不同种源桤木嫁接幼苗的生长及光合特性
2018-05-02白文玉铁烈华冯茂松汪亚琳李文兵
白文玉,铁烈华,冯茂松,汪亚琳,李文兵,谭 飞
(四川农业大学林学院/长江上游林业生态工程四川省重点实验室,成都 611130)
桤木(Alnus nepalensis)又称四川桤木,桦木科桤木属落叶高大乔木,喜温暖、湿润、光照良好的环境,树干通直,为西南地区优良速生用材树种[1]。桤木根部着生根瘤菌,对土壤改良效果明显,在传统农业生态系统中扮演着重要的角色[2]。桤木良好的适应性和速生丰产性,让其成为理想的用材林和生态防护林树种,已被列入“森林资源发展和保护项目(FRDPP)”,是国家二级重点保护野生植物[3-4]。近年来,随着桤木造林面积的不断扩大,国内外学者对桤木属树种的丰产栽培技术[5]、光合生理[6]和生态恢复作用[7]等做了许多研究。
光合作用将H2O和CO2合成有机物,为植物生长发育提供物质和能量基础,光合速率(Pn)受光合色素含量、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)等因素的影响,提高光合性能是植物增产、增收的重要途径[8-9]。不同种源桤木长期处于不同的生境中,由于遗传变异和自然选择的结果,其生长情况、光合生理特性和形态结构等将可能发生明显变化,这为桤木良种育苗提供了繁育材料。
为研究不同种源桤木嫁接幼苗的生长及光合特性,选育有栽培前景的桤木良种幼苗,本试验在四川省平昌县、宣汉县、恩阳区和金堂县共选择桤木优树150株,采集健壮的穗条与实生桤木幼苗进行嫁接,分别测定4个种源桤木无性系嫁接幼苗的生长情况和光合生理指标,分析并比较不同种源桤木嫁接幼苗生长情况和光合生理特性,为四川省桤木良种育苗基地的建立提供基础理论数据。
1 材料和方法
1.1 试验区概况
巴中市平昌县(31°16'~31°52'N,106°15'~107°34'E)位于四川省东北部,属于中亚热带湿润季风气候,年降雨量为1 213 mm。四季分明,气候温和,平均气温16.8℃,年平均日照时数1 239.3 h,无霜期297 d。境内植被丰富,主要乔木树种有桤木、柏木(Cupressus funebris)、马尾松(Pinus massoniana)等。本次试验地位于平昌县张公乡的四川省桤木良种育苗基地,基地朝向东南方,坡度约10°,配套有灌溉设备,光照充足,土壤为紫色壤土,呈弱酸性,土层深约40 cm。
1.2 试验设计
1.2.1 优树选择与穗条采集
经过调查,四川省平昌县、宣汉县、恩阳区和金堂县均为桤木适生区,区域内桤木生长良好,病虫害较少,分布相对集中成片,4个种源的主要生态因子如表1所示。2014年12月底,选择郁闭度大于0.6,林相整齐的15~20 a生桤木纯林,采用优势木对比法筛选出自然整枝良好、树干通直、无病虫害的桤木优树。共选择优树150株(依次编号为1~150号),每株采集穗条约200枝,采集穗条时选择树冠中上部生长健壮、叶芽饱满、粗度3~5 mm,长度30~50 cm的1 a生枝条。将采集好的穗条打捆,用湿报纸包裹,放入编号,做好登记,并尽快带回实验室,保存于4℃冰柜中待用。
1.2.2 嫁接育苗
2014年11月,将苗圃地划分为若干个1.2 m×6 m的小区,选择生长健壮、长势基本一致、无病虫害的当年生桤木实生营养袋苗,按株行距20 cm×30 cm进行栽植,作为砧木,每个小区约栽植140株。2015年1月初,采用随机区组设计方法,采集的桤木穗条嫁接于长势基本一致、无病虫害的砧木上,同株优树上采集的穗条分别嫁接于3个小区,每小区内约嫁接100株,共嫁接约300株。挂上对应的标号牌,做好登记,搭建小拱棚,及时浇水、除草,并完善田间管理日志。
1.3 指标测定
本试验共采集平昌、宣汉、恩阳和金堂4个种源共150株优树,其中150株优树无性系全部参与生长特性、光合色素和光合速率的测定,平昌、宣汉、恩阳和金堂种源分别随机选择30、10、5和5株优树无性系用于光响应曲线的测定。

表1 4个种源基本情况表Table 1 Basic situation of different provenances
