沙生植物沙鞭叶表皮微形态特征及其生态适应性研究
2018-06-23刘玉萍周勇辉张晓宇
吕 婷 刘玉萍 周勇辉 刘 涛 张晓宇 苏 旭*
(1.青海师范大学生命科学学院,西宁 810008; 2.青海省青藏高原药用动植物资源重点实验室,西宁 810008; 3.青海省自然地理与环境过程重点实验室,西宁 810008; 4.青藏高原环境与资源教育部重点实验室,西宁 810008)
营养叶是禾本科(Poaceae)植物中发育最为完善、表现最为充分、潜载信息量最大的一类叶性器官[1],尤为重要的是营养叶表皮层形态稳定、结构精细、类型多样,这些独有特征不仅使得营养叶表皮层的表皮结构细胞在禾本科植物类群划分中具有极其重要的分类价值,而且也可为禾本科植物系统亲缘关系分析提供有力旁证。20世纪30年代以来,诸多学者[1~9]曾先后将叶表皮解剖特征应用于禾本科植物的各级研究中,解决了经典分类难以处理的一些疑题,取得了令人瞩目的成效。譬如,我国学者蔡联炳等[1,6]通过对禾本科203属328种植物叶表皮微形态特征的研究,系统探讨了结构细胞的常见类型和演化趋势,认为结构细胞在类群中呈现的类型、形态、数量及分布式样等对禾本科植物的鉴定划分和亲缘关系探讨具有重要意义;同时,依据结构细胞在类群间呈现的差异,显示的类型数、类型演化水平和变异趋势,将禾本科划分为5亚科、3超族,并对各大类群进行了系统位置的评认、亲缘关系的探讨和演化途径的推导。马海英等[10]通过对国产5种拂子茅属和26种、1变种野青茅属植物叶表皮微形态特征的观察,发现广义拂子茅属的叶表皮特征为典型的狐茅型,许多结构在种间具有丰富的变异式样,认为这种分异与物种的海拔分布有关,而与该属属下系统相关性不大。新近,李兆孟等[11]采用常规叶表皮解剖法对国产赖草属24个种、72个代表种群叶表皮微形态性状特征的观测,发现叶表皮微观差异性状可将赖草属鉴分为多穗组、少穗组和单穗组,同时依据演化关系和系统位置分析认为多穗组植物最原始、少穗组植物较进化、单穗组植物最高级,多穗组可能直接派生了较进化的少穗组,并在少穗组基础上进而产生了最高级的单穗组。
然而,从调查的情况和报道的资料来看,先前未见对任何荒漠禾本科植物进行过叶表皮微形态解剖特征方面的研究报道,这对于类群庞大的禾本科来说,其论述实体显得明显不足。因此,研究更多的类群,阐明禾本科植物的叶表皮解剖特征及其对生态环境的适应性仍然是植物形态解剖学研究的主要任务;尤其是选择荒漠地区的特有物种来进行这方面的研究无疑可能会取得令人满意的结果。根据我们长期的野外调查研究,发现沙生植物沙鞭是进行这一研究的最佳代表。沙鞭(Psammochloavillosa(Trin.) Bor)是禾本科(Poaceae)针茅族(Stipeae)沙鞭属(Psammochloa)的一种多年生、沙生草本植物,主要分布于我国内蒙古高原及其毗邻地区,具有较强的耐旱、耐寒、耐盐碱以及抗病和抗风沙能力,是草原带、荒漠草原带以及荒漠地区流动沙丘上的特有先锋植物和建群草本物种,也是这些极端沙地环境分布的关键或唯一草本物种[12~15],因而它不仅具有重要的经济和生态价值,而且也是农牧业上禾草良种繁育和牧草利用的重要基因资源。鉴于此,本文利用光学显微镜对沙生植物沙鞭50个代表种群的叶表皮微形态特征进行了观测和研究,对结构细胞的分类价值、演化趋势以及生态适应性进行了较为全面的分析,以期为禾本科其它沙生植物的叶表皮微观特征以及对干旱环境生态适应性探讨提供微形态学证据。
