Hsa-miR-219-5p靶基因的预测及其生物信息学分析
2018-06-13刘润燕张运峰
屈 燕,刘润燕,黄 帅,高 晶,张运峰,胡 芬
(1. 华北理工大学 生命科学学院,河北 唐山 063000;2. 唐山师范学院 生命科学系,河北 唐山 063000)
miRNA在胚胎的发育、细胞的增殖和分化以及肿瘤的发生等许多生物学功能上都发挥着重要的作用[1-3]。目前,许多研究结果均已证实在肿瘤组织中miRNA的异常表达会导致其调节的靶基因表达异常,进而使miRNA有促进或抑制肿瘤发生的功能[3]。由于 miRNA是通过其靶基因来发挥功能的,所以找到miRNA调控的靶基因是对其生物学功能研究至关重要的一步。
hsa-miR-219-5p为内含子型miRNA,最新研究证实,miR-219参与了细胞增殖、分化和凋亡调控,并在肝癌[4-6]、脑胶质细胞瘤[7]、胃癌[8]和卵巢癌[9]等多种癌症中发挥着重要的生理功能。miR-219通过靶向 SALL4在脑胶质细胞瘤生长和转移中起抑制作用[7]。此外,miR-219-5p通过下调 Twist/Wnt/β-连环蛋白信号通路抑制卵巢癌细胞增殖、迁移和侵袭[9]。DNA异常甲基化是 microRNA异常表达的机制之一,在肝癌中miR-219上游1094bp区域CpG岛存在异常甲基化,miR-219在肝癌细胞株的表达下调,证实miR-219是潜在的抑癌基因[4]。还有研究发现,长时间上夜班的女性,miR-219启动子高甲基化,从而降低了miR-219的表达,导致免疫介导的抗肿瘤活性下调和乳腺癌风险增加[10]。本研究利用生物信息学的工具进行多重比较,寻找新的hsa-miR-219-5p调控靶基因,进行功能分析。
1 方法
1.1 miR-219-5p的成熟序列的获得及其物种进化分析
从 miRBase数据库①获得 10个物种的miR-219-5p的成熟序列。采用MEGA5.0软件分析miR-219-5p的物种间进化的保守性。
1.2 has-miR-219-5p的靶基因预测
采用TargetScan7.1和miRDB两大miRNA的靶基因预测中的生物软件对 hsa-miR-219-5p的靶基因进行了预测,获取两大软件预测到的靶基因的交集,再与 miRTarbase中已经证实的hsa-miR-219-5p的靶基因取并集,最终形成靶基因集合用于后续生物信息软件分析。
1.3 靶基因集合的Gene Ontology(GO)富集分析
应用GO生物数据库对上述has-miR-219-5p的靶基因集合进行了生物学过程以及分子功能注释和富集分析,经显著性分析,p<0.01为显著性差异,得到具有统计学意义的基因注释和富集分析结果。
1.4 靶基因集合的 Kyoto Encyclopedia of Genes and Genomes(KEGG)富集分析
