褪黑素诱导sts基因表达提高葡萄白藜芦醇含量
2018-05-23许丽丽岳倩宇卞凤娥姚玉新
许丽丽,岳倩宇,卞凤娥,翟 衡,姚玉新*
(山东农业大学园艺科学与工程学院,山东 泰安 271000)
褪黑素不仅是一种广谱性抗氧化剂,而且具有激素功能,其存在于葡萄果皮、果肉和种子中[1-3]。褪黑素作为抗氧化剂能提高植物的抗旱、抗寒和抗盐等能力[4-6]。作为激素、褪黑素和生长素IAA具有相似的功能,低浓度促进生长而高浓度抑制生长[7],褪黑素具有类似IAA的生长抑制效应[8-9],水稻内源褪黑素水平和根系生长具有直接相关性[10]。尽管褪黑素和IAA具有相似的功能,但是RNA-Seq等数据表明褪黑素对根系生长的调节不依赖于IAA,二者的功能行使相互独立[11-13]。此外,褪黑素作为激素能改变植物器官发生模式,比如褪黑素处理能诱导黄瓜[14]、樱桃[15]和石榴[16]不定根和侧根的发生。褪黑素能够改变果实品质,褪黑素处理能增加番茄果实番茄红素、可溶性固形物和有机酸等含量,促进果实软化、提高水溶性果胶含量[17-18]。蛋白质组学分析进一步揭示了褪黑素介导的番茄果实成熟的生理和分子机制[19],241 个受褪黑素诱导的蛋白分别参与不同的果实发育代谢途径,其中,有8 个蛋白参与花青苷代谢途径。同时,研究学者发现褪黑素处理延迟了桃采后果实衰老[20];转色前褪黑素处理提高了葡萄果实单果质量和果粒成熟一致性,改善了葡萄酒香味[21]。因此,褪黑素对包括果实发育在内的多个生物学过程具有调控作用。
白藜芦醇属于非黄酮类酚类化合物,也是一种强的生物活性物质[22],具有抗氧化性和预防癌症、心血管等疾病的功能[23-24],鲜食葡萄是白藜芦醇的重要食品来源[25]。茋类合酶(stilbene synthase,STS)是白藜芦醇生化合成的关键酶;内源sts基因表达水平和葡萄白藜芦醇积累水平相一致[26],过量表达sts基因能够提高葡萄的白藜芦醇含量[27-28]。sts属于查尔酮合酶家族,葡萄上sts家族至少包含40 个成员[26,29-30]。葡萄sts家族成员具有高度保守的基因结构和高的蛋白相似度,系统进化上可以分为3 个组,A组位于染色体10上,B和C组以基因簇的形式串联排列在染色体16上[29-30]。sts成员在葡萄不同组织和生物、非生物胁迫下具有不同的转录模式,不同成员可能响应不同的生物学现象来提高白藜芦醇含量[29-30]。尤其是葡萄果实中白藜芦醇的合成和积累与脱落酸(abscisic acid,ABA)浓度相关,而ABA在果实成熟过程中起关键作用[23]。以上可知,白藜芦醇的合成受sts基因家族控制,不同的sts基因在响应上游信号控制白藜芦醇合成的角色目前还不清楚。
本研究旨在阐明褪黑素作为生长调节物质是否能够通过调节sts基因表达来促进葡萄中白藜芦醇的积累,研究结果对于应用褪黑素提高葡萄的营养保健价值具有重要实践意义,同时也为探索褪黑素调控白藜芦醇积累的分子机制提供理论支持。
1 材料与方法
1.1 材料与试剂
实验材料为田间7 a生鲜食与酿酒兼用葡萄品种‘摩尔多瓦’(Vitis vinifera×labrusca, cv. ‘Moldova’)和‘克瑞森无核’(Vitis vinifera cv. ‘Crimson seedless’)组培苗。培养基为MS基本培养基加吲哚丁酸(终质量浓度为0.2 mg/L),培养温度为(28±1)℃,光照强度为2 000~2 400 lx,光照时间12~14 h。
MS基本培养基 青岛高科技生物园海博生物有限公司;吲哚丁酸 索莱宝生物科技有限公司。
1.2 仪器与设备
RXZ恒温恒湿培养箱 宁波江南仪器厂;RE-2003型真空旋转蒸发仪 上海科升仪器公司;Prominenece型高效液相色谱仪 日本岛津公司;ABI7500型实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction,qRT-PCR)仪 美国应用生物系统公司。
1.3 方法
