濒危植物羽叶丁香种群结构与动态特征
2018-05-16姜在民和子森
姜在民,和子森,宿 昊,赵 涵,蔡 靖,4,*
1 西北农林科技大学生命学院,杨凌 712100 2 西北农林科技大学林学院,杨凌 712100 3 新疆维吾尔自治区林业有害生物防治检疫局,乌鲁木齐 830000 4 秦岭国家级生态定位站,杨凌 712100
种群是指在一定时间内占据一定空间的同种生物的所有个体的总和,它是物种进化的基本单位。种群结构与动态是种群生态学研究的核心问题之一[1- 2],尤其针对濒危植物而言,可以定量研究其濒危过程及濒危机理等[3]。种群结构包括径级结构、高度结构以及年龄结构等[4],种群结构是种群最基本的特征,它既可反映种群内不同大小个体数量的分布和配置状况,也能反映种群的数量动态及其未来发展与演变趋势[5]。种群生命表与其构建的存活曲线以及生存分析理论分析不仅可以反映种群目前的存活状态,而且还可以反映植物与环境之间的适合度[6- 7],结合种群动态量化分析和时间序列预测还可以阐明种群的受干扰状况及未来的发展趋势[8- 9]。
羽叶丁香是木犀科(Oleaceae)丁香属中唯一具有羽状复叶的植物,为中国特有种,国家三级保护植物[10]。羽叶丁香在丁香属系统演化及对我国植物区系的研究上,具有重要的学术意义[11]。内蒙古与宁夏交界的贺兰山是目前羽叶丁香的主产区,在宁夏中卫香山、陕西秦岭、青海循化、甘肃以及四川西部也有零星分布[12],常生于山坡灌丛,海拔1800—2800m。羽叶丁香不仅具有较高的观赏价值,而且其根茎作为中医和蒙医的名贵药材,具降气、温中、暖胃等功效[13]。但是由于生境恶化,加之人类活动频繁,羽叶丁香种群正在逐步缩小,甚至面临濒危风险,因此对其种群生态学的研究迫在眉睫。目前对于羽叶丁香的报道主要集中于种子萌发[14]、组织培养[15]、药用成分分析[16- 19]及光和生理[20]等方面,而有关其种群结构与动态方面的研究还未见报道。本文通过样地调查和对比分析,首次阐明羽叶丁香天然种群年龄结构特征,分析其生存现状和受威胁状况,利用种群数量变化动态指数及生存分析等数学模型,对种群未来发展趋势进行预测并揭示其形成原因,探索羽叶丁香的濒危机理,以期为羽叶丁香天然种群的保护和恢复提出合理的适应性策略,并为其所处群落及生态系统保护提供科学的决策依据。
1 研究区概况和研究方法
1.1 研究区概况
研究区甘沟(紫花沟)位于宁夏贺兰山区东麓地区,该区地理坐标(37°43′—39°23′N,105°45′—106°47′E),为贺兰山东麓冲积扇与黄河冲积平原之间的宽阔地带。南北长200km,东西宽5—30km,土地总面积2410.7km2。羽叶丁香主要分布于紫花沟尽头的半阴坡中下部或坡底以及水流冲积扇地带,地理坐标为38°31′8″—38°31′32″N,105°52′30″—105°53′51″E,海拔1811—1912m。该区属中温带干旱气候区,具典型的大陆性气候特点,光能资源丰富,热量适中,干旱少雨,昼夜温差大。年均温8.8℃,4月至10月≥10℃的有效积温为3300℃。年均降雨量200mm,年均蒸发量1470mm,为降雨量的7倍,全年日照时数2851—3106h,平均无霜期170d。该区土地成土母质以冲积物为主,土壤含砾石、砾粒,地形起伏小,以淡灰钙土为主,有机质含量0.4%—1.0%。主要地带性植被有旱榆(Ulmusglaucescens)、西北栒子(Cotoneasterzabelii)、黄蔷薇(Rosahugonis)、金露梅(Potentillafruticosa)、置疑小檗(Berberisdubia)、蒙古扁桃(Amygdalusmongolica)、蒙古绣线菊(Spiraeamongolica)、小叶忍冬(Loniceramicrophylla)、小叶鼠李(Rhamnusparvifolia)、虎榛子(Ostryopsisdavidiana)等。
