酸乳发酵中优良共生酵母菌的筛选
2018-04-26袁凤霞曹晓虹刘建利王军节李靖宇李学斌覃璐琪邓鹏程王晓丹
袁凤霞,田 莎,曹晓虹,刘建利,王军节,张 琇,李靖宇,宋 峰,李学斌,覃璐琪,邓鹏程,王晓丹
(北方民族大学生物科学与工程学院;发酵酿造工程生物技术国家民委重点实验室;宁夏特殊生境微生物资源开发与利用重点实验室,宁夏银川 750021)
乳制品由于具有较高的营养价值和保健作用,是人们生活必备食品。研究发现,发酵乳制品制作过程中的微生物与乳制品风味、口感、质地等品质密切相关[1]。乳酸菌最早被发现是乳制品加工过程中发挥作用的主要微生物[2-3]。但自从在开菲尔(Kefir)[4-7]、酸马奶(Koumiss)[8-13]中酵母菌的重要作用被发现后,在越来越多的乳制品如:发酵驼乳酪乳清[14]、原料乳和奶酪[15-16]、奶渣[17]、曲拉和乳饼[18-19]、发酵乳[20]等中也有大量酵母菌被分离出。随着酵母菌抑菌、降胆固醇、抗氧化、活化集体免疫系统等生理功效的发现,传统乳制品中酵母菌越来越被关注。酸乳作为乳制品中销售增长最快的产品,新型酸乳研发是各大乳企研发热点,然而利用特殊酵母菌开发新型酸乳发酵菌剂却鲜有开展。
西北、西南、东北民族地区的少数民族食用各种发酵食品有着悠久的历史,千百年来沿袭古老而传统的方法制作酸乳、奶酪、奶豆腐、奶酒、曲拉、乳扇、乳饼等乳酸菌发酵食品,方法简单、风味独特、组织细腻、口感纯正,在当地自然温度下存放较长时间还能保证其口感和状态。在这些传统乳酸菌发酵食品中应该含有丰富、独特的优良微生物资源包括酵母菌资源,这些共生酵母资源有待挖掘。
本实验以前期从新疆、西藏和青海等地收集的传统自制乳制品中分离的27株酵母菌为基础,将酵母菌株分别和保加利亚乳杆菌(Lactobacillusdelbrueckiisubsp. bulgaricus)共同发酵制备酸乳,以保加利亚乳杆菌单菌发酵作为对照,通过比较凝乳时间、感官特性、质地特性、后酸度、粘度、持水力、产香能力等特征,筛选酸乳发酵中与保加利亚乳杆菌优良共生的酵母菌株,为开发新型酸乳提供参考。
1 材料与方法
1.1 材料与仪器
酵母菌 青海西宁牦牛酸乳分离的“QX1”-“QX15”,青海大通县酸乳中分离的“QD1”~“QD3”,山东酸乳中分离的“SD1”、“SD2”,从西藏拉萨、那曲、山南酸牦牛乳中分离的“XS1”、“XN1”~“XN6”;保加利亚乳杆菌(Lactobacillusdelbrueckiisubsp. bulgaricus) 菌株MUNLd1由本实验室保存。
胰蛋白胨、酵母提取物 英国Oxoid公司;牛肉浸粉 青岛海博生物公司;乙醛标准品 北京中科质检生物技术有限公司;双乙酰标准品 Sigma公司;酵母基因组提取试剂盒 康为世纪生物科技有限公司;2×EasyTaq PCR SuperMix(+dye) 北京全式金生物技术有限公司;柠檬酸二铵、柠檬酸铵、蔗糖、碘、三氯乙酸、邻苯二胺、盐酸、亚硫酸氢钠、酚酞、氢氧化钠、酒精、氯化钙、氯化钠、磷酸二氢钠、磷酸氢二钠、无水乙醇、氢氧化钾 化学纯,天津市大茂化学试剂厂;纯牛奶 伊利乳业有限责任公司。
1%邻苯二胺 准确称取邻苯二胺0.25 g,溶于4 mol/L盐酸中,置于25 mL容量瓶内,用4 mol/L盐酸补足至刻度,贮于棕色瓶中,临用时配制。