1.3.1 前处理及取样
在葡萄转色前期用100 µmol/L的褪黑素添加表面活性剂(2%曲拉通)浸泡果实,每穗浸泡10 s,浸泡3 次,每个处理选取20 穗果实,对照除将褪黑素溶液换成清水外,其他处理方式相同;取果实(去种子)用于提取RNA和白藜芦醇。将长势一致、继代培养1 个月的45 株处理组和45 株对照组组培苗同时平均分成12 组(每个重复3 棵),处理组将组培苗根系浸泡在30 μmol/L的褪黑素溶液(灭菌)中,而对照组将褪黑素溶液换为灭菌水,然后分别于不同时间点取样。所有的样品材料液氮速冻后,置-80 ℃低温冰箱保存。
1.3.2 RNA-Seq及数据分析
本部分实验由北京安诺基因协助完成。方法简要如下:利用Qubit®3.0 Flurometer检测RNA纯度,利用RNA Nano 6000检测RNA完整度和浓度。取2 µg RNA,利用NEBNext®Ultra™ RNA Library Prep Kit for Illumina®构建RNA文库,具体步骤参照试剂盒说明书。利用Qubit®RNA Assay Kit检测文库RNA质量浓度,稀释至1 ng/µL;利用Agilent Bioanalyzer 2100 system评价插入大小。然后利用HiSeq PE Cluster Kit v4-cBot-HS (Illumina)对文库基因进行聚类,最后构建好的文库利用Illumina Hiseq 4000平台测定基因两端各150 bp的序列。单基因表达水平利用RPKM来计算,log2(倍数变化)大于1和P值小于0.05作为显著差异表达的阈值。本实验每个样品有3 个实验重复,其平均值用于热图制作。利用在线工具制作热图(http://www.omicshare.com/tools/Home/Soft/heatmap)。
1.3.3 系统进化树的构建
使用ClustalX软件对葡萄中30 个STS蛋白进行氨基酸序列及该基因家族启动子序列比对,比对结果通过MEGA5软件生成进化树,进化树的构建采用邻接法(Neighbor-Joining,NJ),校验参数Bootstrap重复1 000 次[31]。
1.3.4 顺式作用元件分析
利用启动子在线预测网站(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析启动子上可能结合的顺式作用元件,并借助Adobe Photoshop 13.0.0绘图软件完成启动子各个结合元件的绘画。
1.3.5 qRT-PCR表达分析
各组织RNA提取使用的RNeasy plant mini kit是QIAGEN公司产品(货号:74903),cDNA第1条链的合成采用宝生物(大连)公司产品PrimeScriptTMRT reagent Kit with gDNA Eraser(货号:DRR047A)。各sts基因的qRT-PCR引物如表1所示,选取actin为内参基因,所有qRT-PCR均设置3 次重复。Ultra SYBR Mixture购自康为试剂公司。所得数据采用2-ΔΔCT进行计算分析[32]。
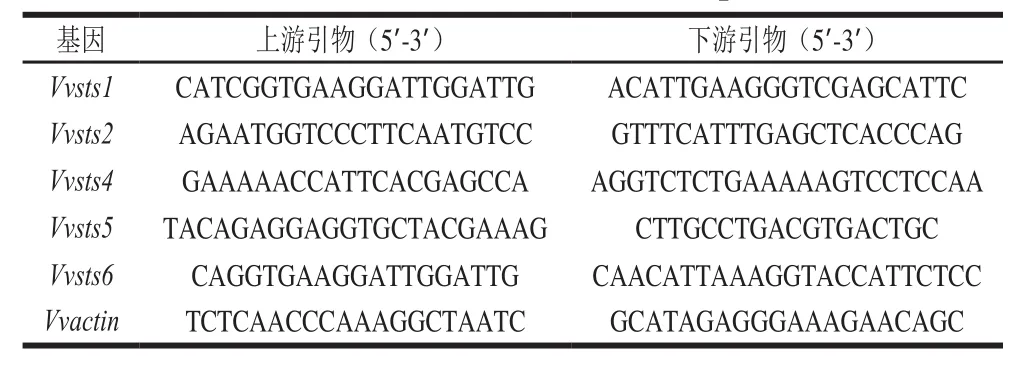
表1 qRT-PCR引物序列Table 1 Primers used for fluorescent qRT-PCR
1.3.6 白藜芦醇含量的测定
白藜芦醇含量的测定参考韩雅珊等[33]所描述方法并改进:取植物材料在液态氮冷冻,研磨,于1%盐酸-甲醇提取24 h后,5 000×g离心10 min,收集上清液,沉淀部分用1%盐酸-甲醇重复提取2次,离心后收集上清液,定容至50 mL,采用真空旋转蒸发仪(40±2)℃浓缩至2 mL,过0.45 μm微孔滤膜,-40 ℃保存备用。采用高效液相色谱测定,检测条件:LC-10AT输液泵,SPD-10A紫外检测器,安捷伦C18柱(250 mm×4.6 mm),柱温30 ℃;流动相A为乙腈,流动相B为水(乙酸调pH值至2.6),流速0.500 mL/min;进样方式为自动进样,进样体积10 µL;检测波长280 nm。
