富含OPO的植物乳脂粉与肠道微生物体外发酵特性研究
2018-06-20刘蔚宇叶子青李琳琳侯爱香李宗军
刘蔚宇,叶子青,李琳琳,侯爱香,李宗军
(湖南农业大学食品科学技术学院 长沙410128)
0 引言
1,3-二油酸-2-棕榈酸甘油三酯,简称OPO,是猪油和母乳脂的主要三甘酯成分之一,其棕榈酸主要分布于Sn-2位。OPO作为营养补充剂,能在人体消化时不易形成钙皂,更易于脂肪酸和钙的消化吸收,目前多添加在婴幼儿配方奶粉中,是婴幼儿配方奶粉母乳化过程中的关键添加成分,对婴幼儿健康成长有至关重要的作用[1-3]。但国内外婴幼儿配方奶粉中添加的多是结构脂质OPO,这种结构脂质OPO产品,通常以两种形式添加到婴幼儿配方奶粉中,一是制成富含OPO的植物乳脂油,湿法加入奶粉中,二是制成粉剂营养补充剂,即富含OPO的植物乳脂粉,干法加入奶粉中,目前,国内多采用干法加入的方式。
人体肠道内约寄生着450种肠道微生物,它们大多具有促进肠道蠕动,帮助人体新陈代谢,预防感染和自体免疫疾病的功能,另有一些特殊菌群甚至还能够控制人体对癌症治疗药物的反应[4]。由此可见,肠道菌群是我们人体必不可少的一部分,有些研究者将其称之为我们身体的一个“器官”[5],在人体粪便中,这些肠道微生物的数量能高达1011~1012cfu/g[6]。人体肠道菌群的数量与种类随着年龄的增长而改变,有研究表明,成人体内的肠道菌群相较于婴幼儿时期的体内肠道菌群更具多样性和稳定性[7]。肠道微生物已成为人们研究膳食因子营养机制的重要载体,其与膳食因子的互作效应也成了研究热点。但大部分传统研究膳食与肠道微生物关系的方式是动物试验,容易引发道德和伦理争议,且试验周期长,操作复杂,不利于科学试验的高效开展。而研究者同时发现,肠道微生物体外厌氧发酵和肠道内发酵在细菌数量、组成、多样性和代谢产物等方面都相似,因此本研究采用静态间歇式体外厌氧发酵模式,研究富含OPO的植物乳脂粉与肠道菌群的体外发酵特性。
多年监测表明:我国青年、少年身体素质呈持续下降趋势[8]。青年大学生是国家的人才主力,但很大一部分大学生由于远离家庭的监管,饮食和作息不规律,同时学习竞争激烈,就业和深造压力大,身体素质大不如前,已经引起社会的广泛关注。目前,营养补充剂结构脂质OPO应用范围相对较小,为进一步拓宽OPO的应用范围,提高大学生的身体素质,本研究以健康普通大学生的新鲜粪便为菌源,从膳食因子对肠道微生态影响的角度入手,研究不同时期(0,4,8,12,24 h)原料对肠道菌群的影响,并计算益生元指数和B/E值,测定发酵环境的pH值,为结构脂质OPO产品的进一步推广提供基础数据,为大学生营养食品的开发提供前期研究。
1 材料与方法
1.1 材料
1.1.1样品
植物乳脂粉,含有50%植物精炼油(含有40%OPO的棕榈油);4.21%蛋白质;41.63%乳糖;3.72%水;0.44%稳定剂;乳化剂;矿物质;维生素和其他物质成分。