研究区中卫香山位于宁夏中部干旱带土石山区,属黄土高原向鄂尔多斯高原过渡带,地貌呈现山地、沙质缓坡丘陵镶嵌分布。该区光照资源丰富,昼夜温差大,无霜期155d左右,年平均太阳总辐射量为6.0×109J/m2;年平均温度6.8℃,年日照时数在2800小时以上,≥10℃积温为2332℃;年平均降水量247.4mm,多集中于7—9月,且多暴雨,年蒸发量2100—2400mm,约为降水量的8—10倍。羽叶丁香分布区面积狭小,位于37°9′34″—37°9′38″N,105°13′56″—105°14′03″E之间,海拔2220—2265m,具有独特的山地小气候特征。由于降水稀少,干旱频发,风沙大,土壤侵蚀严重,故此地植被稀疏,伴生植物多为旱生或超旱生植物,主要有小叶忍冬(Loniceramicrophylla)、鬼箭锦鸡儿(Caraganajubata)、叉子圆柏(Sabinavulgaris)、青甘韭(Alliumprzewalskianum)、香青(Anaphalissinica)、冷蒿(Artemisiafrigida)等。
1.2 研究方法
1.2.1 样地设置与调查方法
本研究选择羽叶丁香在贺兰山甘沟的集中分布地紫花沟以及中卫香山地区作为调查取样地。一共设置10块样地,贺兰山甘沟6块,中卫香山4块,大小为20m×20m,样地之间的距离>100m,在每个样地中以10m为间隔设置成4个10m×10m的样方,共40个样方4000m2,记录样方内所有羽叶丁香的个体数、基径、高度、冠幅等指标。
1.2.2 径级划分
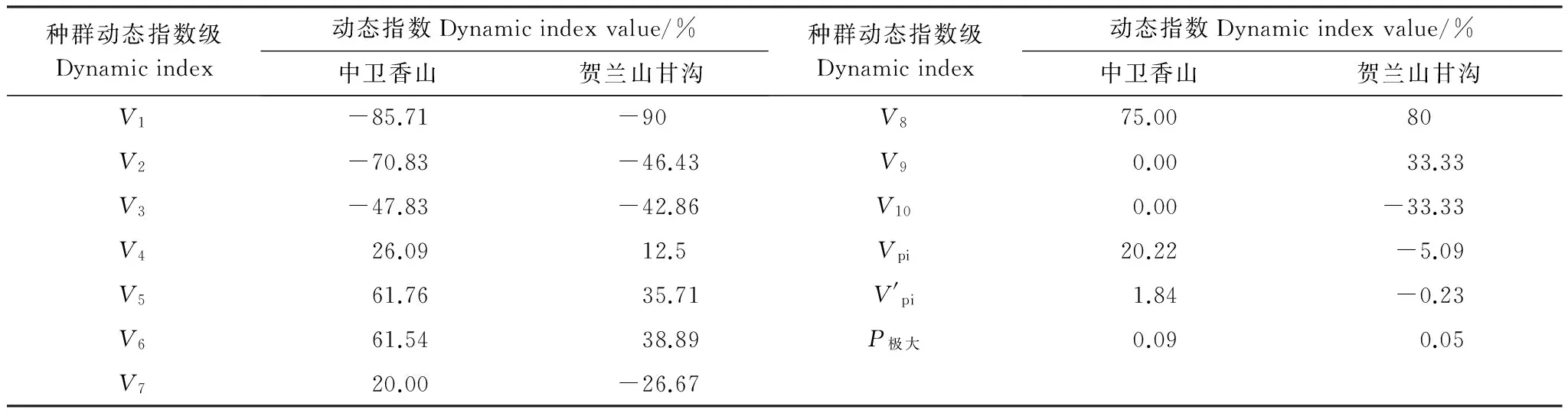
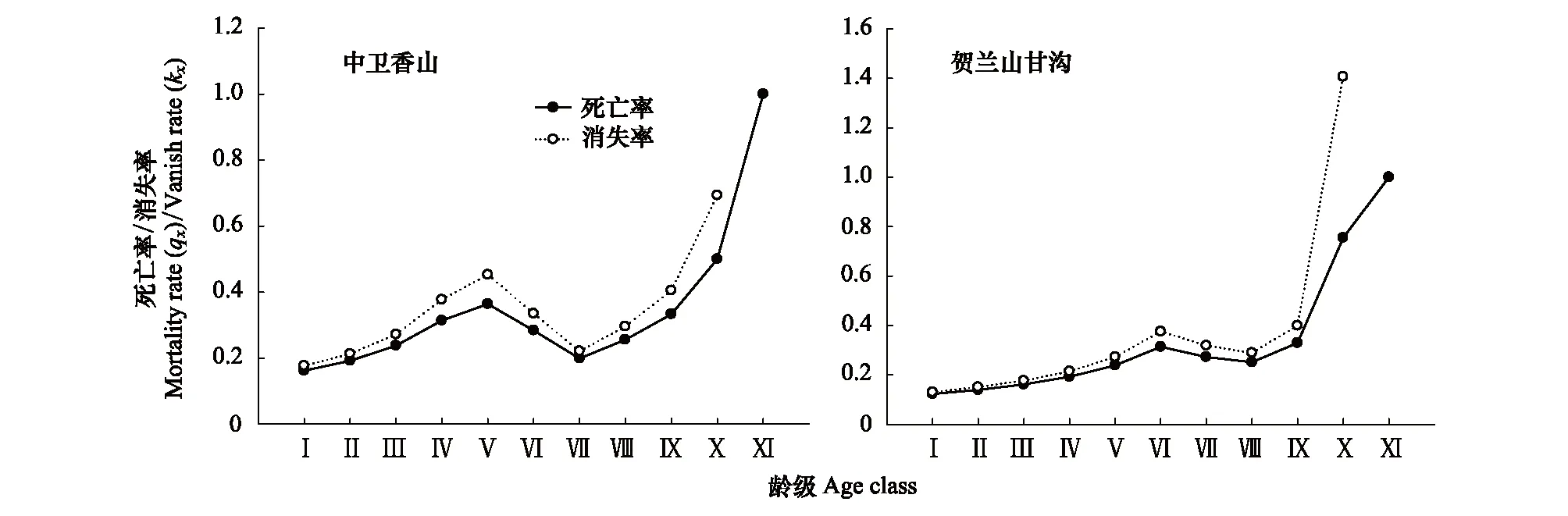
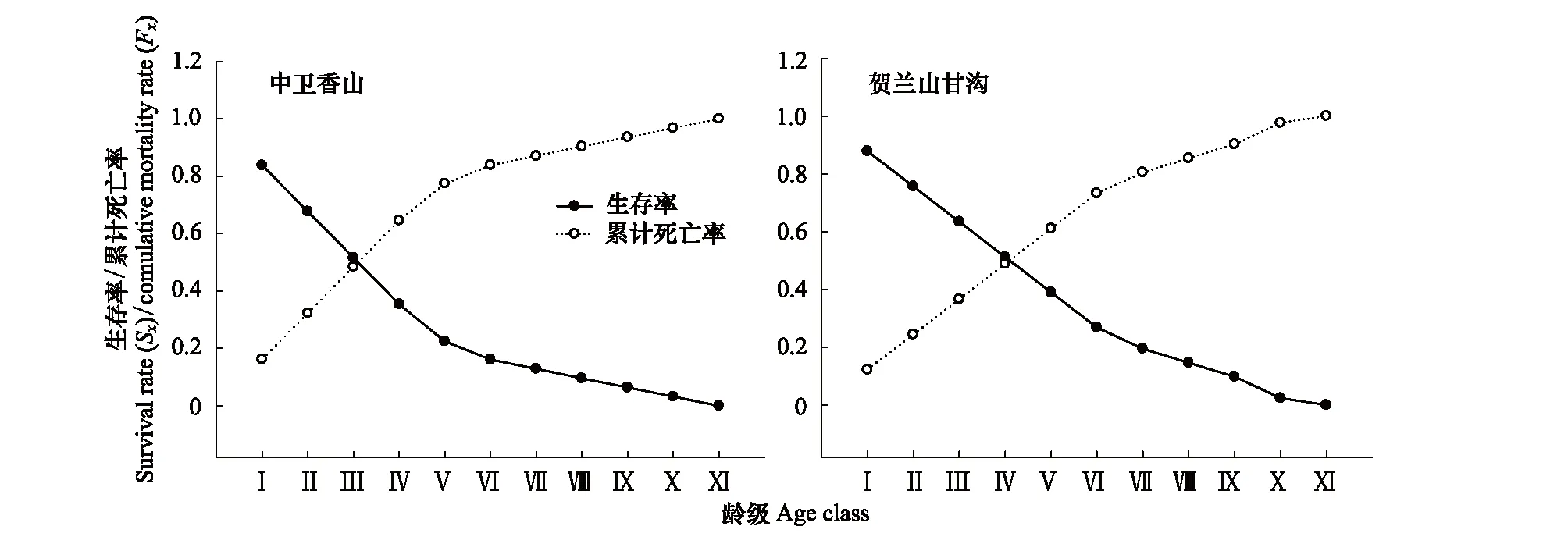
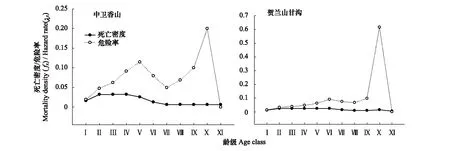
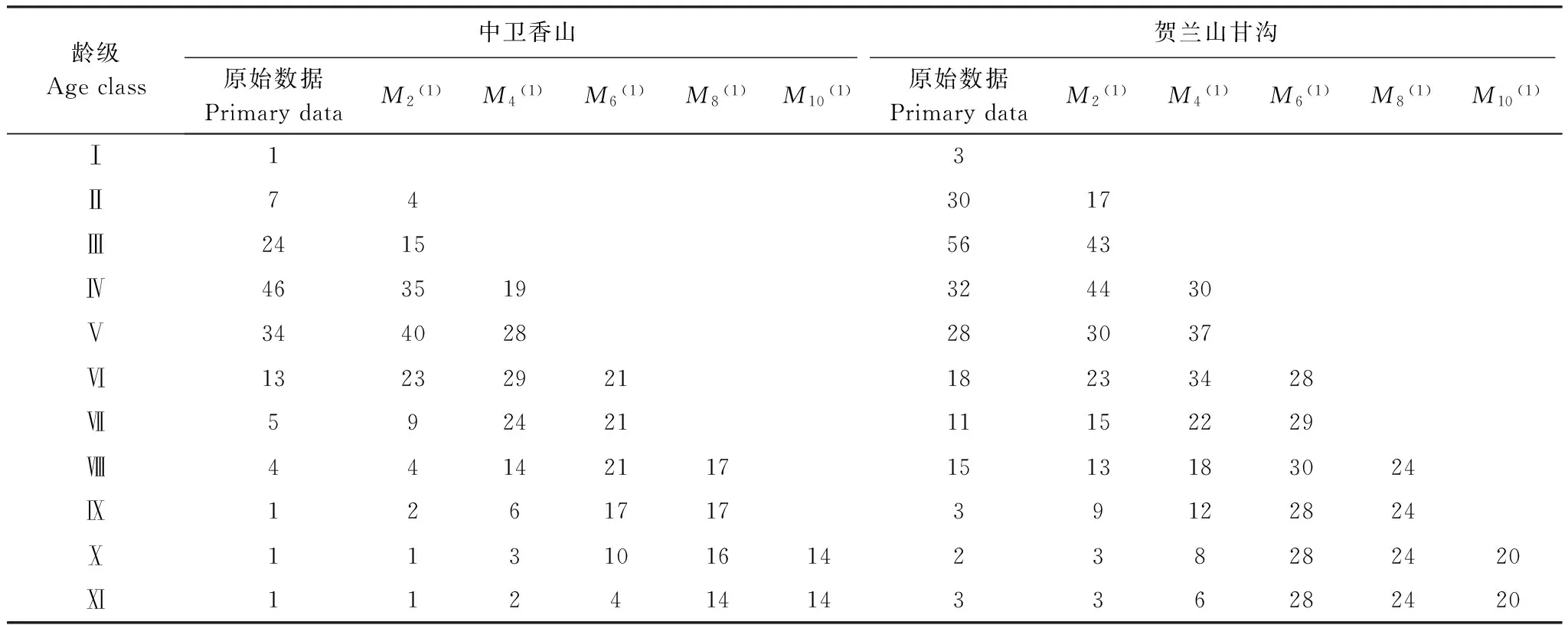
对于木本植物尤其濒危物种而言,大多数学者认为在数量较少,较难获取其年龄的情况下,可用径级结构代替其年龄结构[21]。因此,本研究采用基径大小作为标准划分径级结构,以此代替其年龄结构分析羽叶丁香种群的年龄结构特征。由于羽叶丁香为多分枝状灌木,在测基径时选取最粗枝干的基径大小作为该灌丛的基径大小。根据基径(BD,BaseDiameter)的分布状况,将基径划分为11个等级:Ⅰ级(BD<0.5cm)、Ⅱ级(0.5cm≤BD 1.2.3 种群动态量化方法 种群动态量化分析方法参考陈晓德[22]的种群与群落结构动态量化分析方法: (1) (2) 式中,Vn为种群从n到n+1级的个体数量变化;式(2)中Vpi为整个种群结构的数量变化动态指数(忽略外部干扰);Sn、Sn+1分别为第n和n+1级种群个体数;k为种群大小级数量。而当考虑未来的外部干扰时,Vpi还与年龄级数量(k)以及各龄级内的个体数(Sn)相关,可进一步将式(2)修正为: (3) (4) 1.2.4 种群静态生命表建立和生存分析 lx=(ax/a0)×1000 (5) dx=lx-lx+1 (6) qx=(dx/lx)×100% (7) lx=(lx+lx+1)/2 (8) (9) ex=Tx/lx (10) Kx=lnlx-lnlx+1 (11) 静态生命表又称特定时间生命表,多用于对长寿命的木本植物种群的统计研究,主要包括以下参数:Ax是在x龄级内现有个体数;lx是在x龄级开始时标准化存活个体数;dx是从x到x+1龄级间隔期内标准化死亡数;qx是从x到x+1龄级间隔期间死亡率;Lx是从x到x+1龄级间隔期间还存活的个体数;Tx是从x龄级到超过x龄级的个体总数;ex是进入x龄级个体的生命期望或平均期望寿命;Kx为消失率(损失度)。