2 结果与分析
2.1 褪黑素处理对sts家族及pal基因表达的影响
为研究褪黑素处理对sts家族基因表达的直接影响,检测了褪黑素处理6 h后葡萄果实的转录组变化。结果表明,与对照相比,351 个基因被显著上调,18 个基因被显著下调;其中30 个sts基因被上调,占到总上调基因的8.5%,占目前已发表的葡萄sts基因总数的62.5%;未检测到下调的sts基因,表明sts家族基因成员广泛受褪黑素诱导(图1)。上调幅度分布在1.5~2.3 倍之间,VIT_216s0100g01130、VIT_216s0100g00780、VIT_210s0042g00890和VIT_216s0100g00810上调幅度超过2 倍。此外,转录组分析还发现VIT_200s2849g00010、VIT_216s0039g01360、VIT_216s0039g01130、VIT_216s0039g01300、VIT_208s0040g01710和VIT_216s0039g01100这6 个白藜芦醇合成相关的苯丙氨酸氨裂解酶(phenylalanine ammonia-lyase,PAL)基因pal表达被显著上调。

图1 褪黑素诱导的sts基因在不同组织的表达Fig. 1 Expression patterns of the melatonin-induced sts genes in different tissues
2.2 褪黑素诱导的sts基因序列及表达特点
30 个受褪黑素诱导的基因包含了定位于10号染色体的5 个sts,即VIT_216s0100g00810、VIT_216s0100g00920、VIT_216s0100g00840、VIT_216s0100g00930和VIT_216s0100g00910;其余25 个sts均定位在16号染色体上。其中,24 个sts基因具有完整的序列信息,氨基酸序列高度相似,达88.3%(图2)。根据蛋白相似度,24 个STS蛋白可用分为A、B、C组;A组包含12 个STS,蛋白同源性高达91.0%,其中VIT_216s0100g00910和VIT_216s0100g00830同源性最高为99.7%,仅有1 个氨基酸差异;B组仅含有1 个STS;C组含有11 个STS,同源性为96.6%,其中VIT_216s0100g00930和VIT_216s0100g00850同源性最高为99.2%,仅含3 个氨基酸差异(图2)。
30 个sts基因在不同组织中具有不同的表达模式(图1)。就愈伤、根、茎和叶而言,绝大部分sts基因在愈伤和根中具有较高的表达水平,尤其是VIT_216s0100g01100表达水平为参比的8 倍以上;果实中,不同的sts表达水平差异也较大,表达量最高的为VIT_216s0100g00830、VIT_216s0100g01020、VIT_216s0100g01170。并且,遗传距离越近,表达模式越相似,比如,VIT_2160100g00910和VIT_216s0100g00830具有完全相同的表达模式(图1、2)。
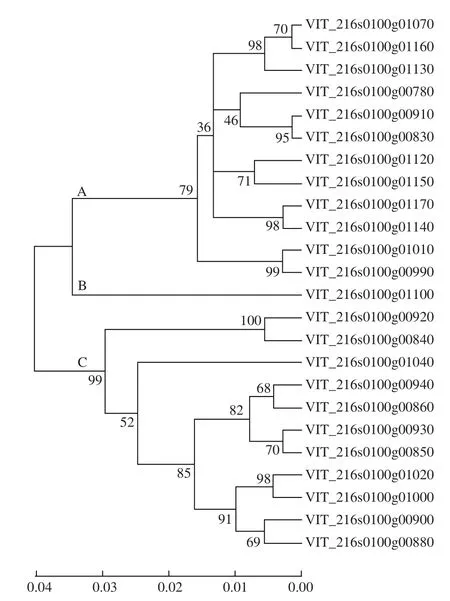
图2 褪黑素诱导的sts基因编码蛋白聚类分析Fig. 2 Clustering analysis of the STS proteins encoded by the melatonin-induced sts genes
2.3 sts基因启动子序列分析
