.730∗∗<0.001苗高Seedling height4825.817∗∗<0.001地径Caliper4821.648∗∗<0.001
注:**在0.001水平上差异极显著,下同。
Note:**means highly significantly at the level of 0.001,the same as below.
表4四川桤木各组合果实鲜重、种子千粒重、苗高和地径多重比较分析
Table4Themultiplecomparisonanalysisoffreshweightoffruit,thousandkernelweightofseed,heightofseedling,andgrounddiameterbetweengroupsofA.cremastogyne

组合Groups果实鲜重Fresh weight of fruit千粒重Thousand kernel weight of seed苗高Height of seedling地径Ground diameter组合Groups果实鲜重Fresh weight of fruit千粒重Thousand kernel weight of seed苗高Height of seedling地径Ground diameterBZ6×BZ61.52defghijk0.57ghijklm5.2abc1.41cdefJG6×JT42.08nopqr0.63klmnopq12.46stuv2.13pqrsBZ6×PC81.53defghijkl0.75stuvw9.79klmnopq1.51cdefghJG6×TT132.14pqr0.52efghi12.21rstuv2.13pqrsBZ6×SW21.76hijklmnop0.71pqrstu11.23prqstu1.85lmnoJG6×QJ11.94lmnop0.55fghijk12.35rstuv2.19qrstBZ6×JG61.75hijklmnop0.67nopqrs13.57v2.47uJT4×BZ61.09abc0.54efghijk10.45mnoprqs1.71ghijklmnBZ6×JT41.04ab0.81vwxy10.04lmnoprq1.84klmnoJT4×PC81a0.52efghi13.67v2.08opqrBZ6×TT131.84ijklmnop0.91z11.69rqstuv1.79ijklmnJT4×SW21.16abcd0.56ghijkl11.11oprqst1.79ijklmnBZ6×QJ11.7ghijklmno0.84wxyz12.81tuv1.75hijklmnJT4×GJ61.22abcde0.46de10.59noprqst1.7fghijklmnPC8×BZ62.44r0.62jklmno5.93abcde1.43cdefJT4×JT41.3abcdefg0.67nopqrs10.73noprqst1.63defghijklmPC8×PC82.16pqr0.53efghij5.21abc1.74hijklmnJT4×TT131.28abcdef0.64lmnopq16.27w2.06opqrPC8×SW21.88klmnop0.65mnopq7.42cdefghijk1.42cdefJT4×QJ11.19abcd0.59hijklmn19.92x2.43tuPC8×JG62.12opqr0.74rstuv6.58bcdefg1.52cdefghiTT13×BZ61.46cdefghijk0.71pqrstu4.98ab1.09abPC8×JT42.06nopqr0.6ijklmn9.21hijklmnop1.84klmnoTT13×PC81.17abcd0.72qrstu6.4abcdefg1.36cdPC8×TT132.41qr0.68nopqrs7.76defghijkl1.38cdeTT13×SW21.45bcdefghij0.84xyz7.56cdefghijk1.56defghijkPC8×QJ12.4qr0.7opqrstu10.09lmnoprq1.74hijklmnTT13×JG60.97a0.78tuvwx7.35bcdefghij1.6defghijklSW2×BZ61.63efghijklm0.32ab9.63jklmnopq1.82jklmnoTT13×JT41.34abcdefgh0.67nopqrs6.9bcdefgh1.37cdSW2×PC81.66fghijklmn0.49defg7.41cdefghijk1.42cdefTT13×TT131.32abcdefg0.6ijklmn6.37abcdef1.45cdefgSW2×SW21.28abcdef0.27a5.4abcd1.44cdefgTT13×QJ11.53defghijkl0.75rstuv9.78klmnopq1.62defghijklSW2×JG61.77ijklmnop0.44cd7.39cdefghij1.97nopqQJ1×BZ63.55tu0.74rstuv8.19efghijklm1.48cdefghSW2×JT41.67fghijklmn0.36bc7.1bcdefghi1.65efghijklmQJ1×PC83.57u0.79uvwx4.25a1aSW2×TT131.44bcdefghi0.5defgh8.55fghijklmn1.91mnopQJ1×SW23.18st0.77tuvwx8.14efghijklm1.55defghijSW2×QJ11.7ghijklmno0.47def7.26bcdefghij1.69fghijklmnQJ1×JG63.33tu0.78uvwx5.39abcd1.37cdJG6×BZ61.95mnop0.63klmnop13.43uv2.28rstuQJ1×JT42.92s0.71pqrstu6.59bcdefg1.25bcJG6×PC82.03mnopq0.62jklmno8.8ghijklmno1.91mnopQJ1×TT133.55tu0.69opqrst9.34ijklmnopq1.59defghijklJG6×SW21.86jklmnop0.66mnopqr11.22prqstu2.37stuQJ1×QJ13.69u0.89yz7.86efghijkl0.97aJG6×JG61.95lmnop0.29ab10.96oprqst1.81jklmno
注:有相同字母的组合差异不显著;无相同字母的组合差异显著 下同。
Note:The groups with the same alphabet means no significant difference;the groups without the same alphabet means significant difference.The same as below.