1 材料与方法
1.1 实验材料
实验材料全部来源于野外采集的新鲜叶片,除个别代表种群选取2~4份叶片外,其余所有代表种群均选取6份材料进行实验,共307份。同时,取材时尽量选取不同经纬度和海拔高度的叶片进行实验研究。此外,沙鞭因在叶片上表皮性状特征上差异较小,故本文重点放在其叶片下表皮性状特征的差异比较上。所有凭证标本均保存于青海师范大学青藏高原植物标本室。沙鞭种群叶表皮解剖材料的详细地理分布采样点如图1所示。

图1 沙鞭地理分布图Fig.1 Sampling localities of P.villosa surveyed in the Inner Mongolian Plateau
1.2 实验方法
用于叶表皮微形态观察的材料均取自花序下第2叶片的中间部段,共50个沙鞭代表种群。按常规叶表皮解剖法操作,即先将叶片水煮30~45 min,待材料软化后置于塑料板上用单面刀片轻轻刮削,使其保留叶片上、下表皮薄膜层,然后用清水洗净薄膜,用1%的番红染色40 s,再置于光学显微镜下观察,同时进行描述、绘图、拍照等工作,最后以常规永久制片法脱水封固保存。长细胞测其绝对长度,每个代表种群取3个视野,每个视野取10个细胞,取平均值;气孔器大小测其长宽比,每个代表种群取3个以上视野,每个视野取5个气孔器,取平均值。叶表皮特征用Metcalfe[3]的微形态术语进行描述。
2 实验结果
光学显微镜下观察了沙鞭50个代表种群叶表皮的微形态结构,发现它们皆由长细胞、短细胞、气孔器细胞和刺毛4类细胞组成(图版Ⅰ~Ⅱ)。长细胞普遍分布于各个代表种群中,通常呈长方形,纵向相接成行,各行平行排列于叶片的脉上或脉间,一般脉上着生4~16列,脉间着生3~14列,长94~197 μm,宽12~32 μm,长细胞细胞壁薄而平直或厚而密波状弯曲;气孔器细胞为椭圆形,带状分布于脉间两侧或正中,通常1~2列,长45~75 μm,宽35~62 μm,副卫细胞平边形或低圆顶形;短细胞单生或孪生,稀不着生,镶嵌于长细胞间,呈新月形、马鞍形或圆球形;刺毛数目相对较少,可生于脉上和脉间,脉上多为刺,脉间多为钩(图版Ⅰ~Ⅱ)。在各个种群中,这几类结构细胞的形态、数量和叶面的组合式样是不尽相同的,尤其是在叶片下表皮长细胞的长短与宽窄、壁的波曲程度,短细胞着生方式,气孔器密度大小及副卫细胞形状,刺毛分布形式,以及脉带与脉间带的相对宽窄上是存在差异的(表1),这些差异可将参试的所有种群基本划分为两个大的群体。故下面着重将两个集群下表皮的差异特征概总如下:集群Ⅰ:长细胞短阔,细胞壁常波状弯曲;短细胞单生,呈马鞍形,圆球形和新月形;气孔器密度相对较密集,副卫细胞低圆顶形或近平边形;脉带宽于或等于脉间带。集群Ⅱ:长细胞狭长,细胞壁常平直;短细胞常孪生,呈马鞍形,圆球形和新月形;气孔器密度相对较稀疏,副卫细胞低圆顶形或近平边形;脉带明显窄于脉间带。
表1沙鞭两个集群间叶片下表皮微形态特征比较
Table1ComparisonofmicromorphologicalcharacteristicsaboutthelowerepidermisesbetweentwogroupsinP.villosa
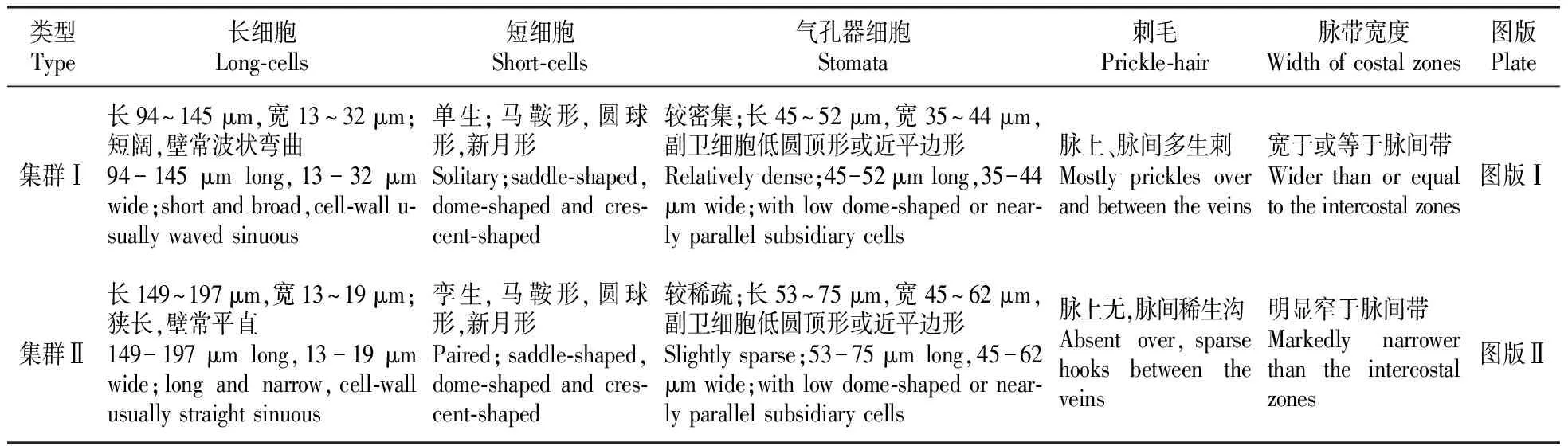