5’-UTR上游1 500 bp的启动子序列分析表明,24 个sts基因分为A、B、C 3组,A组含有13 个sts,序列相似度为55.6%,其中有4 个sts对应了蛋白聚类的A1组;C组含有9 个sts基因,序列相似度为56.1%,完全属于蛋白聚类C组(图2、3);A组中,VIT_216s0100g00910和VIT_216s0100g00830序列相似度最高,对应了其最高的蛋白相似度;同样的现象也出现在VIT_216s0100g00930和VIT_216s0100g00850两个基因上(图2、3)。因此,总体上看,高的STS蛋白序列对应了高的启动子序列。在A组中,在启动子基本核心元件以外发现了一个高度保守的序列,即GG(T/C)TGTTGAG;在B组上也发现了一个基本元件外的保守序列AAGTGGATGAG(A/G)GTTGGTGA。除A1亚组外均含有不同数量的ABA响应元件(ABA response element,ABRE),尤其是B组含有3 个ABRE;A、B组大部分成员均含有乙烯响应元件(ethylene response element,ERE),而C组除VIT_216s0100g00880外均不含ERE;此外,在4 个sts启动子上发现了生长素响应元件(AuxRR-core)等(图3)。
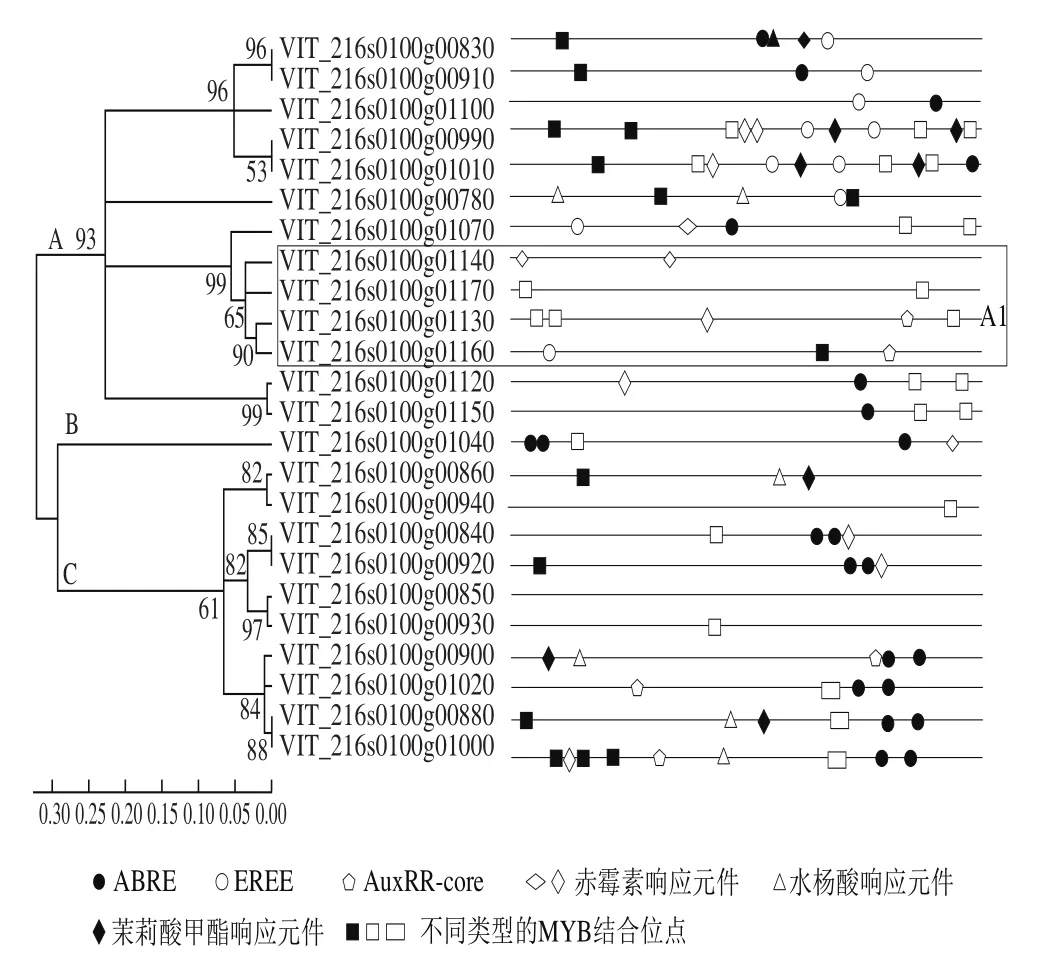
图3 sts基因启动子顺式作用元件分析Fig. 3 Cis acting elements of sts genes in the promoter regions
2.4 果实中sts基因对褪黑素处理的表达响应
为验证转录组数据的可靠性和进一步分析sts基因对褪黑素处理的表达响应,利用qRT-PCR检测了果实中5 个sts基因在褪黑素处理不同时间的表达水平。VIT_216s0100g00990在果实中受褪黑素强烈诱导,在处理后8 d表达为对照的4.7 倍(图4D)。相似地,VIT_216s0100g00780和VIT_216s0100g01130在果实中受褪黑素持续诱导,在处理后期表达水平分别为对照的2.7 倍和2.65 倍(图4B、E)。相比之下,VIT_216s0100g01150和VIT_216s0100g00810在果实中轻微受褪黑素诱导,增幅在1 倍以内(图4A、C)。由此可知,5 个sts基因不同程度受褪黑素诱导,与转录组分析基本一致。
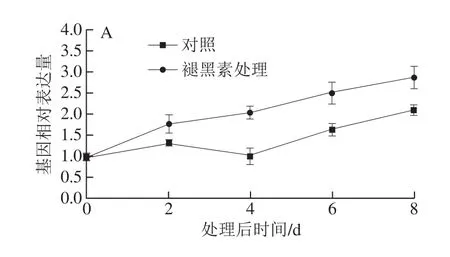
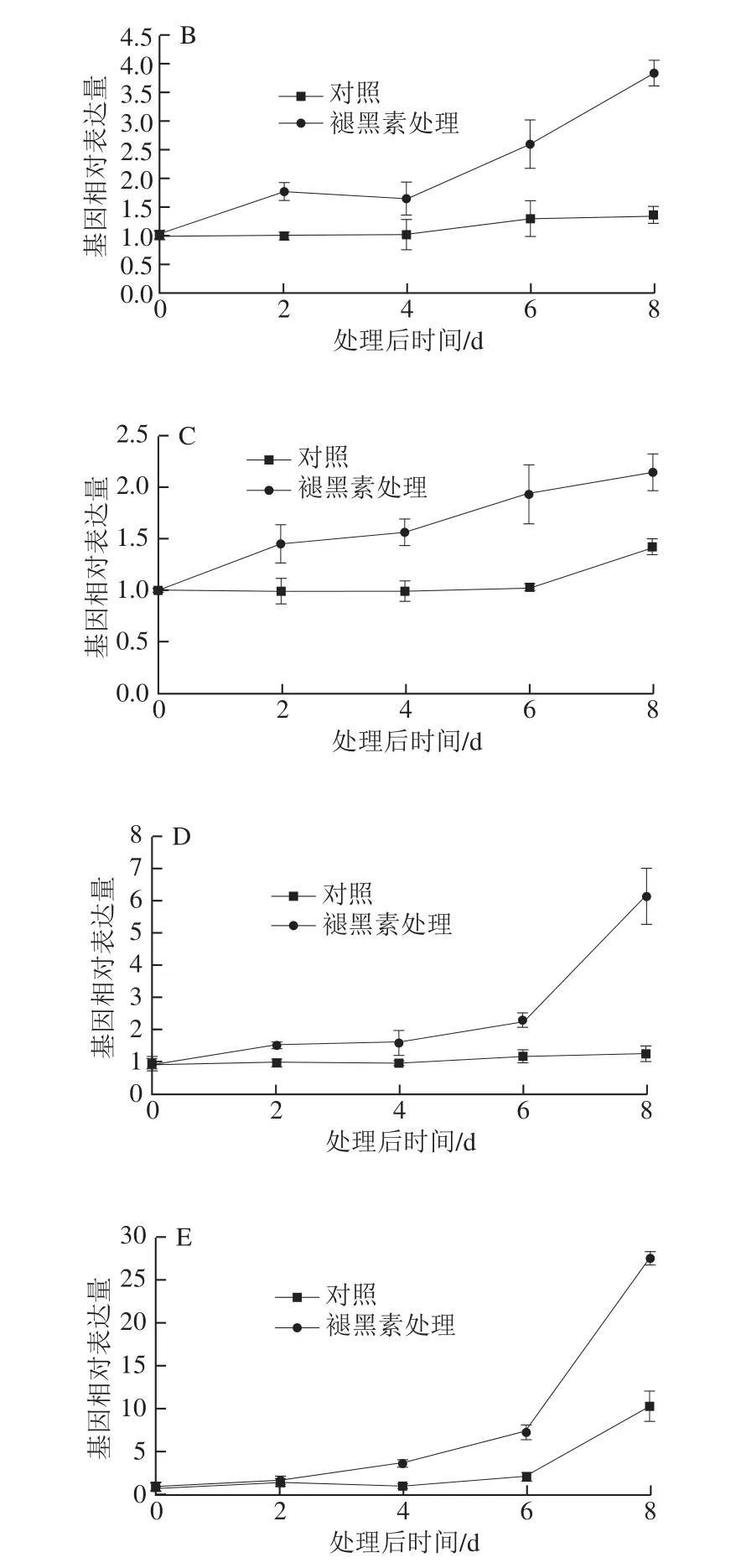
图4 褪黑素处理不同时间点sts基因在果实中的表达水平Fig. 4 Expression profiles of the sts genes in fruits at different days after melatonin treatment
2.5 根系中sts基因对褪黑素处理的表达响应
考虑到sts基因在根系中具有较高的表达水平,利用qRT-PCR检测了根系中5 个sts基因在褪黑素处理不同时间的表达水平。VIT_216s0100g01150和VIT_216s0100g01130根系中受褪黑素强烈诱导,在处理后期表达水平分别为对照的3.8 倍和3.35倍(图5A、E);VIT_216s0100g00780在根中也受褪黑素持续诱导,后期为对照的2.7 倍左右(图5B);相比之下,VIT_216s0100g00810和VIT_216s0100g00990在根中对褪黑素响应不明显(图5C、D)。

图5 褪黑素处理不同时间点sts基因在根系中的表达水平Fig. 5 Expression profiles of the sts genes in root at different days after melatonin treatment
2.6 外源褪黑素处理对白藜芦醇含量的影响
为了进一步阐明褪黑素处理对白藜芦醇含量的影响,检测了褪黑素处理后不同时间果实白藜芦醇的含量,结果表明,在褪黑素处理后42、63、77 d(果实成熟期),褪黑素均明显提高了果实白藜芦醇含量,较对照分别增加了0.90、1.12 倍和1.18 倍(图6A)。此外,还检测了褪黑素处理后3 d葡萄根系中的白藜芦醇含量,结果表明,褪黑素处理大幅度提高了根系白藜芦醇含量,为对照的3.18 倍(图6B)。以上研究表明,褪黑素处理葡萄果实和根系,均能提高其白藜芦醇含量。