MRS培养基(1L) 胰蛋白胨10 g、酵母提取物5 g、柠檬酸二铵2 g、牛肉浸粉10 g、无水乙酸钠5 g、葡萄糖20 g、吐温80 1 mL、硫酸镁0.58 g、磷酸氢二钾2.6 g、硫酸锰0.19 g,调节pH为6.2~6.4之间。
YPD培养基 1%酵母膏、2%蛋白胨、2%葡萄糖。
26S rDNA D1/D2区引物 NL1:5′-GCATATCG GTAAGCGGAGGAAAA-3′;NL4:GGTCCGTGTTTCAA GACGG-3′;ITS1区引物 ITS1:5′-TCCGTAGGT GAACCTGCGC-3′;ITS2 5′-GCTGCGTTCTTCATCG ATGC-3′(生工生物工程(上海)股份有限公司合成)。
TMS-PRO型质构仪 美国FTC公司;NDJ-8S型粘度计 上海庚庚仪器设备公司;S1000型快速梯度PCR仪 美国Bio-Rad公司;1-14型台式高速冷冻离心机 Sigma公司。
1.2 实验方法
1.2.1 酵母菌株和保加利亚乳杆菌株的活化 取保藏的27株酵母菌和保加利亚乳杆菌株分别接入YPD、MRS液体培养基中,37 ℃,150 r/min摇床培养24 h,6000 r/min,4 ℃离心10 min,弃去上层培养液,加入无菌生理盐水调整浓度大于1×1010cfu/mL,制成菌悬液备用。
1.2.2 酸乳的制备 参考刘敏敏[8]、余兰[14]、李先胜[19]等方法,30 mL含蔗糖的牛奶(15%),高压灭菌锅95 ℃灭菌5 min,待冷却后,接入3 mL保加利亚乳杆菌悬液和1 mL酵母菌菌悬液,40 ℃培养,记录凝乳时间,凝乳后4 ℃冷藏24 h后熟,进行后续混菌酸乳特性评价;以接入3 mL保加利亚乳杆菌悬液和1 mL生理盐水的样品作为单菌发酵对照。
1.2.3 酸乳特性评价
1.2.3.1 酸乳感官评价 参考生庆海等[21]方法,制定感官评价表,将后熟24 h的酸乳取出后,随机选15名北方民族大学生物科学与工程学院2014级食品科学与工程专业学生依据感官评价标准对其气味、质地、口感三个指标进行感官评定,计算平均值。

表1 酸乳感官评价打分表Table 1 Sensory evaluation scale of yoghourt
1.2.3.2 酸乳持水力测定 参考张兰威[22]的方法,准确称取后熟24 h的酸乳15 g,4500 r/min离心10 min,取上清液,称取质量。持水力(%)=(样品质量-上清液质量)/样品质量×100。
1.2.3.3 酸乳质构特性测定 参考徐鑫[23]、赫君菲[24]和杨莹莹[25]等方法,测试探头型号为TMS-75 mm圆盘挤压探头P/75。测试条件如下:测前速率为30 mm/min;测后速率与测前速率一致;两次压缩之间的停留间隔:0 s;采样速度:10 Hz;最小触发力:0.3 N。
1.2.3.4 酸乳滴定酸度测定 按照GB 5009.239-2016[26],加5 g 4 ℃贮存1、3、7 d酸乳、40 mL冷却煮沸水和5滴已经配好的酚酞酒精溶液(浓度为5 g/L),于250 mL三角瓶充分摇匀,用NaOH标准溶液(浓度为0.1000 mol/L)滴定至微红色,在30 s内颜色不消失即为滴定终点,记录消耗的NaOH标准滴定溶液毫升数。将所得结果代入公式中计算滴定酸度。
