金盏菊对重金属Zn的生理响应及累积性研究
2018-05-25潘春香王佳豪
庞 琢,潘春香,王佳豪
(1.呼和浩特市土壤肥料工作站,内蒙古 呼和浩特 010020;2.韶关学院 英东农业科学与工程学院,广东 韶关512005)
Zn是植物、动物和人体生命活动不可或缺的微量元素,但过量摄入Zn会刺激胃肠道,损伤免疫功能,引起人体健康问题[1].植物修复重金属污染土壤技术因其治理成本低、污染少,操作简单便捷和不破坏土壤原本组成和结构等优点而成为当前研究和开发热点.已报道的重金属Zn超累积植物有18种,如十字花科遏蓝菜属的天蓝遏蓝菜(Thlaspi carulescens L.)对Zn的累积量达到51 600 mg/kg[2];野菊花、荨麻、白苏、银柳、香根草等对Zn富集系数较高,根系运转系数均大于1,耐受性强,且生物量大、适应性强[3].白菜、垂盆草及萝卜[4]、黑麦草[5]、木麻黄、小叶桉[6]、蜈蚣草[7]、东南景天(Sedum alfredii Hance)[8]等对Zn有较强的富集性.花卉植物作为修复重金属污染土壤的材料,不仅不会进入食物链而危害人体健康,还可美化环境、愉悦心情,近年来备受关注.金盏花(Calendula officinalis L.)由于生物量大、生长迅速、管理简单、适应性和观赏性强,在我国南北方广泛种植.如果该植物对重金属有较强的耐性,将有利于在全国各地迅速推广.本试验采用盆栽砂培法展开了金盏菊对重金属Zn的生理响应及累积性的研究,旨在明确金盏菊对重金属Zn的耐受性,同时也为植物修复重金属污染土壤研究提供参考.
1 材料和方法
1.1 试验时间、地点
试验于2015年9月至2016年1月在韶关学院英东楼C座天台的大棚内进行.为了排除土壤中固有元素,采用砂培盆栽进行试验.
1.2 试验材料
试验以金盏菊为供试材料.
1.3 试验方法
1.3.1 试验设计
将不同Zn浓度的溶液定期定量加入营养液中,与营养液一同加到种植金盏菊的盆中,最后分析植物体内重金属的含量,进而分析其耐受性.Zn浓度确定参照GB 15618-2008《土壤环境质量标准》和农业用地 pH 值划分,200 mg/kg(pH5.5~6.5)为依据分别设 150 mg/kg(1 号)、200 mg/kg(2 号)、250 mg/kg(3 号)、300 mg/kg(4 号)4 个浓度梯度,对应换算的 ZnSO4·7H2O 浓度分别为 791.69 mg/kg,967.62 mg/kg,1 143.55 mg/kg和 1 319.48 mg/kg;0 mg/kg为对照(CK).1.3.2实验材料的管理及重金属Zn处理
选择外形饱满,大小相近的金盏菊种子进行催芽,待发芽后每盆(上口径180 mm×下口径130 mm×高160 mm)播种8粒种子进行育苗;待植株长至4片真叶时定苗,每盆留5株,3次重复.为了避免营养液和Zn溶液渗漏到地面,将花盆置于底部防渗漏的塑料箱中(长71 cm×宽64 cm×高18 cm),每箱放置6盆.基础营养液采用日本园试通用配方,浓度为1个剂量,每次每盆50 ml,每隔2~3 d灌一次.
2015年12月17日开始进行重金属Zn处理,每盆每次50 ml Zn溶液,每2~3 d处理一次,共处理5次;2016年1月7日开始分析测定各项指标.1.3.3测定项目与方法
(1)形态指标和生物量的测定
每处理随机取样10株,测定其株高、叶片数、黄叶落叶数、最大叶长、最大叶宽、根长、地上部和根系的鲜重和干重等.
(2)生理指标的测定
叶绿素、类胡萝卜素含量测定采用比色法[9],可溶性蛋白质含量采用考马斯亮蓝G-250法[7],可溶性糖含量测定采用蒽酮比色法[10].