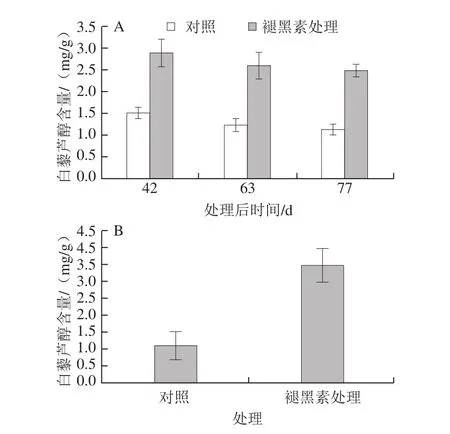
图6 褪黑素处理对果实(A)和根系(B)白藜芦醇含量的影响Fig. 6 Impacts of melatonin treatment on resveratrol contents in fruit (A)and root (B)
3 讨 论
目前,褪黑素对果实品质的影响,尤其是对包括白藜芦醇在内的次生代谢产物的调节还不清楚。葡萄果实中,褪黑素的积累高峰出现在转色早期[34-35],暗示了褪黑素对果实成熟可能具有重要调控作用,因此对转色初期的葡萄果实进行褪黑素处理,来研究其对白藜芦醇含量及其合成基因sts表达的影响。本研究中,褪黑素处理导致30 个sts基因表达上调,同时提高了白藜芦醇含量。相似地,在葡萄组培苗根系中高sts表达对应了高的白藜芦醇含量[36]。在检测的5 个sts基因中,仅VIT_216s0100g01130在正常条件下随果实发育表达明显上调,表明了不同sts基因在果实发育过程中可能具有不同的作用或者对白藜芦醇合成具有不同的贡献;相比之下,在‘Corvina’葡萄果实中,未检测到受果实成熟诱导的sts基因[29]。研究还发现成熟的‘Pinot noir’葡萄果实积累大量的白藜芦醇(20 µg/g)[37],而成熟的‘Corvina’葡萄果实只积累了1.5 µg/g的白藜芦醇[38]。因此,sts表达水平对白藜芦醇的合成具有重要作用,葡萄果实中sts表达和白藜芦醇的积累可能与葡萄基因型有关。
褪黑素对sts的诱导可能基于以下几个方面。首先,研究已证实褪黑素可以作为信号分子引起细胞核转录再编程[2,39],因此褪黑素可能通过核转录再编程来调节sts基因表达。其次,褪黑素处理能影响ABA、乙烯含量[19,40],水杨酸处理可以显著提高葡萄品种果皮中的白藜芦醇含量[41];本研究中,在褪黑素诱导的sts基因上发现了ABA、水杨酸、乙烯等激素响应元件(图2),暗示了sts基因可能受以上激素调控;因此,褪黑素可能通过其他激素来间接调控sts基因的表达。再者,研究表明褪黑素处理能通过上调pal等基因的表达提高花青素含量[42];本研究中褪黑素处理上调了5 个pal表达;因此,褪黑素处理可能通过诱导pal基因表达来提高白藜芦醇合成所需要的上游底物,进而促进了sts基因表达。
葡萄上绝大多数sts基因分布在16号染色体500 kb的区间内,并且在编码和非编码区都存在大量的同源区域,暗示了sts家族主要来自于串联重复和片段重复复制[29]。本研究30 个褪黑素诱导的STS蛋白和启动子序列具有高度相似性,并且在启动子区发现了高度保守的基元,暗示了这些sts可能具有相同的褪黑素响应元件。在褪黑素诱导的30 个sts基因中,仅个别存在IAA响应元件,暗示了褪黑素和IAA可能具有不同的靶基因调节和信号转导途径[2]。葡萄基因组大小为487 Mb,其中85 Mb来自基因组重复复制,尤其是染色体16,重复复制率达25.08%[43]。葡萄基因组在几个次生代谢相关的基因家族上发生了显著的复制、扩张[44];此类基因的拷贝数的增加是植物调节性进化的主要动力之一[45-46]。就sts基因而言,葡萄sts家族比其他作物sts家族大10 倍[30];但在单子叶高粱上sts以单拷贝存在[47],暗示了sts多拷贝事件可能发生在单、双子叶植物分化之后。并且,葡萄上至少32 个sts基因具有合成白藜芦醇的功能,不同sts的功能可能具有冗余性[30];但本研究中不同处理时间的表达响应表明,除了功能冗余外不同的sts对褪黑素诱导的白藜芦醇提高可能具有不同的作用。
4 结 论
葡萄sts基因具有保守的启动子和蛋白质序列。大量的葡萄sts基因在果实中能响应褪黑素处理,但具有不同的响应模式。褪黑素处理诱导了sts基因的表达进而提高了组织中白藜芦醇含量。
参考文献:
[1]IRITI M. Melatonin in grape, not just a myth, maybe a panacea[J].Journal of Pineal Research, 2009, 46(3): 353.