类型Type长细胞Long-cells短细胞Short-cells气孔器细胞Stomata刺毛Prickle-hair脉带宽度Width of costal zones图版Plate集群Ⅰ长94~145μm,宽13~32μm;短阔,壁常波状弯曲94-145μm long,13-32μm wide;short and broad,cell-wall u-sually waved sinuous单生;马鞍形,圆球形,新月形Solitary;saddle-shaped,dome-shaped and cres-cent-shaped较密集;长45~52μm,宽35~44μm,副卫细胞低圆顶形或近平边形Relatively dense;45-52μm long,35-44μm wide;with low dome-shaped or near-ly parallel subsidiary cells脉上、脉间多生刺Mostly prickles over and between the veins宽于或等于脉间带Wider than or equal to the intercostal zones图版Ⅰ集群Ⅱ长149~197μm,宽13~19μm;狭长,壁常平直149-197μm long,13-19μm wide;long and narrow,cell-wall usually straight sinuous孪生,马鞍形,圆球形,新月形Paired;saddle-shaped,dome-shaped and cres-cent-shaped较稀疏;长53~75μm,宽45~62μm,副卫细胞低圆顶形或近平边形Slightly sparse;53-75μm long,45-62μm wide;with low dome-shaped or near-ly parallel subsidiary cells脉上无,脉间稀生沟Absent over,sparse hooks between the veins明显窄于脉间带Markedly narrower than the intercostal zones图版Ⅱ
3 讨论
沙鞭50个代表种群叶表皮微形态解剖结果表明,所有参试个体的叶片下表皮仅由长细胞、短细胞、气孔器细胞和刺毛4类结构细胞所组成,缺乏明显的大毛、微毛和乳突这些表皮细胞附属物,特别是缺乏微毛和乳突的组合特征与先前学者[2~4,6~7]解剖过的其他禾本科植物的叶表皮结构一样隶属于典型的狐茅型(Festucoid type)结构,同时这些结构成分在叶表面的分布和着生情况也跟陈文俐[16]报道的针茅族(Stipa)野青茅属(Deyeuxia)植物的叶表皮基本一致,因而进一步说明其应属于狐茅型中针茅族型叶表皮。正如观察结果所述,沙鞭代表种群的叶片下表皮在长细胞形状和壁的波曲程度、短细胞类型、气孔器密度大小和副卫细胞形态、刺毛分布以及脉带与脉间带相对宽窄等性状上存在较为明显差异,这些性状差异能将参试的所有种群基本划分为两个大的群体。即一个是由内蒙古高原西部的沙鞭种群组成,其下表皮长细胞短阔,具马鞍形、圆球形和新月形短细胞,气孔器密度相对较密集,副卫细胞呈低圆顶形或近平边形,脉带宽于或等于脉间带;另一个是由内蒙古高原东部的沙鞭种群组成,其下表皮长细胞狭长,具马鞍形、圆球形和新月形短细胞,气孔器密度相对较稀疏,副卫细胞呈低圆顶形或近平边形,脉带明显窄于脉间带。印证在外部形态上,则第一群体植株高大,茎秆粗壮,叶片渐狭、先端常内卷,圆锥花序退化成总状花序,外稃略宽,花药较短;第二群体植株较矮,茎秆较细,叶片宽阔、较平展,圆锥花序疏松开展,外稃较宽阔,花药相对较长。因此,我们认为沙鞭种群的叶表皮微形态解剖结构对于其集群划分具有十分重要的意义,完全可以作为该属不同生态集群类型划分的重要参考依据。