X(°T)=c×V×100/m×0.1,X-发酵乳样品的酸度,单位为度(°T),吉尔涅尔度°T,100 g发酵乳消耗0.1 mol/L NaOH标准溶液的体积;c-NaOH标准溶液的摩尔浓度,单位为摩尔每升(mol/L);V-滴定时消耗NaOH标准溶液体积,单位为毫升(mL);m-发酵乳样品的质量,单位为克(g);0.1-酸度理论定义NaOH的摩尔浓度,单位为摩尔每升(mol/L)。
1.2.3.5 酸乳中双乙酰含量测定 邻苯二胺法[27]。标准曲线的绘制:取12支编号试管分成2排放置,并分别加0.0、2.0、4.0、6.0、8.0、10.0 mL标准溶液各2支,分别用蒸馏水补足10 mL,取5.0 mL上述双乙酰标准溶液加入5 mL 16%三氯乙酸溶液,得到双乙酰标准溶液的质量浓度分别为0.0、3.0、6.0、9.0、12.0、15.0 μg/mL,在335 nm波长下用石英比色皿测定吸光度值。以双乙酰质量分数为横坐标,吸光度值为纵坐标绘制标准曲线。样品中双乙酰含量的测定:取后熟24 h的酸乳10 mL加三氯乙酸(浓度为16%)溶液10 mL,静置10 min后4500 r/min,4 ℃离心10 min,过滤后取上清液10 mL,加0.5 mL质量分数为1%的邻苯二胺摇匀,放置黑暗处静置30 min。待反应完全后加入浓度为4.0 mol/L的盐酸,波长335 nm石英比色皿测定吸光度值,代入标曲计算样品中含量。
1.2.3.6 酸乳中乙醛含量测定 滴定法[28]。取5 mL样品加入5.00 mL三氯乙酸(质量分数为16%),充分摇匀,12000 r/min,4 ℃离心10 min,在250 mL锥形瓶中加入5 mL上清液2.00 mL 1% NaHSO3的摇匀,室温放置1 h后,加入1 mL 1%淀粉溶液,用0.01 mol/L碘液滴至淡蓝色,且在30 s不褪色即为滴定终点。再加入20 mL 1 mol/L NaHCO3,振荡到蓝色退去,再用0.01 mol/L碘标准溶液滴定至淡蓝色,30 s不褪去,记录消耗标准碘液的体积,按如下公式计算乙醛的含量。
乙醛(mg/L)=(V1-V2)×c×0.022/10
式中:V2为空白滴定消耗碘液标准的体积,mL;V1为样品滴定消耗碘液标准的体积,mL;c为1/2碘液标准溶液的浓度,mol/L;10为乙醛样品称样量,mL;0.022为乙醛化学反应基本单位,g。
1.2.4 数据处理与分析 每个实验重复3次,采用SPSS11.0软件ANOVA对实验数据进行方差分析,用LSD进行多重比较,图表在Excel中生成。
1.2.5 优良共生酵母菌株分子鉴定 参照酵母菌基因组DNA提取试剂盒说明书;PCR扩增26S rDNAD1/D2区、ITS1区反应体系为(50 μL):10×PCR Mix 25 μL、10 μmol/L的上下游引物各1 μL、模板DNA 1 μL、ddH2O 22 μL。PCR反应程序为参考Kurtzman等[29]:取3 μL PCR产物进行1.5%琼脂糖电泳检测,剩余PCR扩增产物由南京金斯瑞生物公司进行测序。将测序序列在NCBI数据库中比对鉴定。