它们之间的计算关系如下: 由于静态生命表是用同一时期收集的种群所有个体的径级编制而成,反映了多个时代重叠的年龄动态历程中的一个特定时间,而不是对这一种群全部生活史的追踪[1],而且在调查中存在系统误差,在编制生命表时会出现死亡率为负的情况,这与生命表分析中的一些数学假设不符[23],故而采用匀滑技术进行处理[24],由Ax得到ax。 为了检验种群存活状况是符合Deevey-Ⅱ型还是Deevey-Ⅲ型曲线,本文采用Hett和Loucks提出的数学模型对羽叶丁香种群存活曲线进行检验。即用指数方程Nx=N0e-bx和幂函数方程Nx=N0x-b分别描述Deevey-Ⅱ、Deevey-Ⅲ型存活曲线[25]。式中Nx、N0分别代表x龄级内存活数和种群形成初期的个体数,b为死亡率。 为了更好的揭示羽叶丁香种群的动态变化规律,本文引入生存分析中的4个函数,即生存率函数Sx、累计死亡率函数Fx、死亡密度函数fx和危险率函数λx,参考杨凤翔等[8]的公式进行计算。 1.2.5 种群数量动态的时间序列预测 本研究采用时间序列分析中的一次移动平均法[26]对羽叶丁香种群的年龄结构进行预测。 (12) 两个分布区羽叶丁香种群结构均呈现不规则金字塔型(图1),经计算得出,中卫种群个体主要集中在第Ⅲ—Ⅴ龄级,占比高达75.91%,贺兰山甘沟种群个体主要集中在第Ⅱ—Ⅴ龄级,占比达72.64%,说明两地区中幼龄级个体数很充足,但幼龄个体数占比较少,中卫种群第Ⅰ和Ⅱ龄级仅占总数的5.84%,贺兰山甘沟种群第Ⅰ和Ⅱ龄级也仅占16.42%,足以说明两地区严重缺乏幼龄个体。中老龄个体数总体均呈下降趋势,中卫种群个体数从第Ⅳ龄级后急剧减少,甘沟种群从Ⅲ龄级开始,除第Ⅷ龄级有少量增加外,总体也呈下降趋势。 图1 羽叶丁香种群径级结构 Fig.1 Size structure of Syringa pinnatifolia population根据基径(BD,Base Diameter)的分布状况,将基径划分为11个等级:Ⅰ级(BD<0.5cm)、Ⅱ级(0.5cm≤BD 表1 羽叶丁香种群动态变化指数 静态生命表分析结果显示,随着年龄增加,存活数(lx)逐渐减小,个体的期望寿命(ex)在Ⅰ、Ⅱ级后逐渐降低,这与其生物学特性一致。存活曲线是反映种群个体在各年龄级的存活状况曲线,其绘制方法有两种,一是以存活量的对数值lnlx为纵坐标,以年龄为横坐标作图;另一种是用存活数量对年龄作图,但年龄用平均寿命期望的百分离差来表示。本研究所作存活曲线是以径级相对龄级为横坐标,以lnlx为纵坐标获得的(图2)。按Deevey的划分,一般有3种基本类型,Ⅰ型是凸曲线,属于该型的种群绝大多数都是能活到该物种年龄,早期死亡率较低,但当活到一定生理年龄时,短期内几乎全部死亡;Ⅱ型是直线,也称对角线型,属于该型的种群各年龄的死亡率基本相同;Ⅲ型是凹曲线,早期死亡率高,一旦活到某一年龄,死亡率就较低。依据Hett和Loucks的数学模型得出的检验方程及参数见表2。 模型检验结果显示,两种模型的拟合结果在两地均达到了极显著水平,而指数模型的R2值均大于幂函数模型,且P值更小,说明两地区羽叶丁香种群的存活曲线都趋近于Deevey-Ⅱ型。 以径级相对龄级为横坐标,以各龄级的死亡率(qx)及损失度(kx)为纵坐标作死亡率和消失率曲线。