目前,国内外学者已经对沙鞭克隆基株及分株种群特征[17]、克隆生长对切断根茎处理反应[18]、等位酶变异和克隆多样性[19]、克隆生长在沙地斑块动态中的作用[20]、种子生物学特性[21]、种子萌发生态学[22~23]、根围AM真菌空间分布及其与土壤因子的关系[24]等进行了研究。然而,迄今为止仍未见关于内蒙古高原及其毗邻地区、不同来源沙鞭种群叶表皮微形态解剖特征生态适应性的比较研究。先前研究表明,内蒙古高原的光辐射和气温由东向西逐渐增加,而年降水量由东向西则逐渐减少,从而形成光照、温度和湿度的明显梯度以及多种多样的水热组合形式,依次可以划分为湿润、半湿润偏湿、半湿润偏干、半干旱偏润、半干旱偏干、干旱、很干旱、强干旱和极干旱9个水分型[25]。其中,水分是植物生长发育影响最大的生态因子。据此,本研究通过比较内蒙古高原不同干旱环境条件下沙鞭种群叶表皮微形态解剖特征的差异,以此来阐述它对干旱胁迫的生态适应性。研究表明,在长期干旱环境条件胁迫下,旱生植物特别是荒漠植物将会产生一系列适应这种特殊逆境生态因子的特定形态、解剖和生理特征,以此来抵御或忍耐这种干旱胁迫环境对它的伤害,进而产生耐旱性[26~29]。尤其是植物叶片上分布的表皮毛和气孔器对干旱胁迫生态因子响应最为强烈,它们的数量和密度与干旱胁迫环境因子强度呈现明显的正相关关系[30~32]。本研究结果显示,内蒙古高原的沙鞭种群叶表皮微形态解剖结构随分布地域不同呈现规律性变化,主要体现为具有抗旱适应能力的叶表皮微形态特征的变化上,即分布于内蒙古高原西部的沙鞭种群,其气孔器体积越小、数目越多,并且下陷表皮细胞之下越严重(图版Ⅰ);而内蒙古高原东部的沙鞭种群气孔器体积相对较大、数目相对较多,并且下陷表皮细胞之下相对较弱(图版Ⅱ)。王勋陵和马骥[33]曾认为,气孔数目多反映植物具有较高的光合作用速率,而光合作用速率的提高是植物抵抗干旱环境胁迫的重要方式之一,这与我们对沙鞭可以适应不同干旱环境胁迫条件的论点是完全一致的。不仅如此,我们还认为气孔器在控制植物蒸腾作用方面应具有双重作用,通常水分充足时气孔打开,促进植物进行蒸腾作用并防止过热灼伤;当水分缺乏时气孔关闭,将会抑制植物发生蒸腾作用。因此,小而数目多的气孔器应是沙鞭抵抗不良干旱环境的重要措施和手段,它通过这些叶表皮微形态特征的不同而产生具有差异的抗旱性。所以,本研究认为荒漠植物沙鞭具有的上述叶表皮微形态解剖结构特征与其呈现出来的外部形态特征和生长状况相一致,其主要通过“保护型”适应对策来抵御和适应不同程度的干旱胁迫环境,总之,我们认为研究荒漠植物沙鞭的叶表皮微形态特征及其生态适应性,不仅能够为以后沙鞭的抗旱生理研究提供必要的解剖学证据,而且对其将来种质资源的综合评价及引种驯化和栽培都具有十分重要的指导意义。
1.蔡联炳,王世金,李建华.中国主要禾本科植物叶片表皮细胞、表皮附属物的常见类型及其分类价值的初步探讨[C].高原生物学集刊,1991(10):1-14.