2.2 一般配合力、特殊配合力和反交效应分析
在各性状差异显著的基础上,根据固定模型对各性状进行配合力分析(表5)。结果显示,果实鲜重、种子千粒重及其表型性状、苗高、地径的一般配合力(GCA)、特殊配合(SCA)和反交效应(REC)差异均达到了极显著水平。
表5四川桤木7×7全双列杂交配合力及反交效应值的方差分析
Table5Varianceanalysisofgeneticeffectsofcombiningabilityandspecificreciprocalcrossin7×7completediallelcrossofA.cremastogyne

性状TraitsGCASCAREC果实Fruit鲜重Fresh weight32.29∗∗∗∗1.83∗∗9.70∗∗长度Length13.17∗∗1.68∗∗3.54∗∗宽度1 Width 152.32∗∗2.04∗∗15.72∗∗宽度2 Width 238.26∗∗1.81∗∗12.12∗∗果柄长Carpopodium length32.53∗∗2.86∗∗13.60∗∗种子Seed千粒重Thousand kernels weight93.96∗∗15.61∗∗26.41∗∗表面积Area160.24∗∗7.32∗∗33.49∗∗长度Length153.21∗∗5.122∗∗43.45∗∗宽度Width75.83∗∗4.99∗∗18.71∗∗苗高Seedling height75.30∗∗17.39∗∗46.04∗∗地径Caliper73.58∗∗19.70∗∗30.25∗∗
在四川桤木7×7全双列杂交中,对果实鲜重、千粒重、苗高及地径进行各亲本GCA效应值多重比较分析(表6)。结果表明,不同的性状,各亲本GCA表现不同。QJ1、JG6和PC8的果实鲜重GCA效应值为正值,它们的果实鲜重高于群体平均值。QJ1、TT13、BZ6及PC8的千粒重GCA效应值为正,其中QJ1、TT13和BZ6与其它4个亲本的差异达到显著水平。亲本JG6、JT4、BZ6和QJ1的苗高GCA效应值为正,它们的子代表现均优于群体平均水平。地径的GCA效应值,JG6、JT4及SW2为正值,与其它亲本的差异均达到显著水平。综合分析以上结果,亲本JG6和JT4对苗高和地径生长影响较大,子代苗高GCA效应值分别为1.93和0.97,是GCA正效应值亲本QJ1的97和193倍;地径GCA效应值分别为0.26和0.10,是GCA正效应值亲本SW2的10和26倍。QJ1与JG6正好相反,其果实鲜重和种子千粒重GCA效应值均为正值,且均显著高于其它亲本,但QJ1的果实鲜重GCA极低,仅为0.01。BZ6千粒重和苗高GCA为正值,但在果实鲜重和地径GCA为负值;PC8果实鲜重和千粒重GCA为正值,但苗高、地径GCA为负值;TT13仅千粒重GCA为正值,SW2仅地径GCA为正值,其它均为负值。考虑到苗高、地径是苗期选择的主要表型依据,因此最终确定JG6和JT4为优良亲本。