(3)重金属 Zn的测定
取植物干样品0.500 g,加入消化管中,用硝酸∶高氯酸=4∶1的比例配制成混酸,取10 ml混酸加入到植物干样品中,将消化管放入消化炉中进行消化,消化成淡黄色或透明的液体至2 ml左右,静止,用滤纸进行过滤后定容到50 ml的容量瓶中,用原子吸收分光光度计测定[11].
(4)统计分析
所得数据采用SPSS 19.0软件包进行方差分析,用Duncan新复极差法进行平均数的显著检验.
2 结果与分析
2.1 不同浓度重金属Zn处理后金盏菊植株生长的变化
由表1可见,1号处理的株高、茎粗、叶片纵径、叶片横径、叶片数及根长均高于CK和其余处理组,其中茎粗达到了极显著水平,根长达到了显著水平,其它指标不显著.

表1 不同浓度重金属Zn处理后金盏菊生长情况
2.2 不同浓度重金属Zn处理后金盏菊生物量的变化
由表2可见,1号处理的金盏菊的根系和地上部的鲜重、干重以及干物率均超过了CK和其余处理组,且除CK地上部和4号根系的干物率外,与其它处理间差异显著.其中地上部鲜重为1号、CK与其余处理差异极显著,2号、3号与CK、4号差异极显著,3号与2号、4号之间差异极显著;地上部干重为1号与CK、其余处理之间,3号与2号、4号之间差异极显著,3号与CK之间差异显著;地上部干物率为1号与其余各处理差异显著,CK与2号差异极显著,与3号、4号差异显著;根系鲜重为1号与其余处理间显著,其余各处理间差异不显著;根系干重为1号与其它处理差异显著,其余各处理间差异不显著;根系干物率为1号与其它处理差异显著,其它各处理间差异不显著.

表2 不同浓度重金属Zn处理后金盏菊生物量的变化
2.3 在不同浓度重金属Zn处理下金盏菊生理指标的变化
由表3可见,1号金盏菊的叶绿素、地上部和根系的可溶性糖、可溶性蛋白质含量均居第一,且基本呈随着Zn浓度的增大而呈提高的趋势.叶绿素的1号、4号与CK、3号之间差异显著;地上部可溶性糖为1号与2号、3号和4号处理之间差异极显著;根系可溶性糖为1号与CK、3号、4号处理间差异极显著,其它处理间差异显著;地上部可溶性蛋白质各处理间差异均极显著;根系可溶性蛋白质为1号、CK和3号与2号、4号间差异显著.
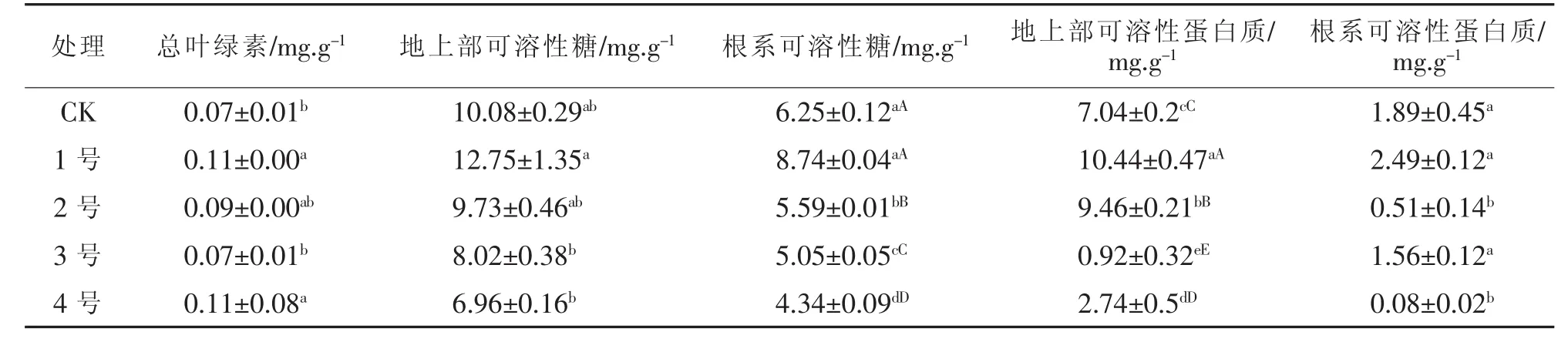
表3 不同浓度重金属Zn处理后金盏菊各生理指标变化
2.4 不同浓度Zn处理后对金盏菊植株体内Zn含量的变化
由表4可见,各处理的金盏菊植株地上部和根系Zn含量均超过了CK,并随Zn浓度的增大呈直线上升趋势,且根系Zn含量明显高于地上部,各处理组与CK之间差异均极显著.其中4号与其余处理间差异极显著,3号、2号与1号处理间差异极显著.Zn运转系数,即地上部Zn含量与根系Zn含量比值(S/R)均低于1,且均低于CK,CK的植株体内仍有Zn积累,是由于营养液中含有Zn元素.