利用 KEGG数据库对 hsa-miR-219-5p的靶基因集合进行了信号转导通路富集分析,通过Fisher Exact Test统计分析具有统计差异的p值,p<0.01表示显著性阈值,代表靶基因集合相对于背景具有统计学意义的信号传导通路。
2 结果
2.1 miR-219-5p成熟序列进化分析
从 miRBase数据库中获取了多个物种的miR-219-5p成熟序列,其中人的hsa-miR-219-5p为内含子型miRNA,hsa-miR-219的前体序列位于人的六号染色体33207835-33207944[+],成熟序列是 5’-UGAUUGUCCAAACGCAAUUCU-3’。采用MEGA5.0对人、猕猴、小鼠、安大略鲑、大鼠、牛、大猩猩、非洲瓜蟾、马、斑胸草雀10个物种的 miR-219-5p的成熟序列进行了比对分析,结果显示miR-219-5p在牛中是22个碱基,在安大略鲑和斑胸草雀中是 23个碱基,其余的都是 21个碱基,且碱基序列完全一致,如图 1所示。该结果表明 miR-219-5p的成熟序列在脊椎动物进化过程中具有高度的保守性。
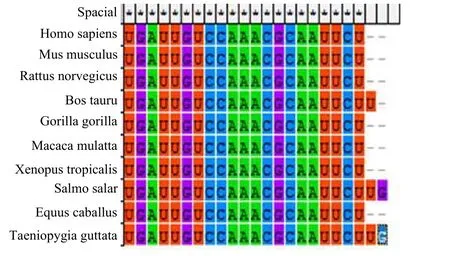
图1 10种不同物种中miR-219-5p成熟序列的比对分析
2.2 has-miR-219-5p的靶基因预测
TargetScan7.1预测hsa-miR-219-5p有445个靶基因,而 miRDB预测其靶基因数量为 174,对上述两种软件的预测结果取交集,最后共获得123个靶基因。另外,miTarbase数据库中显示的经双荧光素酶报告基因检测和/或 Northern blotting试验证实的 hsa-miR-219a靶基因共有 6个(CAMK2G、GPC3、MT1F、PSMD6、EGFR、TRIB3),其中 3个和预测的一致,占所有预测靶基因集合的 2.4%。合计预测靶基因和已经实验验证的靶基因,共计获得126个靶基因的数据集合用于后续的基因注释和信号通路富集分析。
GXYLT1, EYA1, FAM91A1, ELMOD2, GBA2, LEF1,SLC16A7, SDK1, CD164, INPP5J, DAZAP1, UBASH3B,PPARGC1A, ANKRD44, THRB, CHD7, TRHDE, RECK, PIGR,PDGFRA, ABCB10, AFF4, MEF2D, DCAF10, KCNA4, DDAH1,SOX6, CAPS, RORB, PIP5K1C, FBXO30, CBX2, RASSF3,PCDH17, PRDM16, ETV5, SMC4, ATG14, SEMA4G, FOXJ3,CGNL1, TSC22D2, FZD4, CRLF3, PHF