表6四川桤木亲本的一般配合力差异显著性比较
Table6ComparisonofthedifferencesingeneralcombiningabilityofparentsofA.cremastogyne
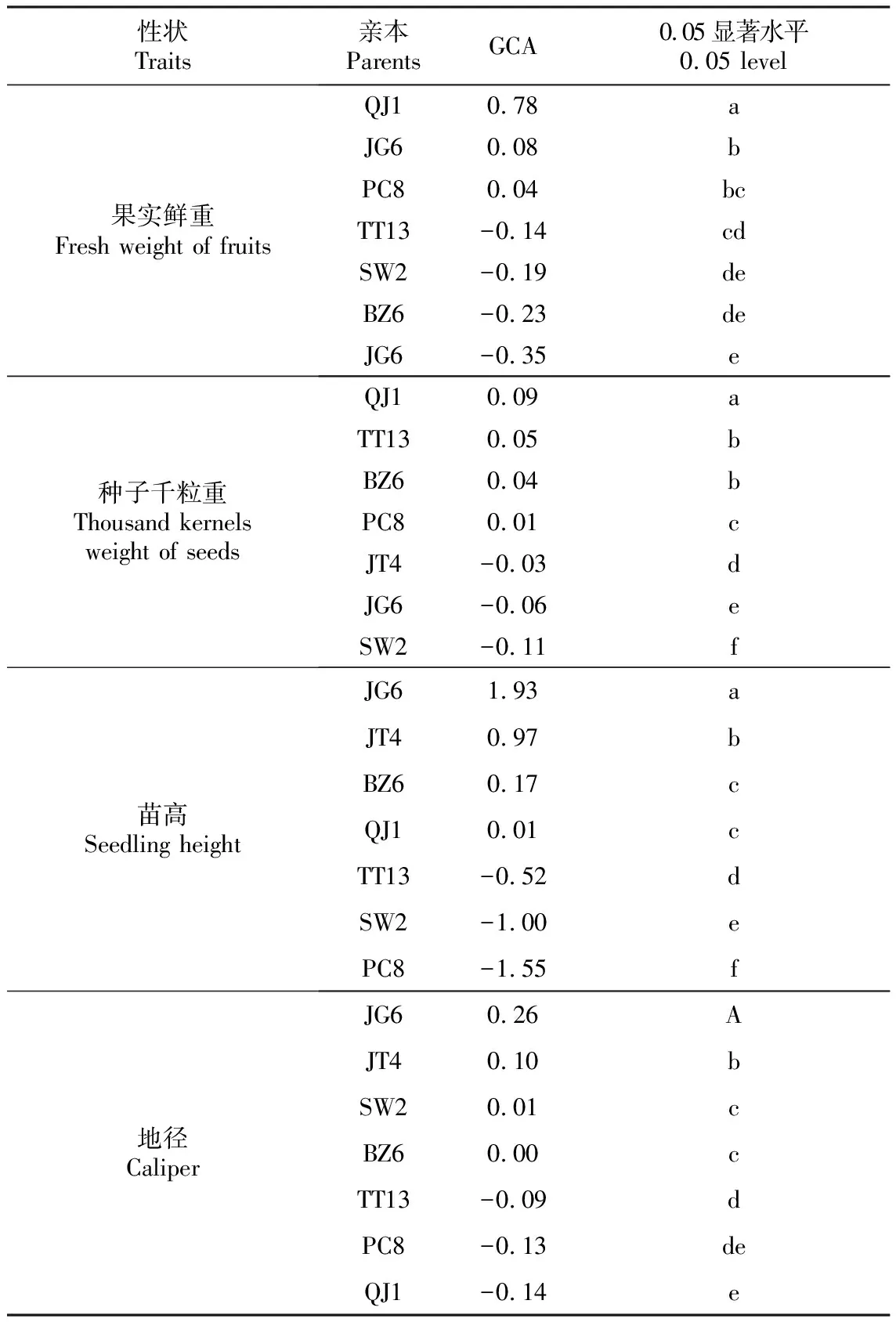
性状Traits亲本ParentsGCA0.05显著水平0.05 level果实鲜重Fresh weight of fruitsQJ10.78aJG60.08bPC80.04bcTT13-0.14cdSW2-0.19deBZ6-0.23deJG6-0.35e种子千粒重Thousand kernels weight of seedsQJ10.09aTT130.05bBZ60.04bPC80.01cJT4-0.03dJG6-0.06eSW2-0.11f苗高Seedling heightJG61.93aJT40.97bBZ60.17cQJ10.01cTT13-0.52dSW2-1.00ePC8-1.55f地径CaliperJG60.26AJT40.10bSW20.01cBZ60.00cTT13-0.09dPC8-0.13deQJ1-0.14e
特殊配合力(SCA)是评估杂交子代表现是否优于双亲的遗传指标。由表7可以看出,从果实鲜重SCA效应值大小分析,优良杂交组合为PC8×JG6(JG6×PC8)、TT13×QJ1(QJ1×TT13)、JG6×JT4(JT4×JG6)。从千粒重的SCA效应值分析,优良杂交组合为PC8×JG6(JG6×PC8)、SW2×TT13(TT13×SW2)、BZ6×TT13(TT13×BZ6)和SW2×JG6(JG6×SW2)。根据苗高SCA效应选出的优良杂交组合按照SCA效应大小依次为BZ6×JG6(JG6×BZ6)、PC8×JT4(JT4×PC8)、BZ6×SW2(SW2×BZ6)、JT4×QJ1(QJ1×JT4)、JT4×TT13(TT13×JT4)、BZ6×QJ1(BZ6×QJ1)。根据地径SCA效应值分析,优良组合依次为BZ6×JG6(JG6×BZ6)、PC8×JT4(JT4×PC8)、SW2×JG6(JG6×SW2)、TT13×QJ1(QJ1×TT13)、JT4×QJ1(QJ1×JT4)。

表7 四川桤木各组合的特殊配合力及反交效应值
注:奇数组合为正交组合,偶数组合为反交组合。
Note: Odd number combinations were orthogonal combinations,even number combinations were orthogonal combinations.