1.1.2 培养基
(1)含氮基础培养基:1L纯净水含有蛋白胨2 g,酵母粉 2 g,N aC l 0.l g,K2HPO40.04 g,M gSO4·7H2O 0.01 g,CaC l2·7H2O 0.01g,N aHCO32 g,氯化血红素0.05 g,L-半胱氨酸0.5 g,胆汁酸盐0.5 g,吐温-80 2 m L,VK 10μL,0.025 g/100m L刃天青4m L。
(2)选择性培养基:营养琼脂用于计数总需氧菌[9];W ilkins and Chalgren琼脂用于计数总厌氧菌[10];Beerens琼脂用于计数双歧杆菌[11];R ogosa培养基用于计数乳酸杆菌[12];EC培养基用于计数肠球菌[13];Sulfite-polym yxin-m ilk琼脂用于计数梭状芽孢杆菌[14];紫红胆盐葡萄糖琼脂用于计数肠杆菌科菌;拟杆菌矿物盐琼脂(自制)用于计数拟杆菌[15]。拟杆菌矿物盐琼脂按配方自制:葡萄糖15.0 g,磷酸二氢钾4.0 g/L,磷酸氢二钠2.0 g/L,硫酸铵0.5 g/L,氯化钠9.0g/L,氯化镁0.15 g/L,氯化钙0.01 g/L,氯化锰0.05 g/L,氯化钴0.05 g/L,半胱氨酸0.8 g/L,碳酸氢钠1.5 g/L,氯高铁血红素0.01 g/L,VB120.005 g/L,七水合硫酸铁0.001 g/L,萘啶酮酸0.01 g/L,万古霉素0.0025 g/L,琼脂20.0 g/L。(除特殊规定外,本标准所用试剂均为分析纯,水为蒸馏水或相应纯净水。)
1.2 主要试剂和仪器
稀释液(0.9%生理盐水);粪便取样器,压舌板,一次性手套,镊子,牛皮纸,自封袋,EP管离心管,试剂瓶(采样工具提前灭菌,部分塑料取样用具再用DEPC水浸泡24 h以上);电热恒温水浴锅HH-8(上海浦东物理光学仪器厂),分析天平TP-213(北京赛多利斯仪器系统有限公司),高压蒸气灭菌锅SP500(日本YAMATO),超净工作台SW-CJ-2D(苏州净化设备有限公司),YQX-I厌氧培养箱(上海跃进医疗器械厂),恒温生化培养箱SPX-250BS-II,pH计Testo205(Testo AG)。
1.3 试验方法
1.3.1 样品采集及准备
以3个健康大学生(女,年龄20~22岁)的粪便作为发酵菌种,要求实验对象1个月内未服用抗生素,且无肠胃病史。采集每位试验者的新鲜粪便50g于粪便取样管中,存入-80℃冰箱备用。将3名志愿者的粪样混合,然后将50 g混合粪样稀释到500 m L冷却后的稀释液(0.9%生理盐水)中,加几粒无菌玻璃珠涡旋15 s使粪便充分分散。稀释的粪液用4层无菌纱布过滤,并将过滤得到的粪便悬浮液密封于一无菌器皿中。
1.3.2 肠道菌群富集培养