19, TMEM98, FMNL2,SNTB2, CLOCK, ZBTB18, FAM199X, DIAPH3, GBP5,LAPTM4A, FURIN, ISL1, GLTSCR1L, COL9A1, ASH1L,SLC41A1, ZCCHC14, KMT2D, DNAL1, SP4, ELOVL7,CCDC28A, ROR1, NR2C2, SLC35F5, MECOM, SCARB2,SLC39A10, SLC31A1, FBXO3, TGFBR2, KIAA1549, RIMS1,SLC16A6, ADD2, PKNOX1, MIER3, RNF6, SNRK, MMS19,NDRG4, CC2D1A, RPRD2, SOX14, TMX4, UBE3A, ABHD13,THSD7B, PKDCC, TPCN1, DOK6, AFAP1L2, KBTBD8, HAS3,CBFA2T3, ZNF609, CACUL1, ACOX1, SLC30A4, LGALSL,PIGG, DNAJC6, AKAP13, LPP, TRIB3, EGFR, PSMD6, GPC3,UBE2Z, KLF16, KCNH8, NEK6, T, MT1F, STRBP, CAMK2G,UBR1, PDE4D, MFNG, INSIG1, SYT5, RBMS3
2.3 hsa-miR-219-5p靶基因的GO富集分析
为进一步分析hsa-miR-219-5p的生物功能,应用GO数据库对上述靶基因集合进行了GO注释、生物学过程和分子功能分类富集分析,结果126个靶基因中,共获得122个基因的GO注释描述和生物功能注释信息。通过分析发现,hsa-miR-219-5p靶基因的功能主要集中基因的转录与表达、细胞内合成与代谢等,如表1所示。
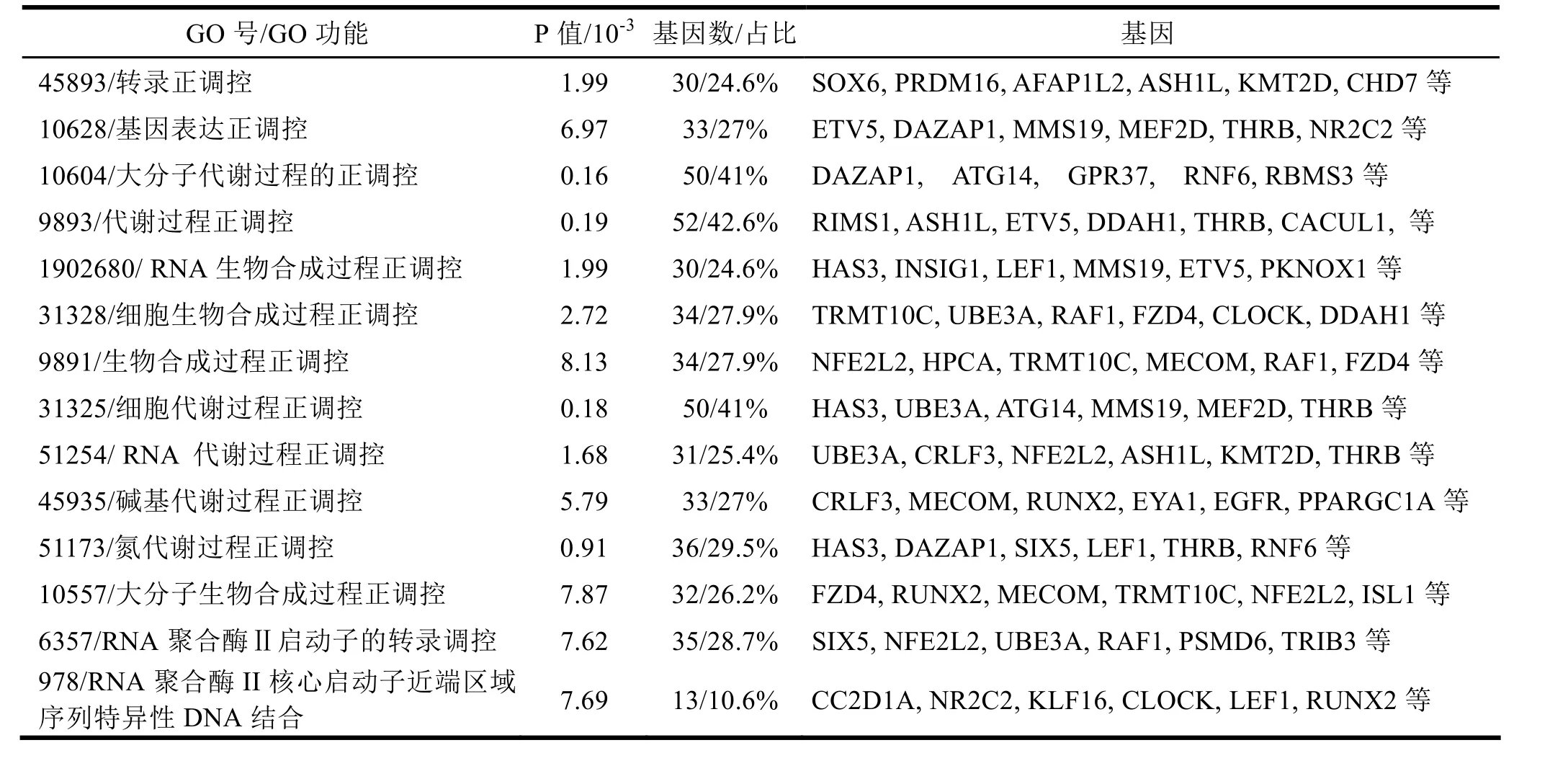
表1 hsa-miR-219-5p靶基因集合的GO生物学过程和分子功能分类结果
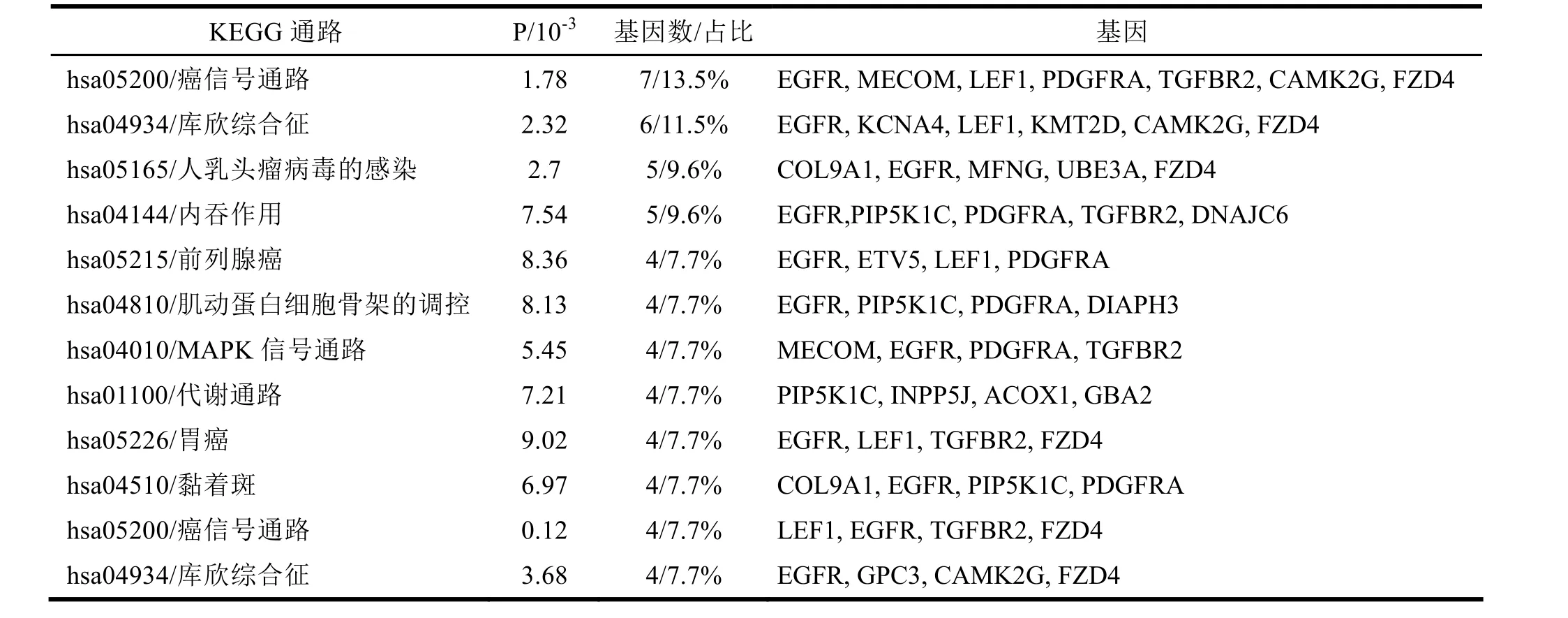
表2 hsa-miR-219-5p靶基因集合的KEGG通路富集分析结果
2.4 hsa-miR-219-5p靶基因集合的 KEGG富集分析
miRNAs的众多靶基因通常协同参加同一细胞信号通路,本研究采用KEGG经典通路数据库对 hsa-miR-219-5p的靶基因集合进行了信号通路富集分析。hsa-miR-219-5p的靶基因显著富集于癌信号通路、肌动蛋白细胞骨架调控、MAPK等相关信号通路以及前列腺癌、胃癌、肝癌等疾病信号通路,如表2所示。
3 讨论
miR-219-5p在多个物种中的成熟序列高度一致,进化过程中相对保守,表明该基因参与了重要的生命基本活动,GO富集分析也表明miR-219-5p的靶基因参与基因转录翻译多个过程。