[2]ARNAOM B, HERNÁNDEZ-RUIZ J. Melatonin: plant growth regulator and/or biostimulator during stress[J]. Trends in Plant Science, 2014, 19(12): 789-797. DOI:10.1016/j.tplants.2014.07.006.
[3]SUN Q, ZHANG N, WANG J, et al. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life[J]. Journal of Experimental Botany, 2015, 66(3): 657-668. DOI:10.1093/jxb/eru332.
[4]ARNAO M B. Phytomelatonin: discovery, content, and role in plants[J].Advances in Botany, 2014, 2: 1-7. DOI:10.1155/2014/815769.
[5]JANAS K M, POSMYK M M. Melatonin, an underestimated natural substance with great potential for agricultural application[J]. Acta Physiologiae Plantarum, 2013, 35(12): 3285-3292. DOI:10.1007/s11738-013-1372-0.
[6]ARNAO M B, RUIZ J H. Melatonin: possible role as light-protector in plants[J]. Physics Research & Technology Series, 2014: 79-92.
[7]HERNÁNDEZ R J, CANO A, ARNAO M B. Melatonin: a growthstimulating compound present in lupin tissues[J]. Planta, 2004, 220(1):140-144. DOI:10.1007/s00425-004-1317-3.
[8]HERNÁNDEZ R J, CANO A, ARNAO M B. Melatonin acts as a growth-stimulating compound in some monocot species[J]. Journal of Pineal Research, 2005, 39(2): 137-142. DOI:10.1111/J.1600-079X.2005.00226.X.
[9]POSMYK M M, KURAN H, MARCINIAK K, et al. Presowing seed treatment with melatonin protects red cabbage seedlings against toxic copper ion concentrations[J]. Journal of Pineal Research, 2008, 45(1):24-31. DOI:10.1111/j.1600-079X.2007.00552.x.
[10]PARK S, BACK K. Melatonin promotes seminal root elongation and root growth in transgenic rice after germination[J]. Journal of Pineal Research, 2012, 53(4): 385-389. DOI:10.1111/j.1600-079X.2012.01008.x.