两地种群的死亡率和消失率曲线变化趋势一致,均呈现出先增长再降低最后急剧增长的状态(图3),认为二者种群数量都具有前期减少,中期短暂增长,后期急剧减少的特点。对比可知,中卫种群的波动幅度更大,说明中卫种群更容易受到环境压力的影响。 表2 羽叶丁香种群存活曲线的检验模型 表3 羽叶丁香种群静态生命表 x:年龄级;Ax:x龄级内现有个体数;ax:匀滑后x龄级内的存活个体数;lx:x龄级开始时标准化存活个体数;dx:从x到x+1龄级间隔期内标准化死亡数;qx:从x到x+1龄级间隔期间死亡率;Lx:从x到x+1龄级间隔期间还存活的个体数;Tx:从x龄级到超过x龄级的个体总数;ex:进入x龄级个体的生命期望或平均期望寿命;Kx:消失率 图2 羽叶丁香种群存活曲线Fig.2 Survival curve of Syringa pinnatifolia population 图3 羽叶丁香种群死亡率和消失率曲线Fig.3 Mortality and vanish rate of Syringa pinnatifolia population 羽叶丁香种群生存分析结果显示,生存率和累计死亡率呈互补状态,即随着龄级的增加,羽叶丁香种群生存率逐渐下降,累计死亡率逐渐上升,中卫种群在第Ⅲ龄级达到平衡,贺兰山甘沟种群在第Ⅳ龄级达到平衡(图4),表明中卫种群比甘沟种群更早进入衰退期。贺兰山甘沟羽叶丁香种群死亡密度曲线趋势较为平缓,都保持在0.024以下,危险率整体上升趋势,两地种群均在Ⅹ龄级出现峰值(图5),这和死亡率曲线单调上升所得结论一致,后期个体随着逐渐进入生理衰退期,数量逐渐减少。比较可知,中卫香山种群危险率波动起伏更大,表明其受环境影响更明显。 图4 羽叶丁香种群生存率和累计死亡率曲线Fig.4 Survival rate and cumulative mortality rate of Syringa pinnatifolia population 图5 羽叶丁香种群死亡密度和危险率曲线Fig.5 Mortality density and hazard rate of Syringa pinnatifolia population 羽叶丁香种群数量动态变化的时间序列预测是以羽叶丁香种群各龄级株数为原始数据,按照一次移动平均法预测出各龄级在未来2、4、6、8和10个龄级时间后的个体数。分析结果显示,经历2、4、6个龄级时间,两地种群从Ⅴ龄级开始之后均有不同程度的增加,经过8和10个龄级时间,从Ⅷ龄级之后开始减少,趋于稳定,说明一旦过了一定的生理年龄,羽叶丁香种群就开始逐渐走向衰退(表4)。对于种群前中期,中卫种群在第Ⅱ—Ⅳ龄级经过2个龄级时间后个体数分别下降了42.86%、37.5%和23.91%,第Ⅳ龄级经过4个龄级时间后下降了45.71%;甘沟种群在第Ⅱ、Ⅲ龄级经过2个龄级时间后分别下降了43.33%和23.21%,Ⅳ龄级个体数在经历4个龄级时间后也下降了31.8%,这表明羽叶丁香种群严重缺乏幼龄个体。综合其整个发展趋势来看,羽叶丁香种群正常更新难以维持,未来仍然会不同程度地趋于衰退,甚至濒危。 表4 羽叶丁香种群动态变化的时间序列分析 Mt(1):经过2、4、6、8和10龄级时间后t龄级的种群大小;(1):移动平均法中的一次移动平均法标识 植物种群结构特征不仅能够体现种群内部个体的发展过程,而且也是对立地条件优劣及植物对环境适应性的反映[9],其径级结构能很好地反应种群动态变化[27]。本研究认为两地区羽叶丁香种群的年龄结构均呈不规则的金字塔型,动态指数结果显示中卫种群为增长型种群,而贺兰山甘沟种群为衰退型种群,且都对生存环境具有较高的敏感性,这可能和中卫香山地区当年生长季节的降雨量有关系。