将上述含氮基础培养基各配置450m L分装于两个试剂瓶中(标记A组,B组),灭菌备用。在A组中加入67.5 g的植物乳脂粉作为实验组(根据国家食品安全标准GB1488-2012年食品营养补充使用标准,婴幼儿配方奶粉OPO的添加量为24~96 g/kg,根据奶粉冲泡的一般稀释:4.5 g奶粉/30m L水,制备发酵液A。本研究发酵液A以50 g/kg的添加量乘以配方奶粉稀释比,使OPO初始浓度为质量浓度0.75%,B组为含氮基础培养基,作为空白对照。各吸取50 m L粪便悬浮液加入发酵液A和B中,即接种量为10%,37℃厌氧发酵。
1.3.3 发酵液的分段收集及检测
在发酵0,4,8,12,24 h时,将A组与B组分别吸取5个1m L液体分装在含有9m L发酵液的EP管中密封,混匀后放入-80℃冰箱保存,备用。采用10倍梯度稀释法稀释发酵液(稀释液为0.9%生理盐水),选取各梯度稀释液100μL均匀涂布8个肠道微生物指标的不同的选择培养基上,从取样至涂布结束在2 h内完成。涂布后总需氧菌、乳酸杆菌、肠球菌、肠杆菌科在37℃需氧培养24~48 h后分别计数,总厌氧菌、双歧杆菌、拟杆菌和梭状芽孢杆菌在37℃厌氧培养48~96 h分别计数。培养计数后菌落数在30~300的稀释度即为该肠道菌种指标进行计数所需的最适稀释度。选取各指标的最适梯度稀释液100μL均匀涂布于不同的选择培养基上,对菌落数在30~300的平板进行计数,并统计数据。
1.3.4 益生元指数的计算
益生元指数的计算公式如下:
PI=(Bif/Total)-(Bac/Total)+(Lac/Total)-(C los/Total)
上式中,Bif是指取样时发酵液中双歧杆菌的数量和接种时发酵液中双歧杆菌的数量的比值,Bac是指取样时发酵液中拟杆菌的数量和接种时发酵液中拟杆菌的数量的比值,Lac是指取样时发酵液中乳酸杆菌的数量和接种时发酵液中乳酸杆菌的数量的比值,C los是指取样时发酵液中梭状芽孢杆菌的数量和接种时发酵液中梭状芽孢杆菌的数量的比值,Total则是指取样时发酵液中总肠道菌的数量和接种时发酵液中总肠道菌的数量的比值。该公式是以双歧杆菌和(或)乳酸杆菌数量的增长有正面作用,而拟杆菌和(或)梭状芽孢杆菌数量的增长有负面作用为基础的。
1.3.5B/E值的计算
将双歧杆菌的数量除以肠杆菌的数量即得到B/E值,B/E>1表示肠道菌群中双歧杆菌的水平高于肠杆菌科的水平,B/E<1表示肠道菌群中双歧杆菌的水平低于肠杆菌科的水平。
1.3.6 PH值测定
本研究采用pH计Testo205(Testo AG)对样品发酵液进行pH值的测量。将pH计校准后放入厌氧培养箱,在箱内把探头插入装有样品发酵液的10m L EP管中,显示测得的温度和PH值,读数每秒更新2次,AUTO HO LD闪烁直到点亮,读数固定不动后记录读数即为待测样液pH值,按ON/HO LD键重新开始计数,记录三组数据。