[11]KOYAMA F C, CARVALHO T L, ALVES E, et al. The structurally related auxin and melatonin tryptophan-derivatives and their roles in Arabidopsis thaliana and in the human malaria parasite Plasmodium falciparum[J]. Journal of Eukaryotic Microbiology, 2013, 60(6):646-651. DOI:10.1111/jeu.12080.
[12]PELAGIO F R, MUÑOZ P E, ORTIZ C R, et al. Melatonin regulates Arabidopsis root system architecture likely acting independently of auxin signaling[J]. Journal of Pineal Research, 2012, 53: 279-288.DOI:10.1111/j.1600-079X.2012.00996.x.
[13]ZHANG N, ZHANG H J, ZHAO B, et al. The RNA-seq approach to discriminate gene expression profiles in response to melatonin on cucumber lateral root formation[J]. Journal of Pineal Research, 2014,56: 39-50. DOI:10.1111/jpi.12095.
[14]ZHANG N, ZHAO B, ZHANG H J, et al. Melatonin promotes waterstress tolerance, lateral root formation, and seed germination in cucumber (Cucumis sativus L.)[J]. Journal of Pineal Research, 2013,54: 15-23. DOI:10.1111/j.1600-079X.2012.01015.x.
[15]SARROPOULOU V N, THERIOS I N, DIMASSI T K N. Melatonin promotes adventitious root regeneration in in vitro shoot tip explants of the commercial sweet cherry rootstocks CAB-6P (Prunus cerasus L.),Gisela 6 (P. cerasus-P. canescens), and MxM 60 (P. avium-P. mahaleb)[J].Journal of Pineal Research, 2012, 52: 38-46. DOI:10.1111/j.1600-079X.2011.00914.x.
[16]SARROU E, THERIOS I, DIMASSI-THERIOU K. Melatonin and other factors that promote rooting and sprouting of shoot cuttings in Punica granatum cv. Wonderful[J]. Turkish Journal of Botany, 2014,38: 293-301. DOI:10.3906/bot-1302-55.
[17]SUN Q, ZHANG N, WANG J, et al. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life[J]. Journal of Experimental Botany, 2015, 66(3): 657-668. DOI:10.1093/jxb/eru332.
[18]LIU J, ZHANG R, SUN Y, et al. The beneficial effects of exogenous melatonin on tomato fruit properties[J]. Scientia Horticulturae, 2016,207: 14-20. DOI:10.1016/j.scienta.2016.05.003.
[19]SUN Q, ZHANG N, WANG J, et al. A label-free differential proteomics analysis reveals the effect of melatonin on promoting fruit ripening and anthocyanin accumulation upon postharvest in tomato[J].Journal of Pineal Research, 2016, 61(2): 138-153.
[20]GAO H, ZHANG Z K, CHAI H K, et al. Melatonin treatment delays postharvest senescence and regulates reactive oxygen species metabolism in peach fruit[J]. Postharvest Biology & Technology,2016, 118: 103-110. DOI:10.1016/j.postharvbio.2016.03.006.
[21]MENG J F, XU T F, SONG C Z. Melatonin treatment of pre-veraison grape berries to increase size and synchronicity of berries and modify wine aroma components[J]. Food Chemistry, 2015, 185: 127-134.DOI:10.1016/j.foodchem.2015.03.140.
[22]MONAGAS M, BARTOLOME B, GMEZ C C. Updated knowledge about the presence of phenoliccompounds in wine[J]. Critical Reviews in Food Science and Nutrition, 2005, 45(2): 85-118.DOI:10.1080/10408690490911710.
[23]WANG J F, WANG S Q, LIU G T, et al. The synthesis and accumulation of resveratrol are associated with veraison and abscisicacid concentration in Beihong (Vitis vinifera × Vitis amurensis)berry skin[J]. Frontiers in Plant Science, 2016, 7(212). DOI:10.3389/fpls.2016.01605.
[24]TOMÉC J, GONZLVEZ M, LARROSA M, et al. Resveratrol in primary and secondary prevention of cardiovascular disease: a dietary and clinical perspective[J]. Annals of the New York Academy of Sciences, 2013, 1290(1): 37-51. DOI:10.1111/nyas.12150.
[25]VANNOZZI A, BOSS P K, WALKER A R, et al. Identification of an R2R3 MYB transcription factor involved in the regulation of the stilbene synthase pathway in grapevine[J]. Arabian Journal of Geosciences, 2014, 8(7): 1-13. DOI:10.17660/ActaHortic.2014.1046.5.
[26]SHI J L, HE M Y, CAO J L, et al. The comparative analysis of the potential relationship between resveratrol and stilbene synthase gene family in the development stages of grapes (Vitis quinquangularis and Vitis vinifera)[J]. Plant Physiology & Biochemistry, 2014, 74: 24-32.DOI:10.1016/j.plaphy.2013.10.021.
[27]SERAZETDINOVA L, OLDACH K H, LRZ H. Expression of transgenic stilbene synthases in wheat causes the accumulation of unknown stilbene derivatives with antifungal activity[J]. Journal of Plant Physiology, 2005, 162(9): 985-1002. DOI:10.1016/j.jplph.2004.11.005.