采用生物信息学方法进行miRNA靶基因预测方法很多,但应用最为广泛的是 miRDB、TargetScan7.1和PicTar三大预测软件[11-12]。这三种软件算法不同、各有优缺点,采用TargetScan7.1和miRDB两种软件联合应用的方法对hsa-miR-219-5p的靶基因进行预测,并取两种软件均预测的交集,联合 miTarbase数据库中已经实验验证的靶基因,共同作为hsa-miR-219-5p的靶基因集合,用于后续分析,大大提高了预测的准确性、减少了假阳性。
通过对hsa-miR-219-5p的靶基因进行KEGG信号通路富集分析发现,hsa-miR-219-5p的靶基因参与了 MAPK等信号通路,并且显著地富集于肝癌、胃癌、前列腺癌等多种疾病,因此has-miR-219-5p的表达异常可能参与了一些疾病的发生。这一结果与已有文献报道[4-8,13]相一致。
[注释]
① 从 http://www.mirbase.org/中下载 Homo sapiens(人)、Macaca mulatta(猕猴)Mus musculus(小鼠)、Salmo salar(安大略鲑)、Rattus norvegicus(大鼠)、Bos taurus(牛)、Gorilla gorilla(大猩猩)、Xenopus tropicalis(非洲瓜蟾)、Equus caballus(马)、Taeniopygia guttata(斑胸草雀)共10个物种的成熟序列。
[1] Bartel D P. MicroRNAs: genomics, biogenesis, mechanism, and function[J]. Cell, 2004, 116(2): 281- 297.
[2] Wightman B, Ha I, Ruvkun G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans[J].Cell, 1993, 75(5): 855-862.
[3] Iorio MV, Croce CM. MicroRNAs in cancer: small molecules with a huge impact[J].J Clin Oncol, 2009,27(34): 5848-5856.
[4] Huang N, Lin J, Jian R, et al. MiR-219-5p inhibits hepatocellular carcinoma cell proliferation by targeting glypican-3[J]. Febs Letters, 2012, 586(6): 884-891.