1.4 数据分析
所有数据统计分析采用SPSS14.0和O rigin6.0完成,显著性差异(P<0.05)通过Turkey test程序进行。
2 结果与分析
2.1 发酵过程中微生物的变化及分析
本实验通过微生物的定量分析可知,添加有OPO的人体粪便发酵液与空白对照组相比,其总需氧菌数、总厌氧菌数、双歧杆菌、乳酸杆菌、肠球菌、拟杆菌、梭状芽孢杆菌数和肠杆菌在发酵过程中存在显著差异(P<0.05)。
2.1.1 总需氧菌和总厌氧菌的生长变化
由图1不难看出,A组中总需氧菌的数量在发酵第0 h时几乎与B组数量一致,分别是7.77 logCFU/m L和7.80 logCFU/m L,在发酵0~8 h A组与B组菌数一起增加,发酵第8 h时获得各自生长数量最大值,分别为9.12 logCFU/m L和8.60 logCFU/m L,发酵第8 h后A组、B组菌数又一起下降,到发酵第24 h时菌落生长数分别为8.88 logCFU/m L和8.00 logCFU/m L,在整个发酵过程中,A组中总需氧菌的数量一直显著高于(p<0.05)B组。由图2可看出,A组与B组在发酵第0 h的总厌氧菌生长数量分别为7.81 logCFU/m L和7.62 logCFU/m L,B组中总厌氧菌的数量在0~8h缓慢增加,在发酵到12 h时急速下降,A组总厌氧菌的生长数量在0 h~4 h显著增加(p<0.05),虽其菌数在发酵第8 h减少为8.56 logCFU/m L,但在整个发酵过程中A组的菌群数一直显著高于(p<0.05)B组。因此,与空白对照组比较,OPO结构脂对总需氧菌和总厌氧菌都有增值作用,可明显促进总需氧菌和总厌氧菌的生长繁殖速度。
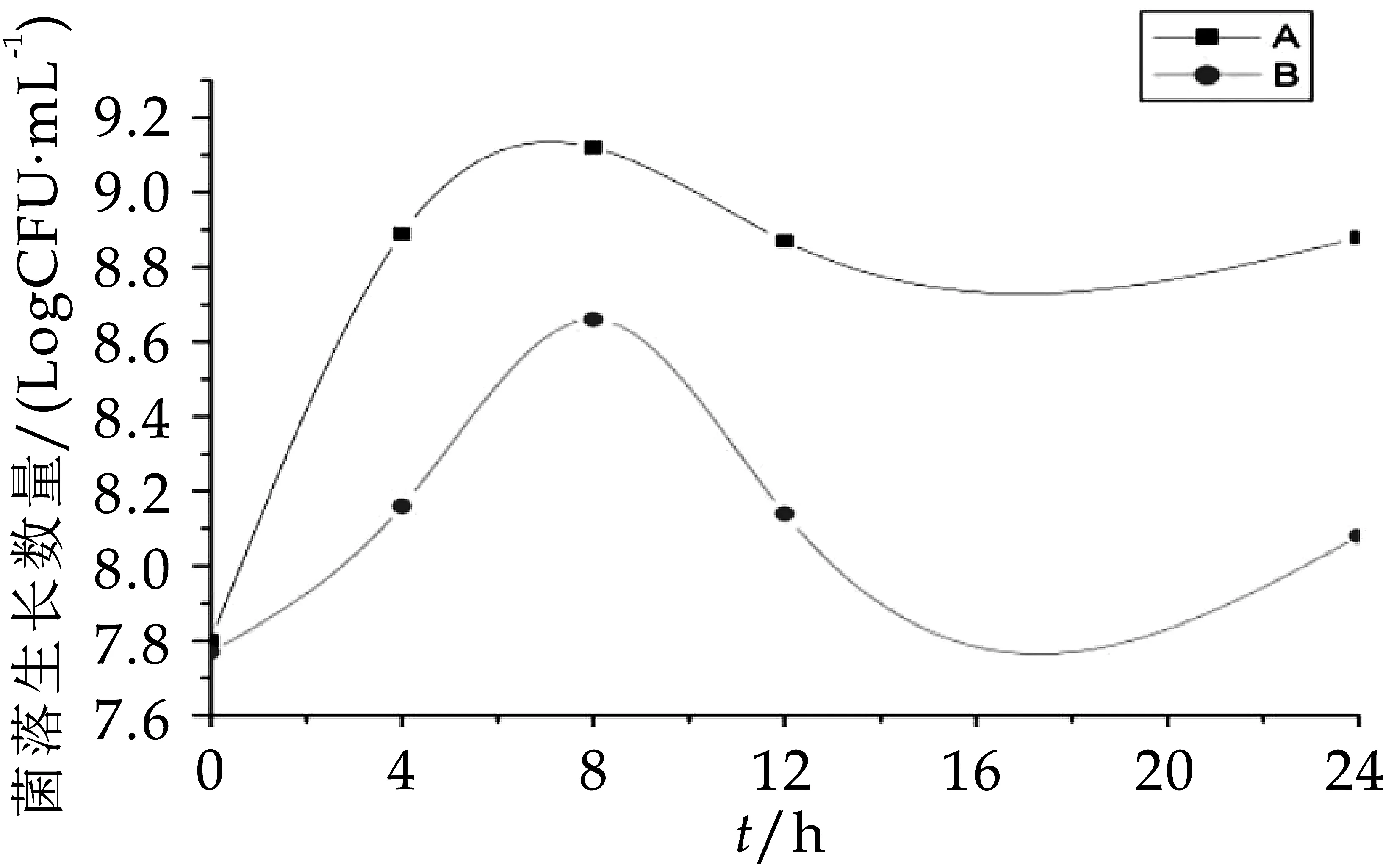
图1 发酵中总需氧菌的生长变化

图2 发酵中总厌氧菌的生长变化
2.1.2 双歧杆菌、乳酸杆菌、肠球菌和拟杆菌的生长变化
从图3-图6可以看出,A组发酵中双歧杆菌、乳酸杆菌、肠球菌和拟杆菌的生长数量均显著高于(p<0.05)B组发酵中的生长数量。A组和B组中的双歧杆菌在发酵第oh的数量差不多,分别为6.86 logCFU/m L和6.84 logCFU/m L,0~8 h A组、B组双歧杆菌数一起增加,到发酵第8h时达到整个发酵过程的菌数最大值,分别为8.61 logCFU/m L和7.80 logCFU/m L,第8 h后双歧杆菌数又一起减少。A组和B组中乳酸杆菌的数量变化趋势十分相似(增加→减少→增加→减少)。A组中肠球菌数量0~8 h显著增加,12 h平缓下降后回升,B组中肠球菌数量不仅数量低于A组,且增长趋势起起伏伏,极不稳定,在发酵第8 h菌数骤减,随后又回升,在16~24 h显著减少。A组和B组的拟杆菌数量变化趋势在0~12h也十分相似,但在12 h后