[28]CHENG S Y, XIE X Q, XU Y, et al. Genetic transformation of a fruit-specific, highly expressed stilbene synthase gene from Chinese wild Vitis quinquangularis[J]. Planta, 2016, 243(4): 1041-1053.DOI:10.1007/s00425-015-2459-1.
[29]ALESSANDRO V, DRY I B, MARIANNA F, et al. Genome-wide analysis of the grapevine stilbene synthase multigenic family: genomic organization and expression profiles upon biotic and abiotic stresses[J].BMC Plant Biology, 2012, 12(1): 130-151. DOI:10.1186/1471-2229-12-130.
[30]PARAGE C, TAVARES R, RÉTY S, et al. Structural, functional, and evolutionary analysis of the unusually large stilbene synthase gene family in grapevine[J]. Plant Physiology, 2012, 160(3): 1407-1419.DOI:10.1104/pp.112.202705.
[31]王小非, 刘鑫, 苏玲, 等. 番茄LBD基因家族的全基因组序列鉴定及其进化和表达分析[J]. 中国农业科学, 2013, 46(12): 2501-2513.
[32]LEWIS A M, RICE K C. Quantitative real-time PCR (qPCR)workflow for analyzing Staphylococcus aureus gene expression[J].Methods in Molecular Biology, 2016, 1373: 143-154.DOI:10.1007/7651_2014_193.
[33]韩雅珊, 陈雷. 高效液相色谱法测定葡萄酒中的白藜芦醇[J]. 色谱,1999(4): 366-368.
[34]VITALINI S, GARDANA C, ZANZOTTO A. The presence of melatonin in grapevine (Vitis vinifera L.) berry tissues[J]. Journal of Pineal Research,2011, 51: 331-337. DOI:10.1111/j.1600-079X.2011.00893.x.
[35]MURCH S J, HALL B A, LE C H, et al. Changes in the levels of indoleamine phytochemicals during veraison and ripening of wine grapes[J]. Journal of Pineal Research, 2010, 49: 95-100. DOI:10.1111/j.1600-079X.2010.00774.x.
[36]AHN S Y, KIM S A, CHOI S J, et al. Comparison of accumulation of stilbene compounds and stilbene related gene expression in two grape berries irradiated with different light sources[J]. Horticulture Environment & Biotechnology, 2015, 56(1): 36-43.
[37]KORHAMMER S, RENIERO F, MATTIVI F. An oligostilbene from Vitis roots[J]. Phytochemistry, 1995, 38(6): 1501-1504.DOI:10.1016/0031-9422(94)00811-7.
[38]GATTO P, VRHOVSEK U, MUTH J, et al. Ripening and genotype control stilbene accumulation in healthy grapes[J]. Journal of Agricultural & Food Chemistry, 2008, 56: 11773-11785. DOI:10.1021/jf8017707.
[39]VERSARI A, PARPINELLO G P, TORNIELLI G B, et al. Stilbene compounds and stilbene synthase expression during ripening, wilting and UV treatment in grape cv. Corvina[J]. Journal of Agricultural &Food Chemistry, 2001, 92: 729-736. DOI:10.1021/jf010672o.
[40]LI X, ZHENG X, YAN S, et al. Effects of salicylic acid (SA),ultraviolet radiation (UV-B and UV-C) on trans-resveratrol inducement in the skin of harvested grape berries[J]. Frontiers of Agriculture in China, 2008, 2(1): 77-81. DOI:10.1007/s11703-008-0014-6.
[41]XU A, ZHAN J C, HUANG W D. Effects of ultraviolet C, methyl jasmonate and salicylic acid, alone or in combination, on stilbene biosynthesis in cell suspension cultures of Vitis vinifera L. cv. Cabernet Sauvignon[J]. Plant Cell Tissue and Organ Culture (PCTOC), 2015,122(1): 197-211.
[42]ZHANG H J, ZHANG N, YANG R C, et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems,ABA and GA4 interaction in cucumber (Cucumis sativus L.)[J].Journal of Pineal Research, 2014, 57(3): 269-279. DOI:10.1111/jpi.12167.
[43]ZHANG N, SUN Q, LI H, et al. Melatonin improved anthocyanin accumulation by regulating gene expressions and resulted in high reactive oxygen species scavenging capacity in cabbage[J]. Frontiers in Plant Science, 2016, 7: 197. DOI:10.3389/fpls.2016.00197.
[44]GIANNUZZI G D, ADDABBO P, GASPARRO M, et al. Analysis of high-identity segmental duplications in the grapevine genome[J]. BMC Genomics, 2011, 12: 436. DOI:10.1186/1471-2164-12-436.
[45]KEELING C I, WEISSHAAR S, LIN R P C, et al. Functional plasticity of paralogous diterpene synthases involved in conifer defense[J].Proceedings of the National Academy of Sciences of the United States of America, 2008, 105: 1085-1090. DOI:10.1073/pnas.0709466105.
[46]OBER D. Seeing double: gene duplications and diversification in plant secondary metabolism[J]. Trends in Plant Science, 2005, 10: 444-449.DOI:10.1016/j.tplants.2005.07.007.
[47]HA M, LI W H, CHEN Z J. External factors accelerate expression divergence between duplicate genes[J]. Trends Genetics Tig, 2007, 23:162-166. DOI:10.1016/j.tig.2007.02.005.