Cai L B,Wang S J,LI J H.Primary study on the classification value and the ordinary type of the attachment for the leaf cell epidermal is of major Poaceae in China [C].The Journal Polymer Published in Plateau Biology,1991,(10):1-14.
2.Prat H.La systématique des Graminées[J].Ann Sci Nat Bot,1936,10(18):165-258.
3.Metcalfe C R.Anatomy of the monocotyledons:Volume 1 Gramineae[M].London:Oxford Clarendon Press,1960:1-713.
4.Clifford H T,Watson L.Identifying grasses:data,methods and illustrations[M].Brisbane:University of Queensland Press,1977:1-141.
5.陈守良,金岳杏,吴竹君,等.叶表皮细胞结构在国产狗尾草属(SetariaBeauv.)分组水平上的应用[J].植物研究,1985,5(2):105-112.
Chen S L,Jin Y X,Wu Z J,et al.The study leaf epidermis at the sectional level on ChineseSetaria[J].Bulletin of Botanical Research,1985,5(2):105-112.
6.蔡联炳,郭延平.禾本科植物叶片表皮结构细胞主要类型的演化与系统分类和发育途径的探讨[J].西北植物学报,1995,15(4):323-335.
Cai L B,Guo Y P.Studies on constituent cells of leaf epidermis,systematics and phylogenetic path of the family Poaceae[J].Acta Botanica Boreali-Occidentalia Sinica,1995,15(4):323-335.
7.蔡联炳,郭延平.禾本科植物叶片表皮结构细胞主要类型的演化与系统分类和发育途径的探讨(续)[J].西北植物学报,1996,16(1):65-72.
Cai L B,Guo Y P.Studies on constituent cells of leaf epidermis,systematics and phylogenetic path of the family Poaceae(Continued)[J].Acta Botanica Boreali-Occidentalia Sinica,1996,16(1):65-72.
8.蔡联炳.禾本科叶片表皮结构细胞的组合式样及其分类学意义[J].植物研究,1999,19(4):415-427.
Cai L B.Combinative patterns of constituent cells of leaf epidermis of Poaceae and their taxonomic significance[J].Bulletin of Botanical Research,1999,19(4):415-427.
9.苏旭,刘玉萍,李兆孟,等.基于形态和叶表皮微形态特征试论西藏固沙草的归并[J].植物研究,2015,35(3):321-326.
Su X,Liu Y P,Li Z M,et al.Reduction onOrinustibeticusbased on the external morphology and micromorphological characteristics of leaf epidermis[J].Bulletin of Botanical Research,2015,35(3):321-326.
10.马海英,彭华,王跃华.禾本科广义拂子茅属的叶表皮形态研究[J].植物分类学报,2006,44(4):371-392.
Ma H Y,Peng H,Wang Y H.Morphology of leaf epidermis ofCalamagrostiss.l.(Poaceae: Pooideae) in China[J].Acta Phytotaxonomica Sinica,2006,44(4):371-392.
11.李兆孟,周勇辉,刘玉萍,等.基于形态和叶表皮微形态特征试论国产赖草属属下类群间的系统关系[J].植物研究,2016,36(1):10-25.
Li Z M,Zhou Y H,Liu Y P,et al.Biosystematic relationships among taxa belowLeymus(Poaceae) from China based on the external morphology and micromorphological characteristics of leaf epidermis[J].Bulletin of Botanical Research,2016,36(1):10-25.