表4 不同浓度重金属Zn处理后金盏菊体内Zn含量变化
3 结论与讨论
3.1 不同浓度重金属Zn处理对金盏菊植株生长的影响
Zn在植物体内的生理作用主要表现在3个方面:参与组成植物体内重要的结构物质成分;是植物生命活动的调节剂参与各种酶的活动,影响植物的代谢;起电化学作用,参与渗透调节、胶体的稳定和电荷中和等[12].因此,适量的Zn能够促进植物的生长发育,但过量植物生长会受阻.本试验结果为150 mg/kg时,金盏菊株高、茎粗、叶片数、地上和根系的鲜重、干重均达到了最大值,之后随着浓度增大,生长明显受到抑制,与丁海东等[13]在番茄上的研究、金新文等[14]在西葫芦上的研究、努扎艾提·艾比布等[15]在香根草上的研究和李翠等[16]在大油芒的研究结果基本一致.但努扎艾提·艾比布等[15]的研究结果显示,当Zn浓度为5 mg/L时促进香根草生物量的累积,20 mg/L浓度时,香根草生物量积累开始下降[15].但努扎艾提·艾比布等[15]的研那么是金盏菊对Zn的耐受性强于香根草?还是因两个试验所用基质不同(本试验为沙培,香根草为水培),还有待进一步考证.
3.2 不同浓度重金属Zn处理对金盏菊生理指标的影响
叶绿素是植物进行光合作用的主要色素,叶绿素的含量与植物的光合速率密切相关,常用叶绿素含量的高低来表征植物在逆境下受伤害的程度[17].Zn能影响碳水化合物的代谢,参与叶绿素的合成,可以促进光合作用[13].缺Zn导致水稻 RNase活性显著提高,RNA含量降低,RNase活性与蛋白质含量呈显著负相关,RNA降解是缺Zn水稻蛋白质合成受阻的原因[18].本试验中,Zn浓度在150 mg/kg条件下叶绿素含量、地上部和根系可溶性糖含量、可溶性蛋白质含量均高于CK,但之后开始下降,且整体上随着Zn浓度的增大而呈现降低的趋势.本试验叶绿素的研究结果与李翠等[16]大油芒 (Spodiopogon sibiricus)的研究,李铮铮等[19]对鱼腥草(Houttuynia cordata)的研究,努扎艾提·艾比布[15]对香根草(V.zizanioides Nash)的研究,崔大练等[20]对田菁(Sesbania cannabina)及蔡卓等[21]在对无芒雀麦(Bromus inermis)等的研究结果是一致的.可溶性糖和蛋白质研究与努扎艾提·艾比布等[15]在香根草上,杨飞红等[22]在油菜(Brassica chinensis L.)上的研究结果基本相符.
3.3 不同浓度重金属Zn处理后金盏菊植株体内重金属Zn的分布
重金属在植物体内的分布,大多数为地上部含量小于根系含量,如香根草[15]、大油芒、狼尾草、柳枝稷[16]及油菜[22].本试验也得到了类似的结果,并随Zn浓度增大而呈现增高的趋势.但有学者认为作为重金属超积累的植物则应该是地上部重金属含量大于根系重金属含量,这样对于被重金属污染的土壤用植物修复方法才更具意义[23].徐卫红等[23]在黑麦草上的研究得出,黑麦草地上部和根系Zn含量均超过了CK,且随着Zn浓度升高呈现增加趋势.Zn污染使油菜幼苗叶片及根中的Zn质量分数明显增加,根部的积累尤其明显.本试验中各种浓度Zn的运转系数(S/R)虽然低于1,但其植物体内重金属Zn的积累量还是比较高,且金盏菊因其生物量大,生长迅速,经田间试验后还是有望成为修复重金属污染土壤的植物之一.