参考序列选自Kurtzman等[29],以SchizosaccharomycespombeD1D2区序列(U40085.1)和Hanseniasporaoccidentalis的ITS序列(AY046203.1)为outgroup,将所有序列输入在Mega Ver6.0软件[30]中打开,alignment文件,手动修正掐头去尾,输出fasta格式;将修正后的fasta文件用DAMBE Ver 5.3.4软件[31]转化成phylip 4格式(间断)(但名称全);再在记事本中打开,将“?”用“-”替换;用CIPRES Science Gateway[32]中在线RAxML-HPC2 Blackbaox[33]建ML进化树;在Figtree中编辑系统发育树。
2 结果与分析
2.1 不同菌株发酵酸乳凝乳时间比较
凝乳时间与乳酸菌在酸乳发酵过程中的生长繁殖、活力、产酸能力密切相关[1],本实验中27株酵母菌与保加利亚乳杆菌混菌发酵样品与对照组发酵样品在6 h都出现部分凝乳,8 h都完全凝乳,二者并未出现明显差别,酵母菌的加入对酸乳发酵中凝乳时间无明显影响。
2.2 不同菌株混菌发酵酸乳感官评价比较
感官评价是酸乳品质测评的重要手段,本实验以保加利亚乳杆菌单菌发酵酸乳为对照,对27株酵母菌与乳酸菌混合发酵后熟24 h酸乳从质地、气味和口感方面进行感官评价,由表2可得,从青海西宁牦牛酸乳分离的“QX1”~“QX15”和青海大通县酸乳中分离的“QD1”~“QD3”共18株酵母菌与保加利亚乳杆菌混菌发酵酸乳样品有明显的酒精味,掩盖了奶香味,但无其他异味,大多乳清析出也较多(质地得分约6分),部分有明显的“豆腐渣”状(质地得分约10分),口感偏甜(得分约10~15分)或酸甜分明(得分约20~25分),总体得分在80分以下;而从山东酸乳中分离的“SD1”、“SD2”和从西藏拉萨、那曲、山南酸牦牛乳中分离的“XS1”、“XN1”~“XN6”这9株酵母菌与保加利亚乳杆菌混菌发酵酸乳样品奶香味比较明显,也无明显的乳清析出,质地均匀细腻,酸乳适中,融合感较好,总体得分在80分以上。因此,选择“SD1”、“SD2”、“XS1”、“XN1”、“XN2”、“XN3”、“XN4”、“XN5”、“XN6”这9株酵母菌作为下一步实验菌株。

表2 不同菌株发酵酸乳感官评定得分Table 2 Sensory evaluation scores of yoghourt fermented with different strains
2.3 不同菌株发酵酸乳持水力比较
酸乳持水力指酸乳保质期内乳清析出的多少,反映不同质地酸乳对乳清的截留能力,越大表示酸乳质地越优。9株酵母菌“SD1”、“SD2”、“XS1”、“XN1”、“XN2”、“XN3”、“XN4”、“XN5”、“XN6”和保加利亚乳杆菌混菌发酵后熟24 h酸乳和对照酸乳样品的持水力均为100%,与感官评价中观察到的乳清析出情况一致。
2.4 不同菌株发酵酸乳质构分析比较
感官评价法一般采用暂时的评分系统,局限性很大,而且这种评定费时费力,受评价员的情绪、健康状况等多种因素影响,稳定性不足。质构仪通过模拟人的口腔咀嚼功能,用具体的数据,客观地将人的触觉感受分解成硬度、咀嚼性、弹性、内聚性、胶黏性等几个部分,9株酵母菌和保加利亚乳杆菌混菌发酵酸乳样品和对照酸乳样品质构数据如图1~图4所示,“XS1”菌株混菌发酵酸乳的质构指标最好。
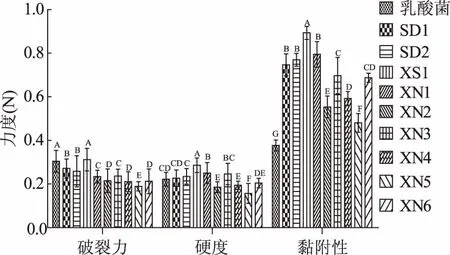