两地种群的中龄个体数量占种群总体数量的比例较高,老龄个体处于衰退状态,种群结构总体呈相对稳定型,有向衰退转化的趋势,这说明羽叶丁香种群虽然能暂时适应当地的生存环境,但由于Ⅰ、Ⅱ龄级个体数较少,幼龄个体无法补充中老龄个体的发展,种群整体的长期稳定难以维持,这与裸果木和水曲柳的种群特征较为相似[28- 29]。有研究表明羽叶丁香在野外主要以克隆繁殖方式为主[30],而这种繁殖方式产生的幼苗由于对环境变迁的适应能力弱且很容易遭到食草动物的采食和破坏,故在林下很难长到成林层以补充中老龄个体的发展,这在一定程度上限制了羽叶丁香种群的自然更新,形成该种群更新和发展的瓶颈。 生命表是判断植物种群发展趋势的重要指标之一,对植物种群的生命表和存活曲线的分析可以反映种群现实状况、展现植物种群与环境的竞争关系,对珍稀濒危植物的保护和利用具有重要意义[31]。对于濒危植物而言,其存活曲线的表现形式并不是单一的,在保护较好的天然群落中,一些濒危植物也属于稳定增长型种群,如长蕊木兰和猪血木等[32- 33];但在生境保护不力或所处生境本就恶劣的情况下,也经常出现幼苗更新不足所导致的衰退型种群,如新疆野苹果、半日花和珙桐等[34- 36]。本研究表明,两地区羽叶丁香种群存活曲线均趋向于Deevey-Ⅱ型,羽叶丁香种群Ⅷ龄级之前的死亡率相对稳定,但是Ⅷ龄级之后死亡率又急剧上升,说明老龄个体的发展遇到了瓶颈,这可能是因为一部分老龄个体的生理生长到了极限,对环境资源(如土壤、水分、光照等)的竞争能力下降,个体逐渐衰老,这与时间序列预测的结果一致。然而,时间序列分析也显示由于缺乏幼龄个体的补充,羽叶丁香种群未来的发展趋于衰退,这也是小范围、片段化分布的濒危植物天然种群必然经历的发展过程[31,37- 38]。生存分析表明,中卫香山种群较早进入衰退期,且受环境影响更明显,这主要是因为中卫香山地区的土壤层相对较薄,个体主要生长在坡的中部,贮藏水分的能力有限,而且较高的海拔与较低的温度导致了其生长季的缩短,较强的蒸发也会限制其个体的生长。综合以上分析,贺兰山甘沟地区更适宜羽叶丁香的长期生存,幼龄个体的缺乏是导致其濒危的主要原因。因此,人工抚育护林是阻止羽叶丁香种群衰退和进行种群恢复的关键。 植物种群数量的减少,一方面是由于幼苗的供给不足,一方面是已成活个体的死亡[39]。羽叶丁香种群幼龄个体严重不足,种群目前的稳定主要靠中龄个体维持,而且龄级越大的植株危险率越高,越容易死亡。分析两地区的气候因子可知,它们的年蒸发量是降雨量的7—10倍,且分布区多为山石区,海拔较高,无霜期较短,加之自然灾害影响及人类活动,其生境日益恶劣,生存条件加剧,故羽叶丁香的濒危现状已亟待解决。 针对羽叶丁香的濒危现状,本文认为可以从以下两个方面考虑该地区的种群保护和恢复策略: (1)从羽叶丁香个体自身因素考虑,其野外繁殖方式主要以克隆繁殖为主[30],野外调查时也发现,新萌生无性系幼苗经常分布在母株树下及其附近,种群扩大受影响。本文作者已对羽叶丁香种子的休眠机制与萌发特性作了详细研究,认为羽叶丁香在野外难以萌发主要是因为种子较轻,成熟后随风散布,很难进入土壤层,且其萌发季节的气候条件尤其是水分不充足[14]。而对于羽叶丁香生殖生态学其他方面的研究还很欠缺,因此,加大科研力度,尤其是高效繁育技术方面,是快速保护和恢复羽叶丁香种群的关键措施。具体来说,可以选择在降雨量较好的年份采集种子,人工散播于水分充足的生境内,并加以管护,提高实生幼苗的数量以扩大其种群分布范围;人工种植,引种栽培并加强推广,扩大其生态位等。 (2)从羽叶丁香种群所处生境考虑,其生存环境降雨少,蒸发快,雨水只在半阴坡的坡底会有少量存积,无法满足种群内所有个体的长期生长需求,结合Cui等人[40]的研究结论,羽叶丁香在迁地保护时首选夏季低温、阴凉和水分充足的环境,而中卫香山种群相对更急需此措施加以保护。