12.刘瑛心.中国沙漠植物志:第1卷[M].北京:科学出版社,1985:34.
Liu Y X.Flora in desertis reipublicae populorum sinarum(Tomus 1)[M].Beijing:Science Press,1985:34.
13.郭本兆.中国植物志:第9卷[M].北京:科学出版社,1987:309.
Kuo P C.Flora Reipublicae Popularis Sinicae(Tomus 9)[M].Beijing:Science Press,1987:309.
14.马毓泉.内蒙古植物志:第5卷[M].2版.呼和浩特:内蒙古人民出版社,1994:115-152.
Ma Y Q.Flora Inner Mongolica(Tomus 5)[M].2nd ed.Hohhot:Inner Mongolia People’s Publishing House,1994:115-152.
15.Wu Z Y,Raven P H,Hong D Y,et al.Flora of China(Vol.22)[M].Beijing:Science Press and St.Louis:Missouri Botanical Garden Press,2006.
16.陈文俐.中国野青茅属的分类学修订[D].北京:中国科学院植物研究所,2001.
Chen W L.Revision of the genusDeyeuxia(Poaceae) in China[D].Beijing:The Institute of Botany,Chinese Academy of Sciences,2001.
17.董鸣,阿拉腾宝,邢雪荣,等.根茎禾草沙鞭的克隆基株及分株种群特征[J].植物生态学报,1999,23(4):302-310.
Dong M,Alateng B,Xing X R,et al.Genet features and ramet population features in the rhizomatous grass speciesPsammochloavillosa[J].Acta Phytoecologica Sinica,1999,23(4):302-310.
18.董鸣.切断根茎对根茎禾草沙鞭和赖草克隆生长的影响[J].植物学报,1999,41(2):194-198.
Dong M.Effects of severing rhizome on clonal growth in rhizomatous grass speciesPsammochloavillosaandLeymussecalinus[J].Acta Botanica Sinica,1999,41(2):194-198.
19.王可青,葛颂,董鸣.根茎禾草沙鞭的等位酶变异及克隆多样性[J].植物学报,1999,41(5):537-540.
Wang K Q,Ge S,Dong M.Allozyme variance and clonal diversity in the rhizomatous grassPsammochloavillosa(Gramineae)[J].Acta Botanica Sinica,1999,41(5):537-540.
20.陈玉福,于飞海,张称意,等.根茎禾草沙鞭的克隆生长在毛乌素沙地斑块动态中的作用[J].生态学报,2001,21(11):1745-1750.
Chen Y F,Yu F H,Zhang C Y,et al.Role of clonal growth of the rehizomatous grassPsammochloavillosain patch dynamics of Mu Us sandy land[J].Acta Ecologica Sinica,2001,21(11):1745-1750.
21.黄振英.鄂尔多斯高原固沙禾草沙鞭种子休眠和萌发与环境的关系[J].西北植物学报,2003,23(7):1128-1133.
Huang Z Y.Adaptation strategies of seed dormancy and germination ofPsammochloavillosa,a sand dune grass inhabiting Ordos Plateau,China[J].Acta Botanica Boreali-Occidentalia Sinica,2003,23(7):1128-1133.
22.黄振英,董鸣,张淑敏.沙鞭(禾本科)种子在沙丘上的萌发策略及幼苗的耐干燥特性[J].生态学报,2005,25(2):298-303.
Huang Z Y,Dong M,Zhang S M.Strategies of seed germination on sand dune and seedling desiccation tolerance,ofPsammochloavillosa(Poaceae)[J].Acta Ecologica Sinica,2005,25(2):298-303.
23.朱雅娟,董鸣,黄振英.沙埋和种子大小对固沙禾草沙鞭的种子萌发与幼苗出土的影响[J].植物生态学报,2005,29(5):730-739.
Zhu Y J,Dong M,Huang Z Y.Effects of sand burial and seed size on seed germination and seedling emergence ofPsammochloavillosa[J].Acta Phytoecologica Sinica,2005,29(5):730-739.