1.3.1 生长指标测定
2015年7月中旬,调查小区内嫁接幼苗的苗高、地径、成活率等。
1.3.2 光合生理特征测定
2015年7月中旬,根据生长指标的测定结果,每小区内筛选出5株长势具有代表性的嫁接幼苗作为待测植株,测定其光合生理指标。于无风的晴天,选取待测植株中上部3片完全成熟且健康的叶片,用便携式光合作用仪 Li 6400(Li-Cor Inc.,USA)分别测定其光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci),测定时控制CO2浓度为400 μmolCO2/mol、温度为 25 ℃、光照强度为 1 500 μmol/(m2·s)。同时,在各待测植株中上部选取1片相同叶位的成熟叶片,用于光响应曲线数据的测定,测定光源为便携式光合作用仪Li 6400(Li-Cor Inc.,USA),自带红蓝光源,光强梯度设定为 0、25、50、75、100、200、400、600、800、1 000、1 200、1600 和2 000 μmol/(m2·s)。测定时设定温度为25℃,空气湿度为70%。
1.3.3 光合色素含量测定
将测定完光合参数的叶片带回实验室,采用丙酮-乙醇混合液提取法提取光合色素,并用723PC型分光光度计测定光合色素含量[10]。
1.4 数据分析与处理
采用非直线双曲线模型拟合光响应曲线,模型表达式如下[11],同时计算光补偿点(LCP)、光饱和点(LSP)、表观量子效率(AQY)和暗呼吸速率(Rd)。
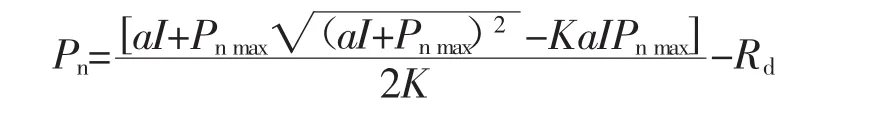
式中,a表示表观量子速率(AQY);Pnmax为最大光合速率;I为光合光合有效辐射(PAR);K为光响应曲线的曲角;Rd为暗呼吸速率。将光强为200 μmol/(m2·s)以下的Pn-PAR响应曲线进行直线回归,其斜率即为表观量子效率,该方程与最大光合速率的交点即为光饱和点(LSP),与x轴的交点即为光补偿点(LCP)。
用Excel 2007和SPSS 19.0统计分析软件(SPSS Inc,USA)对数据进行整理、单因素方差分析(one-way ANOVA)和相关性分析等,显著性分析设定a=0.05。
2 结果与分析
2.1 不同种源桤木幼苗生长特性
由表2可知,金堂种源桤木嫁接幼苗高度最高,达137.1 cm,与恩阳幼苗高度差异不显著(P>0.05),与平昌和宣汉的差异显著(P<0.05),且其基径最大,为 12.0 mm,显著高于其余种源(P<0.05)。平昌、宣汉、恩阳种源桤木嫁接幼苗的苗高和基径差异均不显著(P>0.05)。4个种源幼苗嫁接成活率均未达到50%,其中平昌幼苗嫁接成活率最高,为48.4%,显著高于其余种源(P<0.05),宣汉、恩阳、金堂种源之间嫁接成活率差异不显著(P>0.05),其成活率分别为26.2%、24.7%、30.0%。

表2 不同种源桤木幼苗生长特性Table 2 The growth characteristics of different provenances for the seedlings of Alnus
2.2 不同种源桤木幼苗光合色素含量
恩阳桤木嫁接幼苗的叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量均最高(表3),分别为2.45、0.61、3.06和0.64 mg/g,且均与金堂的差异不显著(P>0.05),与平昌、宣汉的差异显著(P<0.05)。平昌、宣汉、金堂三者间的叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量差异均不显著(P>0.05),且各光合色素含量均表现为金堂>平昌>宣汉。宣汉嫁接幼苗的叶绿素a/b最大,为4.73,显著高于其余种源(P<0.05)。4个种源间类胡萝卜素/总叶绿素的差异不显著(P>0.05)。

表3 不同种源桤木幼苗光合色素含量Table 3 The photosynthetic pigment content of different provenances for the seedlings of Alnus