羽叶丁香作为蒙医常用珍贵药材,以前也曾遭到人为的大量采挖,在多年的封山育林保护措施下,两地羽叶丁香种群才得以保存。因此,还应该在加大科学研究力度的同时继续加强对于羽叶丁香的保护和宣传力度,建立相应自然保护区和专类植物园,促使其种群恢复正常更新。 参考文献(References): [1] 张志祥, 刘鹏, 蔡妙珍, 康华靖, 廖承川, 刘春生, 楼中华. 九龙山珍稀濒危植物南方铁杉种群数量动态. 植物生态学报, 2008, 32(5): 1146- 1156. [2] 张婕, 上官铁梁, 段毅豪, 郭微, 刘卫华, 郭东罡. 灵空山辽东栎种群年龄结构与动态. 应用生态学报, 2014, 25(11): 3125- 3130. [3] 张文辉, 周建云, 李景侠. 秦岭濒危植物种群生态及保育技术研究. 咸阳: 西北农林科技大学出版社, 2015: 4- 6. [4] Holeksa J, Saniga M, Szwagrzyk J, Dziedzic T, Ferenc S, Wodka M. Altitudinal variability of stand structure and regeneration in the subalpine spruce forests of the Pol′ana biosphere reserve, Central Slovakia. European Journal of Forest Research, 2007, 126(2): 303- 313. [5] 刘普幸. 疏勒河中下游绿洲胡杨种群结构与动态研究. 自然资源学报, 2011, 26(3): 429- 439. [6] 张亮, 王孝安, 刘鹏, 王贤, 邓美皎, 朱云云. 陕西子午岭北桑寄生的种群生命表与生存分析. 植物生态学报, 2014, 38(7): 687- 693. [7] 韩路, 王家强, 王海珍, 宇振荣. 塔里木河上游胡杨种群结构与动态. 生态学报, 2014, 34(16): 4640- 4651. [8] 杨凤翔, 王顺庆, 徐海根, 李邦庆. 生存分析理论及其在研究生命表中的应用. 生态学报, 1991, 11(2): 153- 158. [9] 解婷婷, 苏培玺, 周紫鹃, 张海娜, 李善家. 荒漠绿洲过渡带沙拐枣种群结构及动态特征. 生态学报, 2014, 34(15): 4272- 4279. [10] 金山, 胡天华, 李志刚, 邢韶华, 崔国发. 贺兰山羽叶丁香分布区的植物物种多样性特性研究. 西部林业科学, 2008, 37(4): 40- 44. [11] 姜在民, 蔡靖, 崔宏安. 华榛、羽叶丁香种子形态构造特点的研究. 陕西林业科技, 1999, (3): 14- 16. [12] 中国科学院中国植物志编委会. 中国植物志(第八卷). 北京: 科学出版社, 1992, 61: 79- 79. [13] 布日额. 贺兰山濒危植物贺兰山丁香SyringapinnatifoliaHemsl. var.alashanensisMa et S. Q. Zhou的资源现状及药用民族植物学研究 // 第六届中国民族植物学学术研讨会暨第五届亚太民族植物学论坛论文集. 银川: 中国环境科学学会, 中国植物学会, 2012: 198- 202. [14] 和子森, 陈苏依勒, 程明, 温晋芳, 蔡靖, 姜在民. 濒危植物羽叶丁香种子休眠与萌发特性研究. 植物生理学报, 2016, 52(4): 560- 568. [15] 程明, 李厚华, 和子森, 姜在民, 蔡靖. 濒危植物羽叶丁香组织培养. 