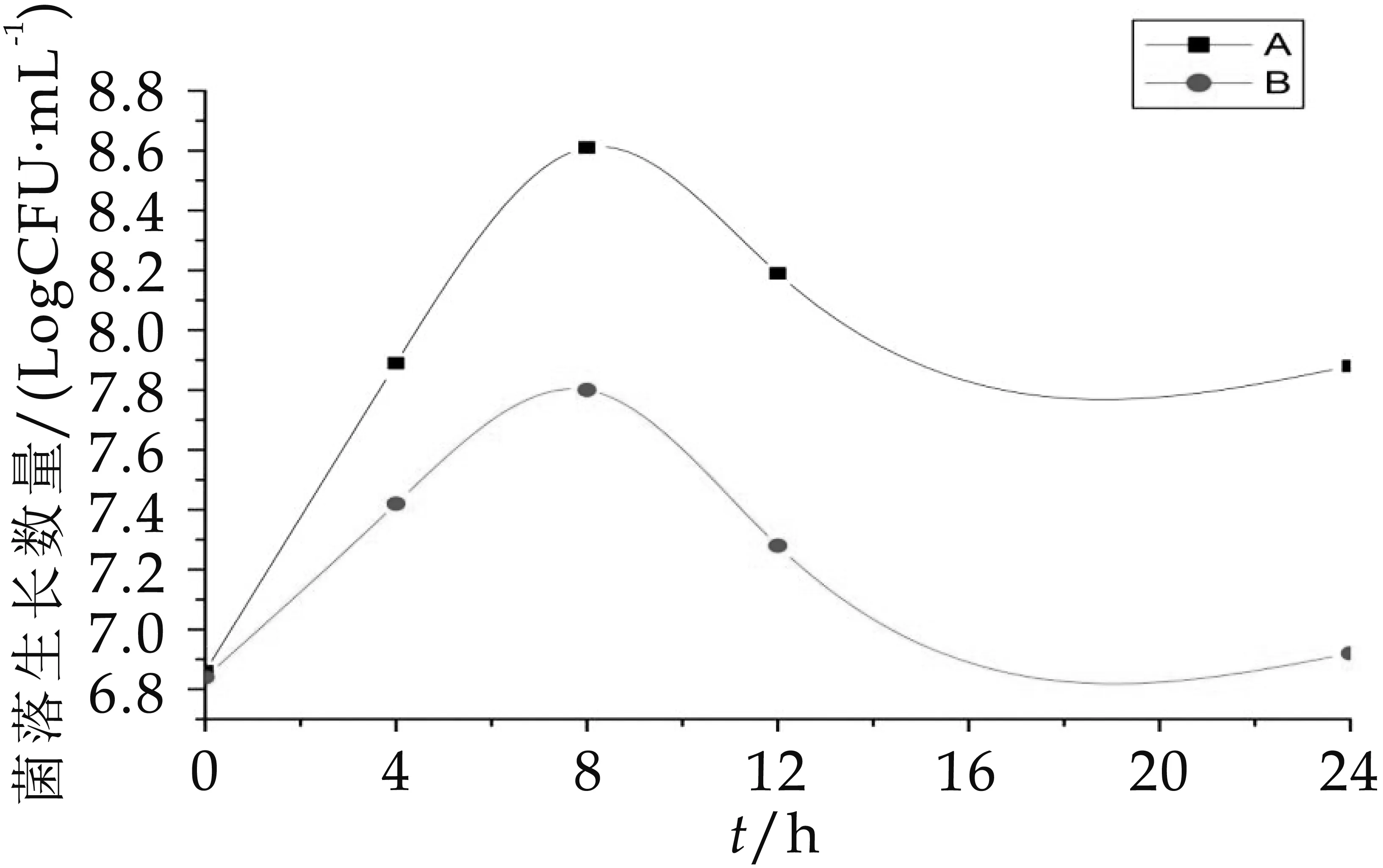
图3 发酵中双歧杆菌的生长变化

图4 发酵中乳酸杆菌的生长变化

图5 发酵中肠球菌的生长变化
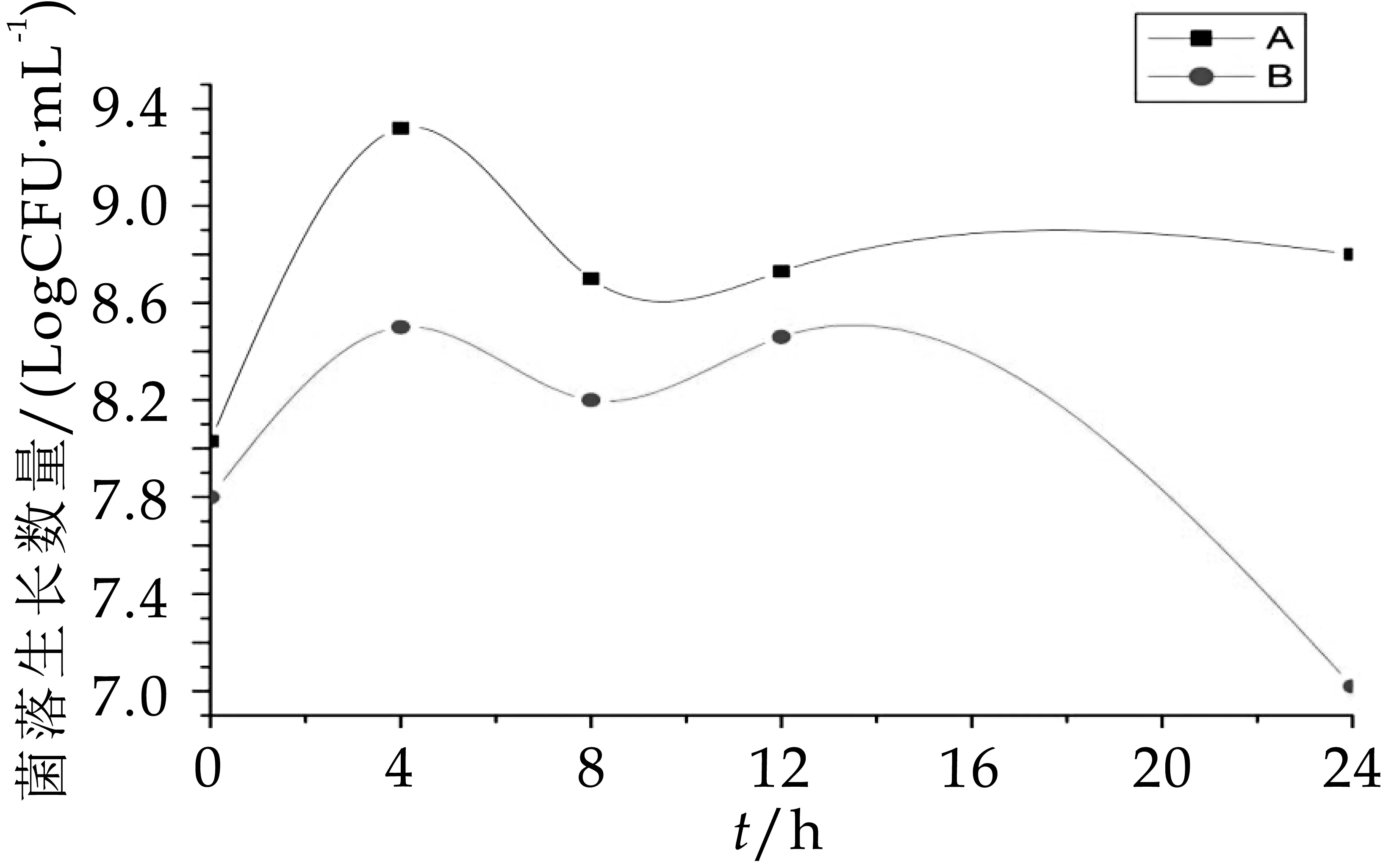
图6 发酵中拟杆菌的生长变化
A组的菌落生长量逐渐趋于平稳,而B组的菌落数却骤降。因此不难看出,OPO结构脂的加入能增加双歧杆菌、乳酸杆菌、肠球菌和拟杆菌的生长数量,且效果显著。
2.1.3 梭状芽孢杆菌和肠杆菌的生长变化
由图7可以看出,B组中梭状芽孢杆菌的数量显著高于(P<0.05)A组,发酵第4 h后A组和B组的菌落生长数分别为7.92 logcfu/m L和8.63 logcfu/m L,4 h后B组的菌落数维持在8.50 logcfu/m L,而A组中梭状芽孢杆菌8 h之后生长数量骤降,到发酵第24 h为7.25 logcfu/m L。由图8可知,A组B组中肠杆菌在发酵0、4、8 h这三个时间段几乎没有生长,其原因可能是人体内肠杆菌的数量本就十分稀少,在发酵第8 h后才有肠杆菌的生长,且B组中的生长数量显著高于A组。因此,从图7~图8可以得知,OPO结构脂能有效抑制梭状芽孢杆菌和肠杆菌的生长数量。

图7 发酵中梭状芽孢杆菌的生长变化
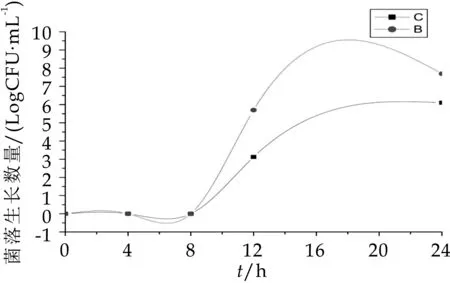
图8 发酵中肠杆菌的生长变化
2.2 发酵中益生元指数计算结果
将肠道菌群的计数值代入PI计算公式后,得出添加了OPO结构脂的A组与未添加OPO结构脂的B组在发酵4、8、12和24 h后的PI值见表1。
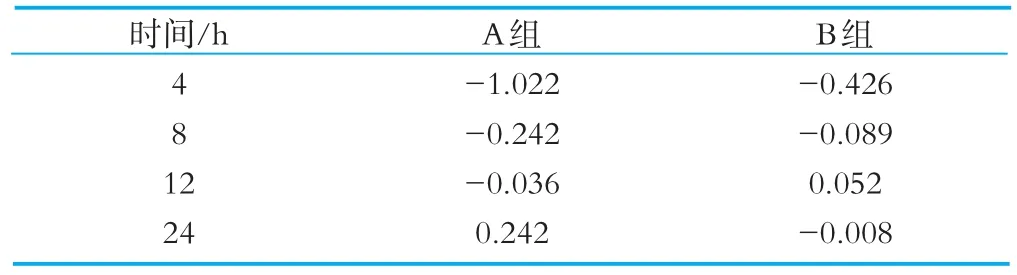
表1 发酵过程中4、8、12、24的益生元指数
由表1的PI计算结果可以看到,A组的PI值在培养到4 h时是负值-0.1022,随着培养时间的增加,从4~24 h逐渐增大,由负值逐渐上升为正值,到培养至24 h时PI值变为正数,为0.242。B组的PI值在培养到4 h时为负值,之后PI值增大,在12 h时PI值转变为正数,为0.052,但培养至24 h时PI值却又变为负数。这些数值的变化说明OPO结构脂具有益生作用,并且其益生作用稳定。
2.3 发酵中B/E值的计算结果
将肠道菌群的计数值带入计算结果如表2所示:
由图8可以得知在0、4、8 h这三个时间段并未检出肠杆菌,而双歧杆菌的生长数量却很大,因此A组和B组在这三个时间段的B/E值都远大于1。由表2的数据可以很明显的看出本次体外发酵实验中加了OPO结构脂的A组在12、24 h这两个时间段的B/E值均大于1,且数值远大于B组。结果表明OPO结构脂能有效抑制肠杆菌等正常菌转变为有害因子、形成有害菌的可能性,并加速双歧杆菌等有益菌的生长繁殖,维持人体肠道平衡,保护人体健康。