2.3 不同种源桤木幼苗光合生理指标
2.3.1 不同种源桤木幼苗的光响应曲线
由图1可知,在有效光辐射小于200 μmol/(m2·s)的弱光环境下,各种源桤木嫁接幼苗的光合速率(Pn)迅速上升;当有效光辐射超过500 μmol/(m2·s)时,随光照强度的增加,桤木嫁接幼苗的光合作用增长速度明显放缓;当有效光辐射超过1 500 μmol/(m2·s)后,其光响应曲线趋近平稳。

图1 不同种源桤木幼苗光合作用的光响应曲线Figure 1 The light-photosynthetic response curves of different provenances for the seedlings of Alnus
由表4可知,采用非直线双曲线模型拟合光响应曲线,其拟合优度(R2)均大于0.924。4个种源的表观量子效率(AQY)大小在 0.030~0.044 mol/mol之间,这与自然条件下一般植物的表观量子效率(0.030~0.050 mol/mol)一致,其中恩阳幼苗表观量子效率(AQY)最大,为0.044 mol/mol,且与宣汉、金堂的差异不显著(P>0.05),与平昌的差异显著(P<0.05);宣汉、恩阳、金堂三者间表观量子效率(AQY)差异均不显著(P>0.05)。恩阳幼苗的暗呼吸速率(Rd)最快,为2.053 μmol CO2/(m2·s),且与宣汉的差异不显著(P>0.05),与平昌和金堂的差异均显著(P<0.05)。此外,恩阳种源幼苗的最大光合速率(Pnmax)最大,为19.1 μmol CO2/(m2·s),与金堂的差异不显著(P>0.05),与平昌和宣汉的差异显著(P<0.05)。金堂幼苗的光补偿点(LCP)最低,为29.17 μmol/(m2·s),与其他种源的差异显著(P<0.05)。金堂种源幼苗的光饱和点(LSP)最大,且与平昌和恩阳的差异均不显著(P>0.05),与宣汉的差异显著(P<0.05)。
2.3.2 不同种源桤木幼苗光合特征参数
由表5可知,恩阳种源幼苗的光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)均最大。其光合速率(Pn)与金堂和宣汉的差异不显著(P>0.05),与平昌的差异显著(P<0.05);气孔导度(Gs)与金堂的差异不显著(P>0.05),与平昌和宣汉的差异显著(P<0.05);蒸腾速率(Tr)显著高于其余种源(P<0.05)。4个种源幼苗的胞间CO2浓度(Ci)差异不显著(P>0.05)。
2.3.3 桤木幼苗叶片气体交换参数间的相关性分析

表4 不同种源桤木幼苗光响应曲线参数Table 4 The light response curve of different provenances for the seedlings of Alnus
由表6可知,光合速率(Pn)与气孔导度(Gs)和蒸腾速率(Tr)之间,以及气孔导度(Gs)与胞间CO2浓度(Ci)和蒸腾速率(Tr)之间,胞间CO2浓度(Ci)与蒸腾速率(Tr)之间均存在极显著相关关系(P<0.01)。光合速率(Pn)与胞间CO2浓度(Ci)不相关(P>0.05)。

表5 不同种源桤木幼苗叶片的气体交换参数Table 5 Gas exchange parameters for the seedlings of Alnus

表6 桤木幼苗叶片气体交换参数之间的相关性Table 6 The correlation among gas exchange parameters for the seedlings of Alnus
3 讨论与结论
3.1 不同种源桤木嫁接幼苗的生长特性