反交效应(REC)是选择优良组合中的正交组合还是反交组合的重要参考依据。在果实鲜重上,根据选出的3组杂交组合的REC效应值的正向还是负向效应值,确认优良杂交组合为JG6×PC8、QJ1×TT13和JG6×JT4。根据千粒重REC效应值确认优良杂交组合为PC8×JG6、TT13×SW2、BZ6×TT13和JG6×SW2。从苗高REC效应值的正向还是负向效应值,确认优良杂交组合为JG6×BZ6、JT4×PC8、BZ6×SW2、JT4×QJ1、JT4×TT13及BZ6×QJ1。根据地径的SCA和REC确定的优良组合有BZ6×JG6、JT4×PC8、JG6×SW2、TT13×QJ1和JT4×QJ1。
综合分析42个杂交组合在果实鲜重、种子千粒重、苗高和地径的SCA,组合JG6×JT4除在果实鲜重上SCA为正,表现较优,其它性状均为负值,表现不良而被淘汰。组合PC8×JG6和BZ6×TT13在果实鲜重和千粒重上SCA较高,苗高和地径SCA为负值,因此这些组合被淘汰。而组合PC8×JT4和JT4×QJ1正好相反,苗高和地径SCA为正值,果实鲜重和千粒重SCA为负值,因此这两个组也被淘汰。所以,最终确定的最佳杂交组合为BZ6×JG6、BZ6×SW2、JT4×TT13、BZ6×QJ1。
2.3 四川桤木种实、种苗性状的遗传控制方式
四川桤木7×7全双列杂交组合种实性状(果实鲜重、千粒重等)的加性方差明显大于非加性方差(表8)。果实鲜重、千粒重的加性方差分别是非加性方差的9.25和1.34倍,可见这两个性状均以加性基因效应控制为主,非加性基因效应控制次之,尤其是果实鲜重则主要由加性基因效应控制。种苗苗高、地径与之相反,非加性方差分别是加性方差的1.12和1.37倍,这两个性状均以非加性基因效应控制为主,加性基因效应控制次之。苗高和地径的遗传力均较高,广义遗传力分别为37.06%和38.04%,狭义遗传力分别为17.47%和16.03%,说明在苗期阶段,种苗生长性状受环境效应的影响较大。
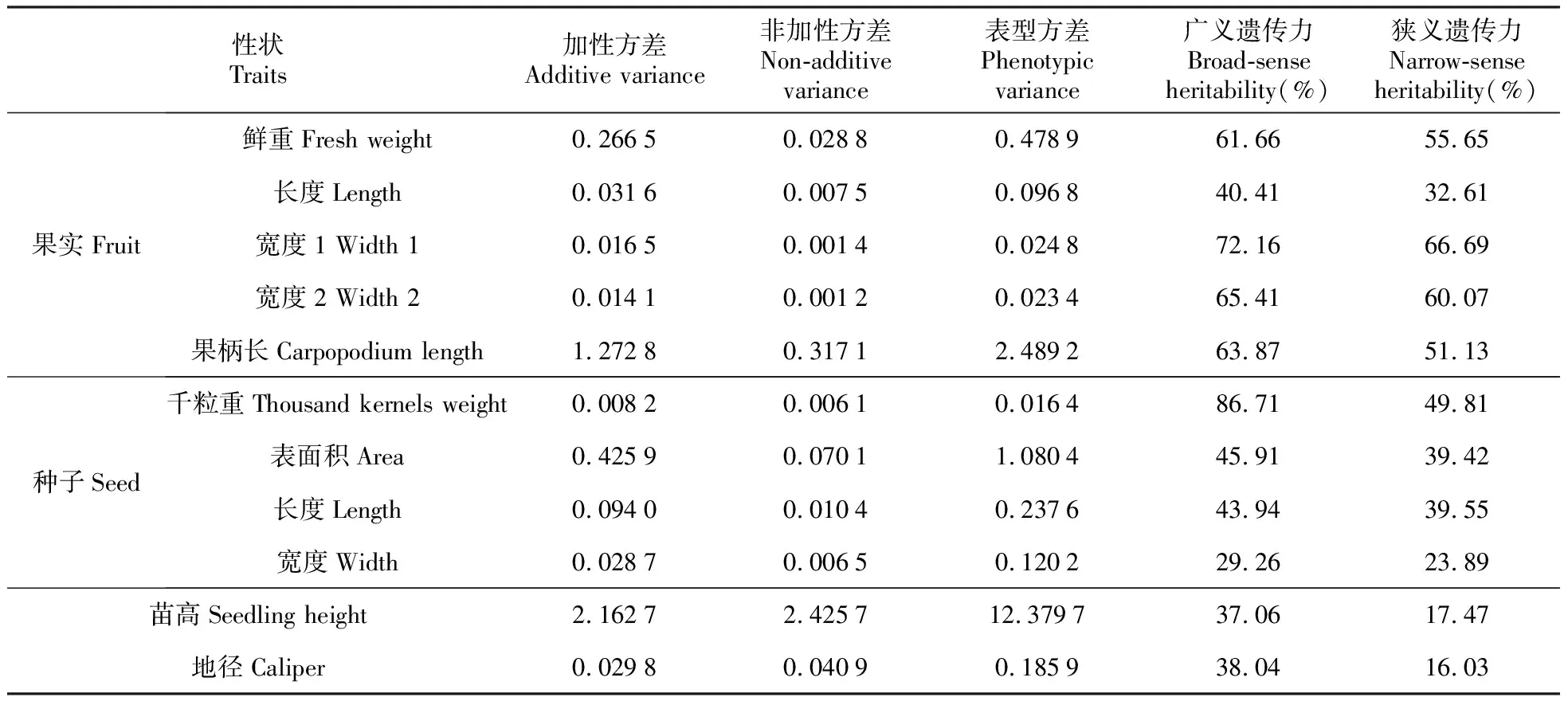
表8 四川桤木种实、种苗性状的主要遗传参数
2.4 优良杂交子代的评价
对依据SCA和REC效应值确定的7个最佳杂交组合,进行配合力组成分析,初步解析其优良性。因苗高和地径是苗期选择的主要表型依据,对这些杂交组合的苗高和地径进行配合力组成分析。JT4×TT13的苗高和地径均最大,BZ6×JG6次之,这两个组合均有一个亲本为确定的优良亲本,亲本间特殊配合力也较大,苗高分别实现了76.22%和44.44%的遗传增益,地径分别实现43.18%和23.13%的遗传增益。
3 讨论
一般配合力(GCA)由亲本基因的累加效应决定,一般配合力效应值的大小表明亲本对子代的影响程度,是亲本选择的重要依据之一[20,27]。本研究对四川桤木完全双列交配设计的种实和种苗性状的配合力分析结果显示,JG6和JT4的一般配合力效应值均显著高于其它亲本,被确定为优良亲本。特殊配合力(SCA)是基于杂交组合的显性、超显性和上位偏差所致,是选取优良杂交组合的依据之一[20,28]。根据特殊配合力和反交效应值确定苗期最佳杂交组合为BZ6×JG6、BZ6×SW2、JT4×TT13、BZ6×QJ1。在四川桤木49个杂交组合中,自交授粉的果实、种子性状与异交授粉无明显差异,甚至优于部分异交授粉组合,但其苗高、地径均处于较低的水平。据此认为,雌雄同株异花的四川桤木为自交亲和植物,其自交子代是可育的,但自交子代的生长发育性有所降低,不能用于生产制种。