[1]Bin H E,Yun Z J,Shi J B,et al.Research progress of heavy metal pollution in China:Sources,analytical methods,status,and toxicity[J].Chinesse Science Bulletin Kexue Tongbao,2013,58(2):134-140.
[2]韦朝阳,陈同斌.重金属超富集植物及修复技术研究进展[J].生物学报,2001,21(7):1196-1203.
[3]徐慧,张银龙.重金属污染废弃地修复植物种类的筛选与评价[J].污染防治技术.2009,22(1):44-48,55.
[4]陶星星.两个典型矿区植物积累重金属调查与研究[D].长沙:湖南农业大学,2008.
[5]朱佳文.湘西花垣铅锌矿区重金属污染土壤生态修复研究[D].长沙:湖南农业大学,2012.
[6]陆彬斌.海南昌化铅锌矿废弃地土壤重金属含量与优势植物富集特征[D].海口:海南师范大学,2013.
[7]陈红林.汉源铅锌矿区 Pb、Zn、Cd重金属富集植物筛选研究[D].成都:四川农业大学,2009.
[8]叶海波,杨肖娥,何冰,等.东南景天对锌、镉复合污染的反应及其对Zn、镉的吸收和积累特性[J].植物学报,2003,45(9):1030-1036.
[9]李合生,孙群,赵世杰.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:172-174.
[10]郝再彬,苍晶,徐仲.植物生理实验[M].哈尔滨:哈尔滨工业大学出版社,2004:46-49.
[11]国家技术监控局.土壤质量铜、锌的测定 火焰原子吸收分光光度法:GB/T17138-1997,GB/T17139-1997[S].北京:中国人民共和国国家标准,1997.
[12]李涛,蒲韵婷,王全华,等.Mn、Cu 和 Zn 在植物生长发育中的生理作用[J].河北农业科学,2008,12(6):12-15.
[13]丁海东,万延慧,齐乃敏,等.重金属 (Cd2+、Zn2+)胁迫番茄幼苗抗氧化酶系统的影响[J].上海农业学报,2004,20(4):79-82.
[14]金新文,赵扬,杜刚.不同浓度锌对西葫芦幼苗生长及其同工酶的影响[J].石河子大学学报 (自然科学版),1998,2(1):12-16.
[15]努扎艾提·艾比布,刘云国.重金属Zn、Cu对香根草生理生化指标的影响及其积累特性研究[J].农业环境科学学报,2010,29(1):54-59.
[16]李翠,王庆海,肖波.3种草本能源植物对 Zn胁迫的生理响应与积累特性[J].草业科学,2013,30(10):1555-1560.
[17]孙健,铁柏清,钱湛,等.部分生理生化指标对土壤复合重金属胁迫的响应[J].环境污染与防治,2006,28(9):666-670.
[18]李延,黄毅斌.缺锌对水稻蛋白质合成的影响[J].福建省农科院学报,1996,11(1):22-24.
[19]李铮铮,伍钧,唐亚,等.铅、锌及其交互作用对鱼腥草(Houttuynia cordata)叶绿素含量及抗氧化酶系统的影响[J].生态学报,2007,27(12):5441-5446.
[20]崔大练,马玉心,俞兴伟.重金属 Zn2+·Cd2+对田菁生理生化指标的影响[J].安徽农业科学,2012,40(1):376-378.
[21]蔡卓,毛培春,田小霞,等.无芒雀麦对 Cd和 Zn胁迫的生理响应及富集作用[J].草业科学,2011,28(6):876-882.
[22]杨红飞,王友保,李建龙.铜、锌污染对水稻土中油菜(Brassica chinensis L.)生长的影响及累积效应研究[J].生态环境学报,2011,20(10):1470-1477.
[23]徐卫红,王宏信,李文一,等.重金属富集植物黑麦草对 Zn的响应[J].水土保持学报,2006,20(3):43-46.