北方园艺, 2016, (12): 92- 96. [16] Ao W L J, Bao X H, Wu X L, Wang Q H. Lignans fromSyringapinnatifoliaHemsl. Var.alashanensis. Journal of Asian Natural Products Research, 2012, 14(4): 396- 400. [17] Ao W L J, Wang Q H, Qin S, Dan M, Ya S R T, Dai N Y T, Tu D R S H L. The structural elucidation and antimicrobial activities of two new sesquiterpenes fromSyringapinnatifoliaHemsl. Chinese Journal of Natural Medicines, 2012, 10(6): 477- 480. [18] 陈苏依勒, 苏国柱, 白睿峰, 尹旭, 曹愿, 屠鹏飞, 柴兴云. 羽叶丁香茎皮中环烯醚萜类成分. 中国中药杂志, 2016, 41(7): 1246- 1250. [19] Wang Q H, Bao J T, Han S R N, Ao W L J. A new 6, 9′-cyclolignan fromSyringapinnatifolia. Natural Product Research, 2016, 30(1): 85- 88. [20] 姚军朋, 陈莉, 崔洪霞, 姚拓. 丁香属植物光合机构的热响应特征研究. 草原与草坪, 2010, 30(5): 50- 55, 58- 58. [21] 何亚平, 费世民, 蒋俊明, 陈秀明, 张旭东, 何飞. 不同龄级划分方法对种群存活分析的影响——以水灾迹地油松和华山松种群生存分析为例. 植物生态学报, 2008, 32(2): 448- 455. [22] 陈晓德. 植物种群与群落结构动态量化分析方法研究. 生态学报, 1998, 18(2): 214- 217. [23] Wratten S D. Field and Laboratory Exercises in Ecology. London: Edward Arnold, 1980. [24] 江洪. 云杉种群生态学. 北京: 中国林业出版社, 1992. [25] Hett J M, Loucks O L. Age structure models of balsam fir and eastern hemlock. Journal of Ecology, 1976, 64(3): 1029- 1044. [26] 肖宜安, 何平, 李晓红, 邓洪平. 濒危植物长柄双花木自然种群数量动态. 植物生态学报, 2004, 28(2): 252- 257. [27] Arista M. The structure and dynamics of anAbiespinsapoforest in southern Spain. Forest Ecology and Management, 1995, 74(1/3): 81- 89. [28] 王立龙, 王亮, 张丽芳, 刘玉洋, 徐世健. 不同生境下濒危植物裸果木种群结构及动态特征. 植物生态学报, 2015, 39(10): 980- 989. [29] 易雪梅, 张悦, 王远遐, 姬兰柱, 吴培莉. 长白山水曲柳种群动态. 生态学报, 2015, 35(1): 91- 97. [30] 杨亚珺, 李吉宁, 巩檑, 周坚. 贺兰山丁香自然居群克隆生长格局及遗传多样性的ISSR分析. 