[5] 黄娜.MiR-219-5p在肝细胞癌患者中的表达与其对增殖的作用和机制研究[D].广州:南方医科大学,2012:1-2.
[6] 朱理辉,罗勇,廖文秋,等.MicroRNA-219-5p 靶向 E-钙黏蛋白调控上皮间质转化抑制肝癌细胞侵袭转移[J].中国现代医学杂志,2016,26(18):22-29.
[7] Jiang B, Li M, Ji F, et al. MicroRNA-219 exerts a tumor suppressive role in glioma via targeting Sal-like protein 4[J]. Experimental & Therapeutic Medicine, 2017, 14(6):6213-6221.
[8] Li C, Dong J, Han Z, et al. MicroRNA-219-5p represses the proliferation, migration, and invasion of gastric cancer cells by targeting the LRH-1/Wnt/β-catenin signaling pathway[J]. Oncology Research, 2016, 25(4):617-627.
[9] Wei C, Zhang X , He S , et al. MicroRNA-219-5p inhibits the proliferation, migration, and invasion of epithelial ovarian cancer cells by targeting the Twist/Wnt/beta- catenin signaling pathway[J]. Gene,2017, 637: 25-32.
[10] Shi F, Chen X, Fu A, et al. Aberrant DNA methylation of miR-219 promoter in long-term night shiftworkers[J]. Environmental & Molecular Muta- genesis, 2013,54(6): 406-413.
[11] 高晶,张运峰,胡芬.Has-miR-28的靶基因预测及其生物信息学分析[J].唐山师范学院学报,2017,39(5):61-64.
[12] Lewis B, Shih I, Jones Rhoades M, et al. Prediction of mammalian micro-RNA targets[J]. Cell, 2003, 115(7):787-798.
[13] 田立立.miR-219-5p靶向e-钙粘蛋白促进人前列腺癌细胞迁移侵袭[D].湛江:广东医学院,2012:1-2.