图1 不同菌株发酵酸乳破裂力、硬度、粘附性Fig.1 The rupture force,adhesiveness and hardness of yoghourt fermented with different strains注:不同字母的均值间存在显著差异(p<0.01),图2~图4,图6~图7、表3同。
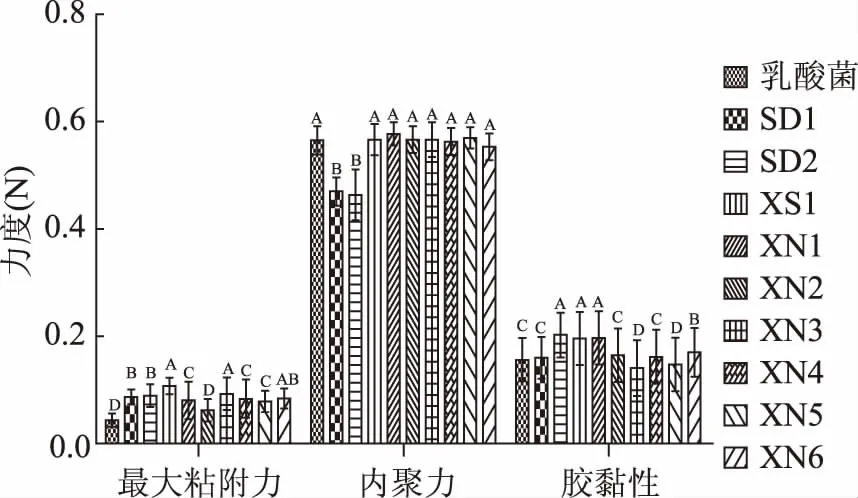
图2 不同菌株发酵酸乳最大粘附力、内聚力、胶黏性Fig.2 The tackiness,maximum adhesion force and hardness of yoghourt fermented with different strains
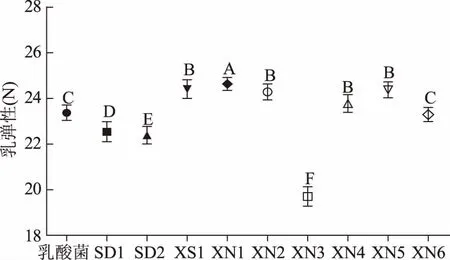
图3 不同菌株发酵酸乳弹性Fig.3 Elasticity of yoghourt fermented with different strains
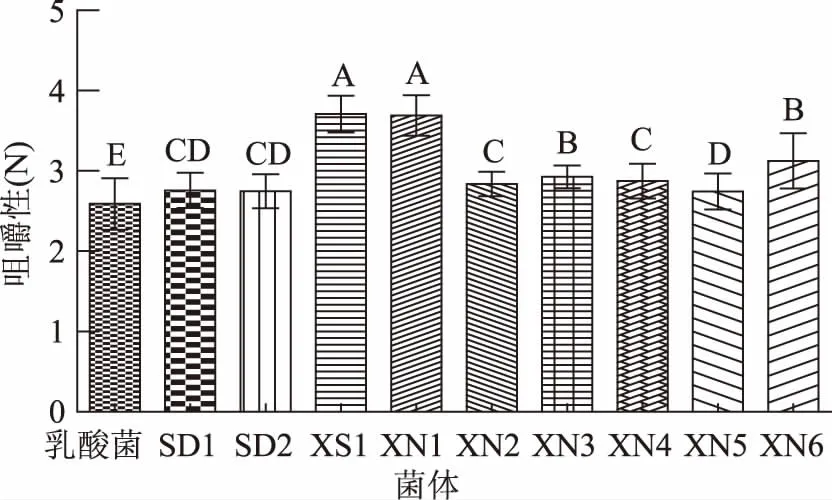
图4 不同菌株发酵酸乳咀嚼性Fig.4 Chewiness of yoghourt fermented with different strains
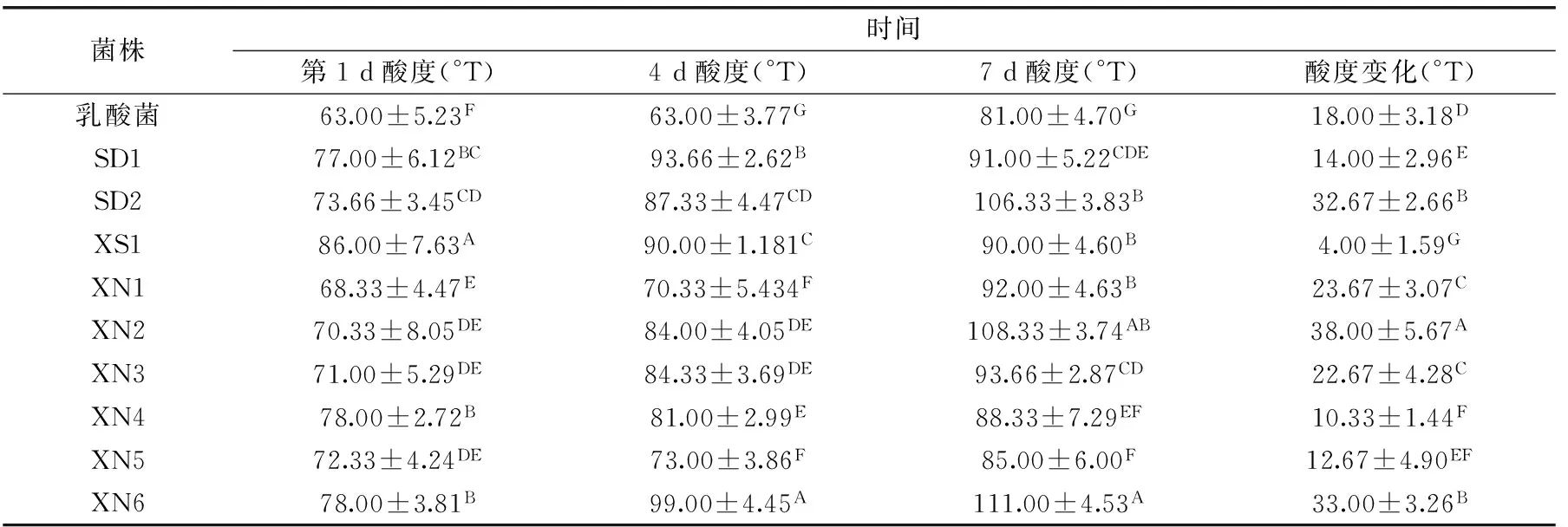
表3 不同菌株发酵酸乳滴定酸度变化Table 3 Titratable acidity of yoghourt with different strains
2.5 不同菌株发酵酸乳滴定酸度比较
发酵结束后,后熟好的酸乳,酸度要适中,同样发酵8 h后的样品,所有混菌发酵酸乳的酸度都比对照高,其中菌株“XS1”混菌发酵样品酸度最高;但冷藏保存7 d后,“XS1”混菌酸乳的后酸变化却最小,只有4 °T,远远低于对照。
2.6 不同菌株发酵酸乳产香能力比较
酸乳中双乙酰和乙醛含量与其香味有关,利用双乙酰标准曲线(图5),检测9株菌混合发酵酸乳样品以及单菌发酵对照样品中的双乙酰含量,除“XN3”、“XN2”、“XN5”混菌发酵酸乳双乙酰含量明显低于对照外(“XN3”、“XN2”、“XN5”的p值分别为0.009、0.003、0.004),其余所有的混菌发酵样品中双乙酰含量明显大于对照(“XN1”、“XN4”、“XN6”、“XS1”、“SD1”、“SD2”p值分别为0.008、0.008、0.008、0.0009、0.009、0.0008),其中“SD2”最高(14.83 μg/mL),“XS1”次之(12.65 μg/mL)(图6)。而在乙醛含量方面,只有“SD2”和“XN3”混菌发酵酸乳的乙醛含量均高于对照,“XS1”混菌发酵酸乳的乙醛含量低于对照,其余样品于对照无显著差异(图7)。
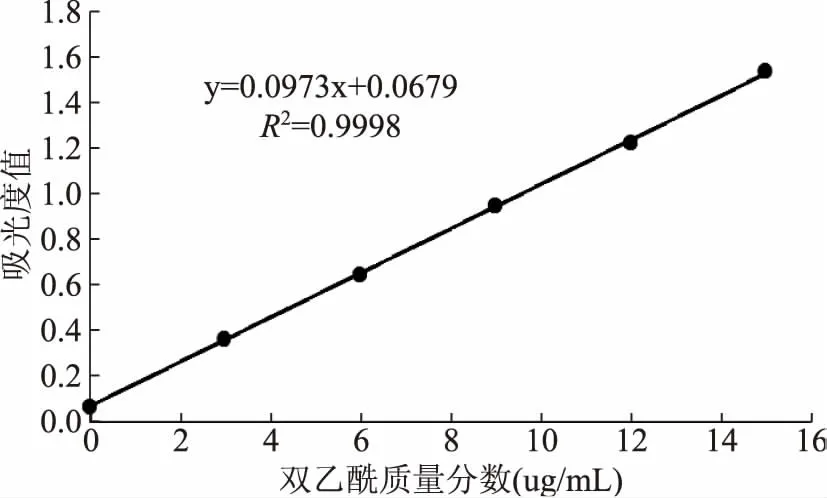
图5 双乙酰标准曲线Fig.5 The standard curve of diacetyl

图6 不同菌株发酵酸乳中双乙酰含量Fig.6 The content of diacetyl in yoghourtfermented with different strains