植物科学学报, 2013, 31(1): 85- 92. [31] 杨小林, 王秋菊, 兰小中, 李春燕. 濒危植物大花黄牡丹(Paeonialudlowii)种群数量动态. 生态学报, 2007, 27(3): 1242- 1247. [32] 袁春明, 孟广涛, 方向京, 柴勇, 李贵祥, 蔺汝涛, 王骞, 李品荣. 珍稀濒危植物长蕊木兰种群的年龄结构与空间分布. 生态学报, 2012, 32(12): 3866- 3872. [33] 申仕康, 马海英, 王跃华, 王博轶, 申国柱. 濒危植物猪血木(EuryodendronexcelsumH. T. Chang)自然种群结构及动态. 生态学报, 2008, 28(5): 2404- 2412. [34] 田润炜, 蔡新斌, 刘丽燕, 江晓珩, 布早拉木·吐尔逊, 买尔燕古丽·阿不都热合曼. 新疆野苹果种群年龄结构特征与动态分析. 西北植物学报, 2016, 36(4): 811- 817. [35] 李清河, 高婷婷, 刘建锋, 辛智鸣, 徐军. 荒漠珍稀灌木半日花种群的年龄结构与生命表分析. 植物研究, 2009, 29(2): 176- 181. [36] 刘海洋, 金晓玲, 沈守云, 张日清. 湖南珍稀濒危植物——珙桐种群数量动态. 生态学报, 2012, 32(24): 7738- 7746. [37] 谢伟东, 陈建华, 赖家业, 石海明, 林少芳, 刘宝, 黎向东. 珍稀濒危植物蒜头果种群生命表分析. 中南林业科技大学学报, 2009, 29(2): 73- 76. [38] 刘海燕, 杨乃坤, 李媛媛, 洪江, 邹天才. 稀有濒危植物长柱红山茶种群特征及数量动态研究. 植物科学学报, 2016, 34(1): 89- 98. [39] Yoshida T, Kamitani T. Growth of a shade-intolerant tree species,Phellodendronamurense, as a component of a mixed-species coppice forest of central Japan. Forest Ecology and Management, 1999, 113(1): 57- 65. [40] Cui H X, Jiang G M, Niu S L, Li Y G, Jiang C D, Liu M Z, Gao L M. Gas exchanges of an endangered speciesSyringapinnatifoliaand a widespread congenerS.oblata. Photosynthetica, 2004, 42(4): 529- 534.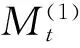
2 结果和分析
2.1 羽叶丁香种群分布现状及结构特征
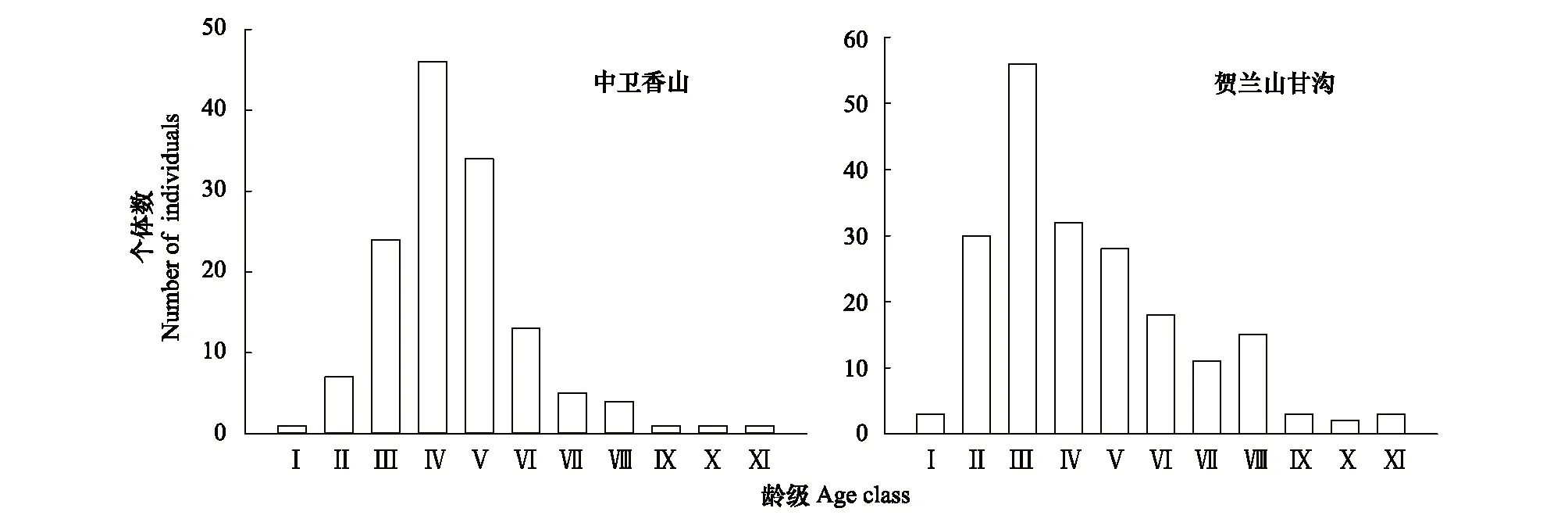
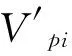
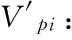
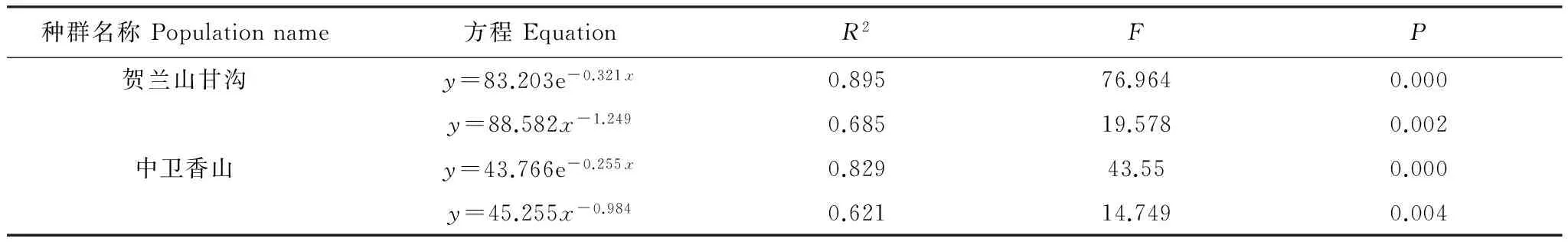
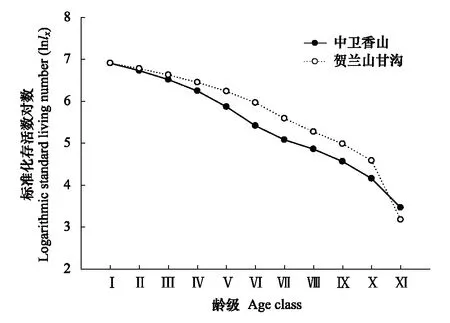
2.2 羽叶丁香种群静态生命表及存活曲线
2.3 羽叶丁香种群的生存分析
2.4 羽叶丁香种群数量的时间序列分析
3 讨论
3.1 羽叶丁香种群的结构特征
3.2 羽叶丁香种群的动态趋势
3.3 羽叶丁香种群的保护和恢复对策