本研究中,7个四川桤木亲本在果实鲜重、种子千粒重、苗高和地径4个性状上的一般配合力大小排序存在明显差异,如果实鲜重一般配合最高的为QJ1,但该亲本的苗高一般配合力极低,地径一般配合力则为负值。这与油茶(Camelliaoleifera)在树高、地径和冠幅3个性状上的一般配合力大小排序基本一致不同[26]。但与大多数树种配合力的表现相一致,如白桦(Betulaplatyphylla)[13,28]、马尾松(Pinusmassoniana)[29~30]、蓝桉(Eucalyptusglobulus)[31]等树种,亲本性状不同,其一般配合力表现均存在较大的差异。本研究的7个亲本中,JG6在苗高和地径上的一般配合力最高,说明其长势旺的优良性状可在杂交中最大程度的遗传给后代,因此可确定为优良亲本。加性效应是影响数量性状的多个微效基因的基因型值的累加,是性状表型值的主要成分[27]。而非加性效应是基因间交互作用所产生的效应的总称,叶志宏等[32]认为非加性效应主要是由母本细胞质效应引起的,而本身的效应是不显著的。本研究中观察到四川桤木果实鲜重、千粒重和苗高、地径均具有显著的GCA、SCA和REC效应,说明这些性状的遗传是由加性和非加性基因共同控制的。通过对11个性状的广义遗传力和狭义遗传力的分析,果实鲜重、种子千粒重、果实和种子表型性状的广义遗传力和狭义遗传力接近,加性方差明显高于非加性方差,说明这些性状主要是加性基因效应起的作用,非加性基因效应作用相对较小。进行性状配合力分析,揭示其基因作用方式,可正确有效地制定育种方案[33]。本研究中,四川桤木种实和种苗各性状的一般配合力和特殊配合力的相对重要性与玉米、杉木的研究结果不同[34~36],其可能原因为本试验所选的7个交配亲本来源于不同的地理种源区,遗传距离较大的优良无性系进行杂交有望产生配合力较强的杂交组合,在全部49个杂交组合中,绝大部分的组合代表了不同种源间的杂交。因此,对四川桤木的遗传改良不仅可通过建立种子园利用一般配合力效应,还可通过创制高特殊配合力的杂交组合,走有性制种无性规模扩繁的路径。遗传力作为亲本遗传某一性状给子代的能力,常根据它来预测选择的作用[37]。四川桤木苗高和地径的广义遗传力和狭义遗传力均处于较低的水平,表明苗高和地径的选择要根据试验结果来确定,不能仅仅根据亲本的生长性状表现来确定子代的生长性状。
四川桤木杂交子代各性状一般配合力和特殊配合力之间无明显的对应关系,如以苗高一般配合力最高的JG6与一般配合力较低的SW2和PC8杂交,两组合的特殊配合力差异很大,分别为0.057 5和-1.135 0,而以苗高一般配合力最高的JG6和JT4为亲本杂交,两组合间的特殊配合力却为负值。因此,四川桤木杂交育种中,根据亲本一般配合力筛选出优良亲本的前提下,还需要充分考虑亲本间的特殊配合力以确定最佳的亲本配置组合。本研究综合考虑一般配合力和特殊配合力初选出了优良杂交组合4个,杂交子代苗高和地径的遗传增益分别在20.47%~76.22%和5.07%~43.18%。