本研究表明,4个种源的桤木幼苗中,平昌种源的嫁接成活率最高,接近50%,其他种源的嫁接成活率均在30%以下。出现这种现象可能是因为平昌本地采集的穗条运输时间短,嫁接及时,穗条水分和营养物质损失较少,活力较高。同时,嫁接时采用平昌桤木实生苗为砧木,砧木与接穗的适应性和亲和度可能较高。故从远距离采集桤木穗条时,应做好保鲜措施,并尽快嫁接,以提高嫁接成活率。本研究还表明,4个种源桤木幼苗嫁接成活率均未超过50%。这可能是因为,一方面桤木幼苗的韧皮部较薄,嫁接后不易成活;另一方面关于桤木幼苗嫁接技术的研究比较少,嫁接技术不够成熟,成活率较低;再者也可能是因为桤木幼苗嫁接后田间管理不足,导致嫁接成活率不高。因此,随采随嫁、提高桤木嫁接技术、增强桤木嫁接苗的田间管理,是提高桤木嫁接成活率的重要措施。
较强的光合利用能力能够促进植物幼苗的生长。本研究表明,金堂和恩阳种源桤木嫁接幼苗的苗高和基径较大。这可能是因为,金堂和恩阳种源桤木嫁接幼苗的光合利用作用较强,促进了幼苗的生长。这与A.C.Moreira等[17]对兰花光合生理研究结果一致。可能是因为,首先,恩阳种源桤木嫁接幼苗光合色素含量较高,在白天,气孔导度较大、蒸腾速率较高,CO2进出胞间的速度较快,光合速率较高,桤木嫁接幼苗储备大量的有机物,但在弱光的傍晚和晚上,虽表观量子效率稍高,但暗呼吸速率较快,光补偿点较高,大量消耗白天储备的有机物,影响其生长速度。其次,金堂桤木嫁接幼苗与恩阳幼苗的光和色素含量相当(P>0.05),在强光照的白天金堂种源桤木嫁接幼苗具有与恩阳嫁接幼苗相似(P>0.05)的最大光合速率和光合速率,光饱和点和蒸腾速率也较高,可以迅速带走叶片周围的热量,减轻高温胁迫对幼苗的危害,气孔开度较大,CO2进出胞间的速度较快,有机物大量积累。同时,在弱光的环境下,金堂种源桤木嫁接幼苗暗呼吸速率和光补偿点显著低于恩阳种源(P<0.05),不至于消耗过多的养分。因此,金堂种源桤木嫁接幼苗的苗高和基径最大,长势最佳。可见,金堂种源能较好地适应川东北的气候环境,光合速率较高,呼吸速率较低,有机物质储存较快。本研究还表明,平昌和宣汉种源桤木嫁接幼苗生长较慢。这可能是因为,平昌和宣汉种源的光和色素含量相对较低(P<0.05),在强光的白天,其蒸腾速率和气孔导度均显著(P<0.05)小于恩阳和金堂种源,最大光合速率和光合速率较低,影响有机物质的合成与积累,其嫁接幼苗生长较慢。
3.2 不同种源桤木嫁接幼苗的光合特性
叶绿素是光合作用过程中最重要的光合色素,在光合作用中起着接受和转换光能的作用,在一定情况下,其含量的高低直接决定了光合速率快慢,影响生态系统物质和能量的输入[18]。本研究表明,金堂和恩阳种源桤木嫁接幼苗的表观量子效率、光饱和点和光合速率较高,其嫁接幼苗的光合利用能力较强。这与应小芳等[19]对大豆叶片光合特性的研究结果相似。这可能是因为,在本次研究中4个种源桤木嫁接幼苗光合色素含量和光合速率存在显著性(P<0.05)差异,恩阳和金堂桤木嫁接幼苗的光合色素含量较高,其光合作用的光反应阶段较快,桤木嫁接幼苗对光能的吸收、转换与利用加强,从而提高桤木嫁接幼苗的光合速率。可见,光合作用的强度受光合色素含量影响。
气孔是植物叶片中最重要的气体运输通道,直接控制CO2向叶片的传导速率和蒸腾效率[14]。本研究表明,4个种源桤木嫁接幼苗的光合速率、气孔导度和蒸腾速率存在显著性(P<0.05)差异,相关性分析发现光合速率与气孔导度和蒸腾速率之间存在极显著相关关系(P<0.01)。这可能是因为,桤木嫁接幼苗光合速率受气孔导度和蒸腾速率的影响,恩阳和金堂桤木嫁接幼苗的气孔导度较大、蒸腾速率较高,光合作用固定CO2的能力较强,光合速率较快。可见,光合作用与气孔导度之间存在一定的调节机制。这与S.C.Wong等[20]的研究结果相似。
综上所述,4个种源桤木嫁接幼苗生长及光合特性之间存在显著(P<0.05)差异,恩阳和金堂种源桤木嫁接幼苗叶片的光合色素含量较高,光合速率较强,生长较快。其中,恩阳嫁接幼苗在弱光下呼吸消耗大,光补偿点和光饱和点较高,适合栽植于光照充足的地区;金堂嫁接幼苗对强光和弱光的利用能力较强,长势最佳,适合栽植的范围更广。另外,在桤木良种育苗时应注意嫁接穗条的随采随嫁、改善嫁接技术、增强田间管理能力,保证桤木嫁接成活率。
参考文献:
[1]杨志成.优良阔叶树种——桤木的分布、生长与利用[J].林业科学研究,1991,4(6):643-648.