图7 不同菌株发酵酸乳中乙醛含量测定结果Fig.7 The content of acetaldehyde in yoghourt fermented with different strains
2.7 优良共生酵母菌株“XS1”的鉴定结果
菌株“XS1”在YPD培养基上菌落形态均为白色、圆形或椭圆形、不透明、有粘性、光滑湿润,美兰染色后显微孢子形态为椭圆形、圆形或卵圆形,可见出芽生殖(图8),提取该菌株基因组DNA,采用PCR扩增26S rDNA D1D2和ITS1序列。测序后分别获得约650 bp和300 bp序列,将该序列在NCBI中比对,结果显示“XS1”的26S rDNA D1D2和ITS1序列与Kluyveromycesmarxianus的序列相似度高达99%;基于Holzapfel等分类系统参考序列,利用“XS1”菌株的26S rDNA D1D2和ITS1区序列,分别构建进化树,结果显示“XS1”与Kluyveromycesmarxianus聚在一起(图9、图10)。得出“XS1”为马克斯克鲁维酵母(Kluyveromycesmarxianus)菌株。
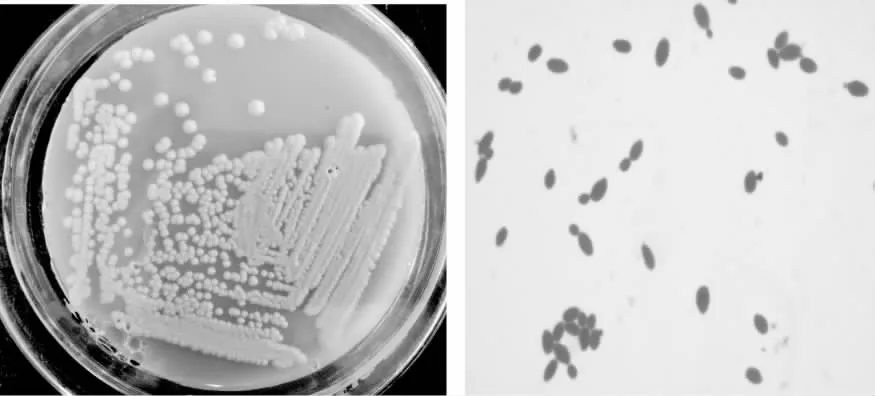
图8 “XS1”菌落图片和细胞显微图片Fig.8 The colony and cell of “XS1” strain

图9 基于26S rDNA D1D2区构建的Kluyveromyces属系统发育树Fig.9 The phylogenetic tree of Genus Kluyveromyces based on 26S rDNA D1D2 sequences
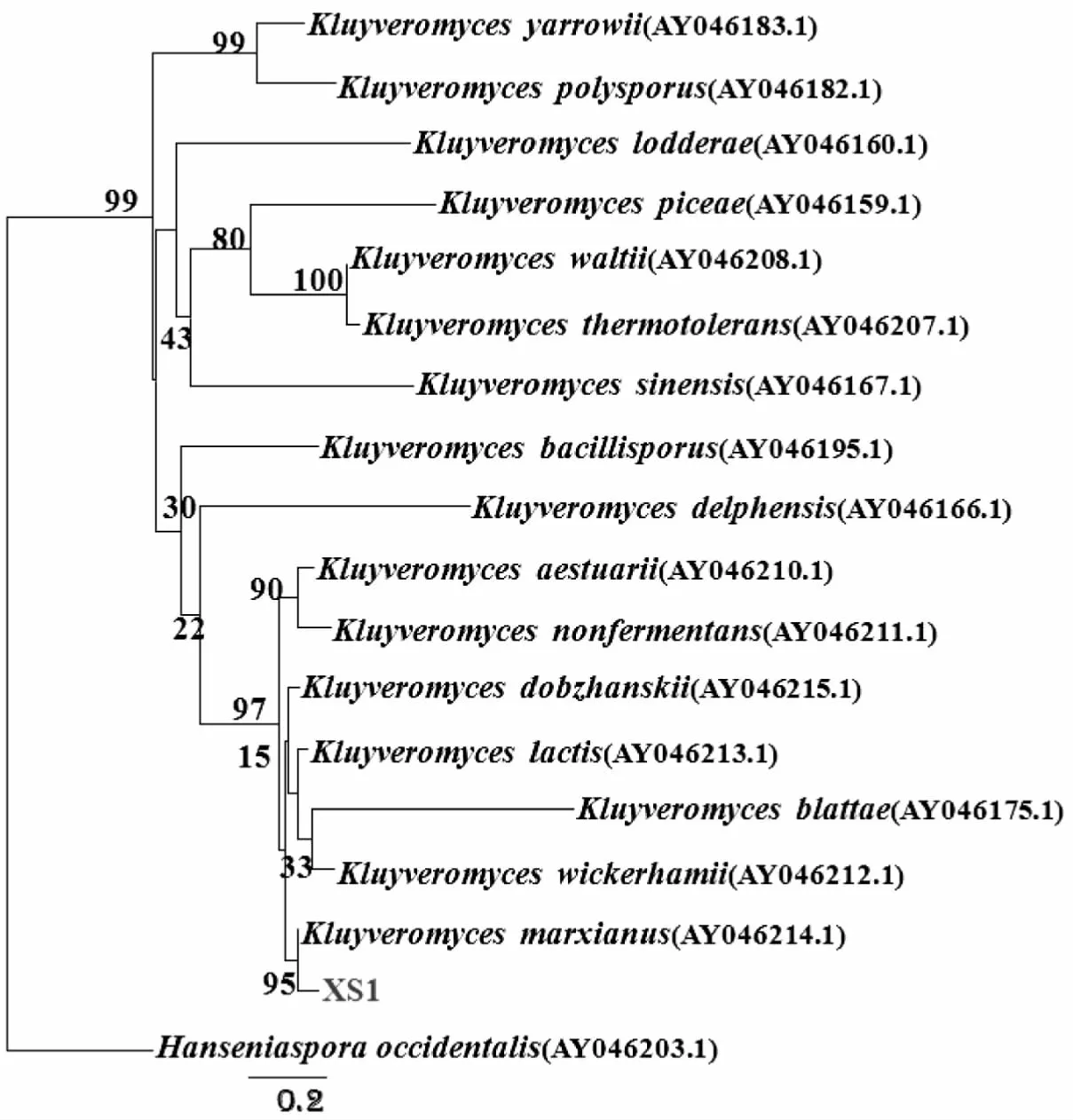
图10 基于ITS1区构建的Kluyveromyces属系统发育树Fig.10 The phylogenetic tree of Genus Kluyveromyces based on ITS1 sequences
3 讨论
乳酸菌和酵母菌是食品工业中应用最广泛的两种微生物。长期以来,乳制品中的酵母仅作为一种污染菌和腐败菌而备受人们关注,随着各种乳制品中酵母被大量分离,酵母菌在乳制品中的有益作用开始被关注,尤其是马克斯克鲁维酵母被发现是开菲尔[4-5]和酸马奶[11-13]中的优势酵母菌。随着对发酵乳制品中酵母菌研究的深入,越来越多的马克斯克鲁维酵母菌在其他各种乳制品中不断被分离出来[34],Bai等[20]分别从青海地区和西藏地区传统发酵牦牛酸乳中分离出马克斯克鲁维酵母;王远微等[35]报道对川西北部分牧区的传统发酵牦牛酸乳样品中酵母菌优势菌是马克斯克鲁维酵母;谢婕等[36]发现羊八井地区传统发酵牦牛酸乳中分离到的酵母菌中有17株是马克斯克鲁维酵母;李宇辉等[37]报道马克斯克鲁维酵母和酿酒酵母(Saccharomycescerevisiae)是新疆伊犁牧区少数民族家中采集奶疙瘩样品中的共同菌株;李银聪[38]发现西藏和川西青藏高原牧民自制的自然发酵酸耗牛奶也存在马克斯克鲁维酵母。本实验中筛选的马克斯克鲁维酵母菌株“XS1”从西藏山南地区收集的牦牛酸乳样品分离获得。