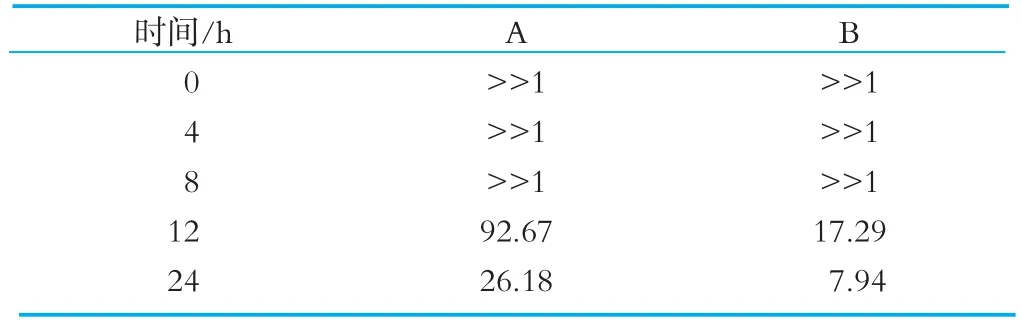
表2 发酵液在0、4、8、12、24h的B/E值
2.4 体外发酵过程中pH值的变化
OPO体外发酵样品pH值测定结果如表3所示。

表3 样品体外发酵pH值
由表3可知,A组和B组的pH值都随着发酵时间的增长而降低,由中性向酸性靠近。A组和B组在0h时的pH值几乎一致,相差不大,但到24 h时A组的pH值降为4.55,而B组的pH值为7.05,可以很明显得出,在pH值降低的过程中,A组的降低速度比B组快,能够更快地向酸性环境靠近,并且A组pH值下降差异显著(p<0.05),下降到8 h以后趋于平缓,维持在4.5左右,B组下降速度缓慢,差异不明显。因此,OPO结构脂能促进肠道菌群发酵,使其产生酸性物质,从而使人体肠道内环境向酸性靠近。
3 讨论
人体内的肠道菌群种类繁多,数量庞大,其生长环境中存在着各种影响其生长发育的因素,例如内在因素:人体的基因组[16]、新陈代谢[17]、性别[18]、种族[18]、免疫系统[19]等;外在因素:地域[20]、饮食习惯[21]等。肠道菌群中有些是厌氧菌,如拟杆菌、梭状芽孢杆菌和双歧杆菌等,有些是兼性厌氧菌如乳酸杆菌、肠球菌和肠杆菌等。
肠道菌群对维持人体内环境,加速新陈代谢具有十分重要的主导地位,不同的肠道菌群对生长坏境的要求也不尽相同。例如双歧杆菌,作为一种人体内普遍存在的、与人体健康密切相关的肠道有益菌,它不仅可以产生一种有机酸来抑制非有益菌的生长,还能够维持人体内肠道菌群的平衡,促进菌群新成代谢,帮助肠道蠕动。但随着人类年龄的增长,肠道内环境的改变,双歧杆菌的数量随之减少,已有研究者证明,一个成年人肠道内的双歧杆菌数量远远少于其婴幼儿时期的肠道内双歧杆菌数量,仅仅只占其体内细菌总数的3%左右[22]。乳酸杆菌也是一种人体内普遍存在的厌氧菌,M onique Haarman等[23]在其2006年的研究报告中指出,乳酸杆菌是一种帮助人体抑制疫病的有益菌,它能有效抑制腹泻和过敏性疾病等的发生。为探寻OPO结构脂对人体内不同肠道微生物生长发育的影响程度,本研究通过预实验获得八种肠道微生物最适生长稀释度后,对其进行增殖培养和数据分析。对照实验表明,整个发酵过程中总需氧菌、总厌氧菌、双歧杆菌、乳酸杆菌、肠球菌、拟杆菌这6个指标,在加了OPO结构脂后发酵液稀释倍数都高于空白对照组。由此可知,OPO结构脂可以增加大学生体内肠道菌群的生长数量,具有维持肠道菌群平衡、增强人体免疫力、预防疾病的功能。