24.徐翥骍,贺学礼,郭辉娟,等.内蒙古农牧交错区沙鞭和羊柴AM真菌侵染及其土壤因子[J].河北农业大学学报,2011,34(1):56-61.
Xu Z X,He X L,Guo H J,et al.AM fungi colonization and soil factors ofPsammochloavillosa(Trin.) Bor andHedysalumlaeveMin.in farming-pastoral area of Inner Mongolia[J].Journal of Agricultural University of Hebei,2011,34(1):56-61.
25.马成仓,高玉葆,王金龙,等.内蒙古高原甘蒙锦鸡儿光合作用和水分代谢的生态适应性研究[J].植物生态学报,2004,28(3):305-311.
Ma C C,Gao Y B,Wang J L,et al.Ecological adaptation ofCaraganaopulenson the inner Mongolia Plateau:photosynthesis and water metabolism[J].Acta Phytoecologica Sinica,2004,28(3):305-311.
26.周云龙.植物生物学[M].2版.北京:高等教育出版社,2004.
Zhou Y L.Plant biology[M].2nd ed.Beijing:Higher Education Press,2004.
27.李长复.叶子表面的适应性能[J].生物学通报,1985(9):2+5-7.
Li C F.Adaptability of leaf surface[J].Bulletin of Biology,1985(9):2+5-7.
28.李芳兰,包维楷.植物叶片形态解剖结构对环境变化的响应与适应[J].植物学通报,2005,22(S1):118-127.
Li F L,Bao W K.Responses of the morphological and anatomical structure of the plant leaf to environmental change[J].Chinese Bulletin of Botany,2005,22(S1):118-127.
29.刘玉冰,李新荣,李蒙蒙,等.中国干旱半干旱区荒漠植物叶片(或同化枝)表皮微形态特征[J].植物生态学报,2016,40(11):1189-1207.
Liu Y B,Li X R,Li M M,et al.Leaf(or assimilation branch) epidermal micromorphology of desert plant in arid and semiarid areas of China[J].Chinese Journal of Plant Ecology,2016,40(11):1189-1207.
30.Barthlott W.Epidermal and seed surface characters of plants:systematic applicability and some evolutionary aspects[J].Nordic Journal of Botany,1981,1(3):345-355.
31.Casson S A,Hetherington A M.Environmental regulation of stomatal development[J].Current Opinion in Plant Biology,2010,13(1):90-95.
32.Ghahremaninejad F,Khalili Z,Maassoumi A A,et al.Leaf epidermal features ofSalixspecies(Salicaceae) and their systematic significance[J].American Journal of Botany,2012,99(4):769-777.
33.王勋陵,马骥.从旱生植物叶结构探讨其生态适应的多样性[J].生态学报,1999,19(6):787-792.
Wang X L,Ma J.A study on leaf structure and the diversity of xerophytes ecology adaptation[J].Acta Ecologica Sinica,1999,19(6):787-792.

图版Ⅰ 内蒙古高原西部沙鞭叶片下表皮光学显微照片 A.脉带;B.脉间带;C.低圆顶副卫细胞;D.平顶副卫细胞;E.长细胞;F.马鞍形短细胞;G.新月形短细胞;H.圆球形短细胞PlateⅠ Light microscopic graphs of the lower epidermises of leaf blades of P.villosa in the western Inner Mongolian Plateau A.Costal zones; B.Intercostal zones; C.Low dome-shaped subsidiary cells; D.Parallel subsidiary cells; E.Long-cells; F.Saddle-shaped short-cells; G.Crescent-shaped short-cells; H.Round short-cells

图版Ⅱ 内蒙古高原东部沙鞭叶片下表皮光学显微照片 A.长细胞;B.平顶副卫细胞;C.低圆顶副卫细胞;D.圆球形短细胞;E.马鞍形短细胞;F.新月形短细胞Plate Ⅱ Light microscopic graphs of the lower epidermises of leaf blades of P.villosa in the eastern Inner Mongolian Plateau A.Long-cells; B.Parallel subsidiary cells; C.Low dome-shaped subsidiary cells; D.Round short-cells; E.Saddle-shaped short-cells; F.Crescent-shaped short-cells