1.刘贤词,文仕知,冯汉华,等.四川桤木人工林不同年龄段生物量的研究[J].中南林业科技大学学报,2007,27(2):83-86.
Liu X C,Wen S Z,Feng H H,et al.Biomass study of the plantation ofAlnuscremastogyneBurkill at different stages of age[J].Journal of Central South University of Forestry & Technology,2007,27(2):83-86.
2.饶龙兵,杨汉波,郭洪英,等.桤木属7种植物的核型分析[J].西北植物学报,2013,33(7):1333-1338.
Rao L B,Yang H B,Guo H Y,et al.Karyotypes of seven species ofAlnusin European and American areas[J].Acta Botanica Boreall-Occidentalia Sinica,2013,33(7):1333-1338.
3.卓仁英.桤木群体遗传分化和抗涝性研究[D].北京:中国林业科学研究院,2003.
Zhuo R Y.Genetic differentitation between population and flooding tolerance ofAlnuscremastogyne[D].Beijing:Chinese Academy of Forestry,2003.
4.王军辉,顾万春,夏良放,等.桤木种源(群体)/家系材性性状的遗传变异[J].林业科学研究,2001,14(4):362-368.
Wang J H,Gu W C,Xia L F,et al.Genetic variation and selection of wood property ofAlnuscremastogyneprovenance and family[J].Forest Research,2001,14(4):362-368.
5.陈益泰,李桂英,王惠雄.桤木自然分布区内表型变异的研究[J].林业科学研究,1999,12(4):379-385.
Chen Y T,Li G Y,Wang H X.Study on phenotypic variation in natural range of long peduncled alder(Alnuscremastogyne)[J].Forest Research,1999,12(4):379-385.
6.陈炳星,周志春,李光荣,等.桤木制浆造纸研究现状与我国桤木浆的开发利用[J].林业科学研究,1999,12(6):656-661.
Chen B X,Zhou Z C,Li G R,et al.Status on pulping/papering research of alder and prospect of utilization of alder pulp wood in China[J].Forest Research,1999,12(6):656-661.