Palfram an等人[24]在2003年首次提出益生元指数的概念,考虑到要维持不同肠道菌群间的平衡和取样接种时带来的误差等影响因素,他们认为可以通过计算益生元指数来衡量益生元的作用,且整个过程可以在体外进行。B/E值是指肠道内双歧杆菌和肠杆菌数量的比值,由双歧杆菌的数量和相对应的肠杆菌科的数值相比得到。最早是由荷兰学者Van DerW aaij等人[25]提出。双歧杆菌是肠道菌群有益菌的典型代表,而肠杆菌则被认为是肠道中正常菌群向不利于机体健康的方向转变的重要因子,也可以作为潜在有害菌的代表,两者的比值可以从正反对比评价肠道菌群结构的状况。本研究发现,OPO结构脂对大学生肠道菌群具有稳定的益生作用。与此同时,OPO结构脂还能够有效抑制肠杆菌等正常菌转变为有害因子,降低有害菌形成的可能性,并且加速双歧杆菌等有益菌的生长繁殖,维护大学生机体健康。
有研究表明,拟杆菌是人体肠道内占绝对优势的一种厌氧菌,无论是其在人体肠道内的种类还是数量都非常庞大。曾红根等[26]在2011年以动物为实验对象,模拟出了拟杆菌微生态系统,通过该系统对动物进行饲养,研究结果表明,在拟杆菌微生态系统中生长发育的动物肠道内PH值比在自然环境中生长发育的动物更低,呈现出一种酸性状态,且在这种拟杆菌微生态系统中生长的动物能产生更多免疫抗体,更具有抵抗疾病的免疫力。乳酸杆菌可以作用于人体内的二氧化碳,使之发酵形成乳酸,从而降低肠道内的pH值,形成一个酸性环境,不仅可以帮助人体消化吸收,还能够阻止肠道内某些有害菌的生长繁殖,降低其生存力,以免对人体新陈代谢造成阻碍,进而改善人体肠道内环境,促进肠道蠕动,降低疾病的发生率[27],并减少人体肠道中潜在致癌物的形成数量[28-29]。通过计算PI值与B/E值,本研究更能确定OPO结构脂能增加
肠道内酸性物质的产生,并具有促进肠道有益菌群生长、增强肠道的消化吸收等功能,保护人体健康。
4 结论
上述结果表明,以OPO结构脂作为样品,通过体外发酵,可以得知OPO对大学生体内肠道菌群有以下作用:有效抑制正常菌转变为有害因子,降低有害菌形成的可能性,加速有益菌的生长速度,帮助肠道蠕动,促进机体新陈代谢。OPO结构脂还能协助益生菌产生酸性物质,如乙酸等,降低肠道pH,从而抑制腐败菌的增殖,增强大学生对外界干扰因素的抵抗力,保护大学生机体健康。本研究着重于比较OPO体外发酵过程中微生物的变化情况,并在此基础上分析OPO结构脂与肠道菌群的相关性和环境对其体外发酵的影响,为以后OPO结构脂的研究提供新思路。
[1]李卫林.猪油的应用前景[J].粮油食品科技,2007,15(1):52-53.
[2]符剑刚,钟宏.猪油的开发利用[J].粮油食品科技,2003,11(4):28-29.
[3]章朝晖,龙罡,黄静.猪油资源的深加工[J].中国油脂,2001,26(4):40-41.
[4]武庆斌,郑跃杰,黄永坤.儿童肠道菌群——基础与临床[M].科学出版社,2012.06:17-18.
[5]O'HARA A M,SHANAHAN F.The gut flora as a forgotten organ[J].EMBO reports,2006,7(7):688-693.