[2]李苏梅,龙春林,刀志灵,等.传统农业生态系统中桤木改良土壤效应研究综述[J].植物生态学报,2006,30(5):878-886.
[3]王军辉,顾万春,李斌,等.桤木优良种源家系的选择研究——生长的适应性和遗传稳定性分析[J].林业科学,2000,36(3):59-66.
[4]王斌,张军鹏,施彬,等.桤木属植物繁育技术研究进展[J].山东林业科技,2016,46(6):106-110.
[5]王勇军,肖前刚,陈斌,等.四川桤木优树选择与育苗技术[J].四川林业科技,2013,34(6):60-62.
[6]TOBITA H,UEMURA A,KITAO M,et al.Effects of elevated atmospheric carbon dioxide,soil nutrients and water conditions on photosynthetic and growth responses of Alnus hirsute[J].Functional Plant Biology,2011,38(38):702-710.
[7]COPOLOVICI L,KANNASTE A,REMMEL T,et al.Volatile organic compound emissions from Alnus glutinosa under interacting drought and herbivory stresses[J].Environmental&Experimental Botany,2014,201(3):887-896.
[8]ROZENDAAL D M A,HURTADO V H,POORTER L.Plasticity inleaf traits of 38 tropical tree species in response to light;relationships with light demand and adult stature[J].Functional Ecology,2006,20:207-216.
[9]CAKMAK I,MARSCHNER H.Magnesium deficiency and high light intensity enhance activities of superoxide dismutase,ascorbate peroxidase,and glutathione reductase in bean leaves[J].Plant Physiology,1992,98:1222-1227.
[10]杨敏文.快速测定植物叶片叶绿素含量方法的探讨[J].光谱实验室,2007,19(4):478-481.
[11]吴秀华,胡庭兴,杨万勤,等.巨桉凋落叶分解对菊苣生长及光合特性的影响[J].应用生态学报,2012,23(1):1-8.
[12]张璐颖,文笑,林勇明,等.盐胁迫对台湾桤木幼苗光合作用和荧光特性的影响[J].福建林学院学报,2013,33(3):193-199.
[13]刘俊祥,孙振元,勾萍,等.镉胁迫下多年生黑麦草的光合生理响应[J].草业学报,2012,21(3):191-197.
[14]李羿桥,李西,胡庭兴.巨桉凋落叶分解对假俭草生长及光和特性的影响[J].草业学报,2013,22(3):169-176.
[15]刘柿良,马东明,潘远智,等.不同光环境对桤木幼苗生长和光合特性的影响[J].应用生态学报,2013,24(2):351-358.
[16]叶子飘,于强.光合作用光响应模型的比较[J].植物生态学报,2007,32(6):1356-1361.
[17]MOREIRA A S,FILHOA J P,ZOTZ G,et al.Anatomy and photosynthetic parameters of roots and leaves of two shade-adapted orchids,Dichaea cogniauxiana Shltr.and Epidendrum secundum Jacq[J].Flora-Morphology,Distribution,Functional Ecology of Plants,2009,204:604-611.
[18]张力文,钟国成,张利,等.3种鼠尾草属植物光合作用--光响应特性研究[J].草业学报,2012,21(2):70-76.
[19]应小芳,刘鹏.铝胁迫对大豆叶片光合特性的影响[J].应用生态学报,2005,16(1):166-170.
[20]WONG S C,COWAN I R,FARQUHAR G R.Stomatal conductancecorrelateswithphotosyntheticcapacity[J].Nature,1994,282:424-426.