7.王军辉,顾万春,李斌,等.桤木优良种源/家系的选择研究——生长的适应性和遗传稳定性分析[J].林业科学,2000,36(3):59-66.
Wang J H,Gu W C,Li B,et al.Study on selection ofAlnuscremastogyneprovenance/family-analysis of growth adaptation and genetic stability[J].Scientia Silvae Sinicae,2000,36(3):59-66.
8.周永丽,刘福云,万军,等.四川桤木木材材性初步研究[J].四川林业科技,2003,24(1):75-78.
Zhou Y L,Liu F Y,Wan J,et al.A prelininary study of the wood property ofAlnuscremastogynein Sichuan[J].Journal of Sichuan Forestry Science and Technology,2003,24(1):75-78.
9.Jakubec V,Komender P,Nitter G,et al.Crossbreeding in farm animals.I.Analysis of complete diallel experiments by means of three models with application to poultry[J].Journal of Animal Breeding and Genetics,1987,104(1-5):283-294.
10.Sharma M K,Fanta S.Variance balanced designs for complete diallel cross[J].Journal of Agricultural Biotechnology and Sustainable Development,2010,2(4):56-60.
11.Bahari M,Rafii M Y,Saleh G B,et al.Combining ability analysis in complete diallel cross of watermelon(Citrulluslanatus(Thunb.) matsum.& nakai)[J].The Scientific World Journal,2012,2012:543158.
12.Moterle L M,De Lucca E Braccini A,Scapim C A,et al.Combining ability of popcorn lines for seed quality and agronomic traits[J].Euphytica,2012,185(3):337-347.
13.Zhao X Y,Bian X Y,Liu M R,et al.Analysis of genetic effects on a complete diallel cross test ofBetulaplatyphylla[J].Euphytica,2014,200(2):221-229.
14.Williams S M,Price S E,Siegel P B.Heterosis of growth and reproductive traits in fowl[J].Poultry Science,2002,81(8):1109-1112.
15.Saadey S M,Galal A,Zaky H I,et al.Diallel crossing analysis for body weight and egg production traits of two native egyptian and two exotic chicken breeds[J].International Journal of Poultry Science,2008,7(1):64-71.
16.Baril C P,Verhaegen D,Vigneron P,et al.Structure of the specific combining ability between two species of Eucalyptus.I.RAPD data[J].Theoretical and Applied Genetics,1997,94(6-7):796-803.
17.董虹妤.马尾松第3代种质幼林遗传分析及其亲本配合力研究[D].北京:中国林业科学研究院,2016.
Dong H Y.Genetic and combining ability analysis of the third generation germplasm ofPinusmassoniana[D].Beijing:Chinese Academy of Forestry,2016.
18.栾启福,姜景民,张建忠,等.火炬松×加勒比松F1代生长、树干通直度和基本密度遗传和配合力分析[J].林业科学,2011,47(3):178-183.
Luan Q F,Jiang J M,Zhang J Z,et al.Estimation of heritability and combining ability for growth,stem-straightness and wood density of the F1generation ofPinustaeda×P.caribaea[J].Scientia Silvae Sinicae,2011,47(3):178-183.
19.李力,陈孝丑,曹汉洋.杉木分组群状多系杂交子代测定林的遗传分析[J].江西农业大学学报,2000,22(3):388-393.
Li L,Chen X C,Cao H Y.Genetic analysis of progeny tests of nested polycross designs of Chinese Fir[J].Acta Agriculturae Universitatis Jiangxiensis,2000,22(3):388-393.
20.盛志廉,陈瑶生.数量遗传学[M].北京:科学出版社,1999.
Sheng Z L,Chen Y S.Quantative genetics[M].Beijing:Science Press,1999.
21.Hohls T,Shanahan P E,Clarke G P,et al.Genotype x environment interactions in a 10×10 diallel cross of quality protein maize(ZeamaysL.)[J].Euphytica,1995,84(3):209-218.
22.Topal A,Aydin C,Akgün N,et al.Diallel cross analysis in durum wheat(TriticumdurumDesf.):identification of best parents for some kernel physical features[J].Field Crops Research,2004,87(1):1-12.
23.童春发,蒋安纳,杨立委,等.WinNc2:因子交配设计遗传分析软件[J].林业科学,2014,50(1):55-62.
Tong C F,Jiang A N,Yang L W,et al.WinNc2:a new software for genetic analysis of factorial mating design[J].Scientia Silvae Sinicae,2014,50(1):55-62.
24.续九如.林木数量遗传学[M].北京:高等教育出版社,2006.
Xu J R.Quantitative genetics in forestry[M].Beijing:Higher Education Press,2006.