[6]KEDIA G,VAZQUEZ JA,CHARALAMPOPOULOSD,et al.In vitro fermentation of oat bran obtained bydebranning w ith a m ixed culture of human fecal bacteria[J].Curr M icrobiol,2009,58(4):338-342.
[7]BLAUT M,CLAVEL T.M etabolic diversity of the intestinalm icrobiota:implications for health and disease[J].The Journal of nutrition,2007,137(3):751S-755S.
[8]冯晓玲.我国青少年身体素质下降的成因分析与对策研究[J].2016:11-12
[9]ALLISON C,MCFARLAN C,MACFARLANE.Studies on m ixed populations of human intestinal bacteria grown in single-stage and multistage continuous culture systems.[J].Applied and environmental m icrobiology,1989,55(3):672-678.
[10]W ILKINST D,CHALGREN S.Medium for use in antibiotic susceptibility testing of anaerobic bacteria[J].Antim icrobial agents and chemotherapy,1976,10(6):926-928.
[11]BEERENSH.An elective and selective isolationmedium for bifidobacterium spp.[J].Letters in Applied M icrobiology,1990,11(3):155-157.
[12]ROGOSAM,M ITCHELL JA,W ISEMAN R F.A selectivemedium for the isolation and enumeration of oral and fecal lactobacilli[J].Journalof Bacteriology,1951,62(1):132-133.
[13]TAYLOR EW,BURMAN N P.The application ofmembrane filtration techniques to the bacteriologicalexam ination ofwater[J].Journalof Applied M icrobiology,1964,27(2):294-303.
[14]DEVOSN,MEVISSEN-VERHAGE E,VAN AMERONGEN W H,et al.A new selectivemedium for the culture of Clostridia from human faeces[J].Eur JClin M icrobiol,1982,1(5):267-271.
[15]MACFARLANE G T,HAY S,GIBSON G R.Influence ofmucin on glycosidase,protease and arylam idase activitiesof human gut bacteria grown in a 3-stage continuousculture system[J].Journalof Applied M icrobiology,1989,66(5):407-417.
[16]ZOETENDAL EG,AKKERMANSA D,AKKERMANSW M,et al.The host genotype affects thebacterial community in the human gastrointestinal tract[J].M icrobialEcolHealthDis,2001’13:129-134.[17]BARTOSCH S,FITE A,MACFARLANEG T,et al.Characterization of bacterial communitiesin feces from healthy elderly volunteers and hospital ized elderly patients by usingreal-time PCR and effects of antibiotic treatment on the fecalm icrobiota[J].ApplEnviron M icrobiol,2004,70(6):3575-3581.
[18]MUELLER S,SAUN IER K,HAN ISCH C,et al.Differences in fecalmicrobiota in differentEuropean study populations in relation to age,gender,and country:a cross-sectionalstudy[J].Appl Environ M icrobiol,2006,72(2):1027-1033.
[19]PETERSON D A.IgA response to symbiotic bacteria as amediator ofguthomeostasis[J].CellHostM icrobe,2007,2(5):328-339.
[20]MUELLER S,SAUN IER K,HAN ISCH C,et al.Differences in fecalmicrobiota in differentEuropean study populations in relation to age,gender,and country:a cross-sectionalstudy[J].Appl Environ M icrobiol,2006,72(2):1027-1033.
[21]LEY R E,HAMADY M,LOZUPONEC,etal.Evolution ofmammals and their gut m icrobes[J].Science,2008,320(5883):1647-1651.
[22]SGHIR A,GRAMET G,SUAU A,et al.Quantification of bacterial groupsw ithin humanfecal flora by oligonucleotide probe hybr idizatIon[J].Appl Environ M icrobiol,2000,66(5):2263-2266.
[23]MONIQUE H,JAN K.Quantitative Real-Time PCR Analysis of Fecal Lactobacillus Speciesin Infants Receiving a Prebiot ic Infant Formula[J].ApplEnviron M icrobiol,2006,72(4):2359-2365.
[24]PALFRAMAN R,GIBSON G R,RASTALL R A.Development of a quantitative tool for the comparison of the prebiotic effect of dietary oligosaccharides[J].Lett ApplM icrobiol,2003,37(4):281-284.
[25]VAN DER WAAIJD,BERGHUIS-DE V J,LEKKERKERK L.Colonization resistance of the digestive tract in conventionaland antibiotic-treatedmice[J].JHyg(Lond),1971,69(3):405-411.
[26]曽红根,付金衡,邓红,等.拟杆茵微生态制剂对仔猪免疫能力的影响[J].中国微生态学杂志,2011,23(11):989-990.
[27]JELENA S,SEPP E,KOLK H,et al.Diversity and metabolic impact of intestinalLactobaci1lus species in healthy adults and the elderly[J].Br JNutr,2011,105:1235-1244.
[28]M ITSUOKA T.Bifidobacteria and their role in human health[J].Journal of Industrial M icrobiology&Biotechnology,1990,6(4):263-267.
[29]FERMANDES C F,SHAHAN I K M,AMER M A.Therapeutic role of dietary lactobacilli and lactobacillic fermented dairy products[J].FEMSM icrobiology Letters,1987,46(3):343-356.