本实验将保加利亚乳杆菌和酵母菌混菌发酵制备酸乳,混菌发酵的优势在于通过不同代谢能力菌种的组合,完成单个菌种难以完成的多种复杂代谢反应,提高生产效率。目前关于混菌发酵过程中酵母菌和乳酸菌之间的共生作用研究甚少。有报道在乳制品发酵过程中,乳酸菌首先利用环境中的乳糖生长,通过自身的代谢途径反应生成乳酸降低了环境的pH,为酵母菌的代谢提供了条件,因为酵母菌最适宜在偏酸性环境中生长,同时一些乳酸菌发酵乳糖的副产物为半乳糖,能够为不发酵乳糖而利用半乳糖的酵母提供碳源;另一方面,乳酸菌产生的乳酸引起pH降低会对自身的代谢产生抑制作用,此时酵母菌在生长消耗乳酸盐,环境pH就不断升高,这样就减缓了环境中的pH太低而导致乳酸菌的反馈抑制。另外,酵母菌代谢所产生的物质,如丙酮酸、维生素和氨基酸等其他复合物也会刺激乳酸菌进一步代谢,这就为两种菌之间的相互促进提供了良好的基础[39-40]。本实验部分菌株混菌发酵结果也证明了这种相互促进作用,但仅使用一种保加利亚乳杆菌和酵母菌混合发酵酸乳进行研究,实际生产中常使用保加利亚乳杆菌和嗜热链球菌复合发酵,球菌、杆菌有各自不同的发酵特性,发酵产物也有差异,菌株越多相互关系就越复杂,以初步筛选的混菌发酵酸乳在感官评价得分较高9株酵母菌为基础,研究保加利亚乳杆菌、嗜热链球菌和酵母菌混合发酵对酸乳的影响,是本实验下一步的工作。
不同类型发酵酸乳中酵母菌和乳酸菌的相互作用可能存在差异,乳酸菌和酵母菌的种类及其接种比例、发酵条件都有可能影响两者之间的共生作用,本实验只选择了乳酸菌和酵母菌比例是3∶1,培养条件也只选择了保加利亚乳杆菌最佳生长温度40 ℃,对于其他条件参数还需后续研究。
4 结论
本实验以保加利亚乳杆菌单菌发酵为对照,从27株酵母菌中筛选获得1株马克斯克鲁维酵母菌株“XS1”,该菌株与保加利亚乳杆菌混菌发酵的酸乳在质地特性、后酸度、粘度、持水力、产香能力等方面表现优良,有望应用于开发独特风味和口感的酵母菌-乳酸菌复合发酵酸乳。
[1]张和平,张列兵. 现代乳品工业手册(第二版)[M]. 北京:中国轻工业出版社,2005:23-33.
[2]张兰威. 乳与乳制品工艺学(第二版)[M]. 北京:中国农业出版社,2006:16-28.
[3]任国谱,肖莲荣,彭湘莲. 乳制品工艺学[M]. 北京:中国农业科学技术出版社,2013:5-33.
[4]卢曼,陈小兵,王兴兴,等. 西藏开菲尔粒中酵母菌的分离与初步鉴定[J]. 上海海洋大学学报,2015,24(1):138-145.
[5]李艳,范佳. 自然发酵乳及传统开菲尔粒中酵母菌的多样性研究[J]. 河北科技大学学报,2014,35(6):561-568.
[6]钟浩. 开菲尔(Kefir)粒中菌种的分离鉴定及优良菌株的复合发酵乳研究[D]. 镇江:江苏大学,2016:33-57.
[7]周剑忠,董明盛,江汉湖. 藏灵菇微生物种群结构的分子特性研究[J]. 微生物学通报,2006,33(4):31-31.
[8]刘敏敏. 酸马奶中乳酸菌和酵母菌潜在共生菌筛选及发酵特性研究[D]. 呼和浩特:内蒙古农业大学,2011:26-41.
[9]赵美霞. 内蒙古地区酸马奶酒中酵母菌的分离鉴定及抗菌特性的研究[D]. 呼和浩特:内蒙古农业大学,2002:21-35.
[10]李丽杰. 酸马奶酒中酵母菌抑菌物质的粗提及生化特性研究[D]. 呼和浩特:内蒙古农业大学,2004:36-57.
[11]张积荣. 酸马奶中酵母菌的分离鉴定及发酵过程中乙醇含量变化的研究[D]. 乌鲁木齐:新疆农业大学,2007:10-28.