25.Kambal A E,Webster O J.Estimates of general and specific combining ability in grain sorghum,Sorghumvulgarepers.1[J].Crop Science,1965,5(6):521-523.
26.林萍,姚小华,滕建华,等.油茶5×5全双列杂交子代幼林生长性状的配合力分析[J].中南林业科技大学学报,2016,36(5):26-32.
Lin P,Yao X H,Teng J H,et al.Analysis of genetic effects of growth traits ofCamelliaoleiferaF1 descendants in complete diallel cross design Ⅲ[J].Journal of Central South University of Forestry & Technology,2016,36(5):26-32.
27.王明庥.林木遗传育种学[M].北京:中国林业出版社,2001.
Wang M X.Forest genetics and breeding[M].Beijing:China Forestry Publishing House,2001.
28.李开隆,姜静,姜莹,等.白桦5×5完全双列杂交种苗性状的遗传效应分析[J].北京林业大学学报,2006,28(4):82-87.
Li K L,Jiang J,Jiang Y,et al.Analysis of the genetic effects of seed and seedling traits ofBetulaplatyphyllain a 5×5 complete diallel cross design[J].Journal of Beijing Forestry University,2006,28(4):82-87.
29.刘青华,金国庆,储德裕,等.基于马尾松测交系子代的生长、干形和木材密度的配合力分析[J].南京林业大学学报:自然科学版,2011,35(2):8-14.
Liu Q H,Jin G Q,Chu D Y,et al.Genetic analysis on combining ability of growth,stem form and wood basic density ofPinusmassonianaby testcross mating design[J].Journal of Nanjing Forestry University:Natural Science Edition,2011,35(2):8-14.
30.刘青华,金国庆,王晖,等.马尾松巢式交配子代产脂力、生长和木材密度遗传分析[J].林业科学研究,2014,27(6):715-720.
Liu Q H,Jin G Q,Wang H,et al.Genetic analysis of resin-producing capability,growth and wood basic density on progeny of nested mating design ofPinusmassoniana[J].Forest Research,2014,27(6):715-720.
31.李淡清,刘永平,曾德贤,等.蓝桉6×6全双列交配生长性状的遗传效应分析[J].遗传学报,2002,29(9):835-840.
Li D Q,Liu Y P,Zeng D X,et al.Analysis of genetic effects for growth traits ofEucalyptusglobulusLabill.in a 6×6 diallel design[J].Acta Genetica Sinica,2002,29(9):835-840.
32.叶志宏,施季森,翁玉榛,等.杉木十一个亲本双列交配遗传分析[J].林业科学研究,1991,4(4):380-385.
Ye Z H,Shi J S,Weng Y Z,et al.Genetic analysis for eleven parent incomplete diallel of Chinese Fir[J].Forest Research,1991,4(4):380-385.
33.金国庆,秦国峰,刘伟宏,等.马尾松生长性状交配效应的遗传分析及杂交组合选择[J].林业科学,2008,44(6):28-33.
Jin G Q,Qin G F,Liu W H,et al.Effects of mating manners on growth traits ofPinusmassonianaand selection of cross combinations[J].Scientia Silvae Sinicae,2008,44(6):28-33.
34.齐明.杉木育种中GCA与SCA的相对重要性[J].林业科学研究,1996,9(5):498-503.
Qi M.Relative importance of GCA and SCA in genstic breeding of Chinses fir[J].Forest Research,1996,9(5):498-503.
35.李力,施季森,陈孝丑,等.杉木两水平双列杂交亲本配合力分析[J].南京林业大学学报,2000,24(5):9-13.
Li L,Shi J S,Chen X C,et al.Combining ability analyses of parents in two-level diallel cross experiment of Chinese fir[J].Journal of Nanjing Forestry University,2000,24(5):9-13.
36.周志春,金国庆,秦国峰,等.马尾松纸浆材重要经济性状配合力及杂种优势分析[J].林业科学,2004,40(4):52-57.
Zhou Z C,Jin G Q,Qin G F,et al.Analysis on combining ability and heterosis of main economic traits ofPinusmassonianafor pulp production[J].Scientia Silvae Sinicae,2004,40(4):52-57.
37.卢超,高明博,焦小钟,等.几个小麦亲本主要农艺性状的配合力评价及遗传力分析[J].麦类作物学报,2010,30(6):1023-1028.
Lu C,Gao M B,Jiao X Z,et al.Combining ability and heritability analysis of main agronomic traits in wheat[J].Journal of Triticeae Crops,2010,30(6):1023-1028.