[12]李静,贾佳佳,杨艳,等. 新疆哈萨克族传统发酵酸马奶中酵母菌的分离鉴定[J]. 食品科学,2012,33(5):203-207.
[13]祝春梅,姚新奎,孟军,等. 新疆自然发酵酸马奶中酵母菌的分离鉴定[J]. 食品与发酵工业,2013,39(4):42-47.
[14]余兰,塔布斯·马那尔,新华·那比. 新疆传统发酵乳品中乳酸菌与酵母菌生长的相互影响[J]. 食品工业科技,2015,36(3):186-189.
[15]芦文娟. 新疆塔城地区原料乳及奶酪中酵母菌多样性的研究[D]. 石河子:石河子大学,2016:31-35.
[16]王文磊,鲁永强,苏东海,等. 蒙古奶酪凝块中酵母菌的分离鉴定[J]. 食品工业科技,2012,33(3):149-151.
[17]杨俊俊. 西藏牦牛奶渣中微生物的分离鉴定及优良乳酸菌的筛选[D]. 杨凌:西北农林科技大学,2014:17-40.
[18]卿蔓君,白梅,张勇,等. 西藏曲拉和云南乳饼中酵母菌的鉴定及其生物多样性[J]. 微生物学报,2010,50(9):1141-1146.
[19]李先胜. 传统乳制品中酵母菌的筛选、鉴定和应用研究[D]. 大连:大连工业大学,2012:19-34.
[20]Mei B,Manjun Q,Zhuang G,et al. Occurrence and dominance of yeast species in naturally fermented milk from the Tibetan Plateau of China[J]. Canadian Journal of Microbiology,2010,56(9):707.
[21]生庆海,张爱霞,马蕊. 乳与乳制品感官品评[M]. 北京:中国轻工业出版社,2009:13-18.
[22]张兰威. 乳酸菌优良菌株的选育及直投式酸奶发酵剂的研制[D]. 哈尔滨:东北农业大学,2002:20-32.
[23]徐鑫,赫君菲,何佳易,等. 测试条件对凝固型酸奶质构实验参数的影响[J]. 食品科学,2013,34(24):263-267.
[24]赫君菲. 建立凝固型酸奶质构参数对感官性质的预测模型[D]. 扬州:扬州大学,2013:14-44.
[25]杨莹莹,贺红军,郭萌萌,等. 响应面法分析TPA测试参数对酸奶质构的影响[J]. 现代食品科技,2013,(6):1281-1284.
[26]中华人民共和国国家卫生和计划生育委员会. GB 5009.239-2016食品酸度的测定[S]. 北京:中国标准出版社,2016.
[27]刘宁宁,郭红敏,葛春美,等. 酸奶中乙醛和双乙酰含量对其风味的影响[J]. 中国食品添加剂,2012(s1):269-273.
[28]席晓霞,于洁,王宏梅,等. 发酵乳中嗜热链球菌产乙醛和双乙酰的动态变化研究[J]. 中国乳品工业,2014,42(7):20-23.
[29]Kurtzman C P,Robnett C J. Phylogenetic relationships among yeasts of the ‘Saccharomycescomplex’ determined from multigene sequence analyses[J]. Fems Yeast Research,2003,3(4):417.
[30]Tamura K,Stecher G,Peterson D,et al. Mega6:Molecular evolutionary genetics analysis version 6.0[J]. Molecular Biology and Evolution,2013,30:2725-2729.
[31]Xia X,Xie Z. Dambe:Software package for data analysis in molecular biology and evolution[J]. Molecular Biology and Evolution,2013,30(7):1720-1738.
[32]Miller MA,Pfeiffer W,Schwartz T. Creating the cipres science gateway for inference of large phylogenetic trees[A]Gateway Computing Environments Workshop(GCE). New Orleans:Ieee Xplore,2010:1-8.
[33]Stamatakis A. RAxML version 8:A tool for phylogenetic analysis and post-analysis of large phylogenies[J]. Bioinformatics,2014,30(9):1312-1313.
[34]Latorre-García L,Castillo-Agudo L D,Polaina J. Taxonomical classification of yeasts isolated from kefir based on the sequence of their ribosomal RNA genes[J]. World Journal of Microbiology & Biotechnology,2007,23(6):785-791.
[35]王远微,张诚民,索化夷,等. 传统发酵牦牛酸奶中马克斯克鲁维酵母菌的分离鉴定及系统发育分析[J]. 食品科学,2014,35(15):216-220.
[36]谢婕,赵欣,骞宇,等.传统发酵牦牛酸乳中酵母菌的分子生物学鉴定[J]. 食品科学,2015,36(11):114-118.
[37]李宇辉,王俊钢,刘成江,等.新疆伊犁牧区发酵乳制品中酵母菌的分离和多样性分析[J]. 食品与发酵工业,2013,39(7):98-103.
[38]李银聪. 自然发酵酸牦牛奶的微生物区系及其抗氧化活性研究[D]. 重庆:西南大学,2011:76.
[39]álvarez-Martín P,Flórez A B,Hernández-Barranco A,et al. Interaction between dairy yeasts and lactic acid bacteria strains during milk fermentation[J]. Food Control,2008,19(1):62-70.
[40]Viljoen B C. The interaction between yeasts and bacteria in dairy environments[J]. International Journal of Food Microbiology,2001,69(1-2):37-44.
