飞蝗表皮蛋白基因LmNCP1的分子特性及功能分析
2018-04-18杨亚亭赵小明秦忠玉刘卫敏马恩波张建珍
杨亚亭,赵小明,秦忠玉,刘卫敏,马恩波,张建珍
飞蝗表皮蛋白基因的分子特性及功能分析
杨亚亭1,2,赵小明1,秦忠玉1,2,刘卫敏1,马恩波1,张建珍1
(1山西大学应用生物学研究所,太原 030006;2山西大学生命科学学院,太原 030006)
【】基于飞蝗()转录组数据库搜索并克隆获得飞蝗表皮蛋白基因(nymph cuticle protein 1),分析其序列特征和表达特性,通过蜕皮激素(20-hydroxyecdysone,20E)诱导和干扰20E受体基因表达研究其表达调控,并基于RNA干扰(RNAi)方法分析其生物学功能,为阐明该基因在飞蝗表皮发育过程中的作用提供理论依据。【】依据飞蝗转录组数据库获得表皮蛋白基因,结合RT-PCR技术克隆获得其全长开放阅读框(ORF)序列,并结合飞蝗基因组序列分析其基因结构及序列特征;使用MEGA 6.0软件中邻接法(neighbor-joining,NJ),与其他昆虫同源序列聚类分析,并根据聚类结果进行命名;利用reverse-transcription quantitative PCR(RT-qPCR)分析其在5龄飞蝗不同组织和不同发育天数的基因表达特性;通过体内注射20E诱导以及干扰20E受体基因的表达,RT-qPCR检测该基因的表达情况;基于RNAi结合H&E染色方法分析其生物学功能。【】通过搜索获得表皮蛋白基因,并进行了克隆和测序验证,获得457 bp的cDNA序列,其全长ORF序列为306bp,编码101个氨基酸。氨基酸序列分析表明该基因编码的蛋白含有1个信号肽,不含几丁质结合域,但包含3个重复基序,Weblogo分析结果显示其在物种间具有保守性。系统进化树分析显示该蛋白与蜚蠊目蟑螂()BcNCP1序列一致度最高,聚为一支。根据聚类结果将该蛋白命名为LmNCP1(GenBank登录号:MF326211)。RT-qPCR结果显示在5龄若虫体壁中高表达,在翅芽、前肠和脂肪体中表达次之,在中肠、后肠、胃盲囊和马氏管中表达量较低或不表达;在5龄早期(N5D1和N5D2)高表达,随后表达量逐渐降低(N5D3—N5D6),到下一次蜕皮前表达有所升高(N5D7),其表达量动态与内表皮形成时间一致。20E诱导不同时间结果显示,与对照组相比,在20E诱导1 h和3 h后表达量显著上调,分别上调了1.3倍和0.9倍;利用RNAi方法干扰48 h和72 h后,与对照组相比,表达量均显著降低,分别降低了71%和87%。利用RNAi方法在4龄和5龄第2天分次注射ds后,飞蝗仍能够正常蜕皮,无致死表型,但对其表皮进行H&E染色发现,与对照组相比,其内表皮形成减少,导致表皮显著变薄,表皮层厚度约减少了33%。【】飞蝗表皮蛋白LmNCP1含有3个物种间保守的重复基序,不含几丁质结合域,属于其他家族蛋白;主要在体壁中高表达,而且响应介导的20E信号通路调控;RNAi试验表明,在飞蝗蜕皮过程中参与内表皮的沉积。
飞蝗;表皮蛋白;;蜕皮激素
0 引言
【研究意义】昆虫体壁覆盖整个虫体,具有维持虫体形态、抑制体内水分蒸发和抵御外源物质入侵等作用[1],其主要由上表皮、原表皮和真皮细胞层组成,原表皮由内表皮和外表皮构成,其中上表皮和外表皮在蜕皮前形成,内表皮在蜕皮后形成[2]。昆虫表皮的主要成分为几丁质和表皮蛋白,两者约占昆虫表皮成分的60%以上,其成分的缺失或含量减少均会导致表皮功能异常甚至丧失,从而影响昆虫的正常生长发育[3]。蜕皮激素(20-hydroxyecdysone,20E)在调控昆虫蜕皮、变态和生殖中具有重要作用。目前,对表皮蛋白的研究主要集中于模式昆虫,而对于非模式的渐变态昆虫研究较少。飞蝗()是一种典型的渐变态昆虫,同时也是一种重要的世界性农业害虫。因此,以飞蝗为对象研究渐变态昆虫表皮蛋白基因的分子特性、表达模式、生物学功能及受20E介导的表达调控方式,可以为表皮蛋白基因在20E介导的蜕皮过程中的作用及调控机制研究提供依据。【前人研究进展】随着昆虫基因组学的发展,越来越多的表皮蛋白被鉴定,其种类繁多,功能也不尽相同。目前已报道的昆虫表皮蛋白序列已有1 400条以上,主要分为5个家族,包括CPR家族、CPF/CPFL家族、Tweedle家族、CPAPs家族以及其他家族。其中,CPR家族是数量最多的表皮蛋白家族,根据其在表皮层的类型和位置的差异性,CPR家族可分成3个亚族,即RR-1、RR-2和RR-3,据报道含有RR-1基序的表皮蛋白主要存在于柔软表皮层,含有RR-2基序的表皮蛋白主要存在于坚硬表皮层[4],目前对含有RR-3基序的表皮蛋白研究报道不多,具体功能未知。CPAPs家族根据其含有的几丁质结合域可分为CPAP1和CPAP3两种类型,在昆虫生长发育中具有重要功能[5-8]。Tweedle家族是含有Tweedle基序的一类表皮蛋白,在昆虫体形和生长发育中具有重要作用[9]。CPF/CPFL是低复杂度表皮蛋白家族,该家族含有44个保守的氨基酸序列,在黄粉虫()和飞蝗中被首次鉴定[10]。随后,Togawa等[11]从冈比亚按蚊()中鉴定出4个CPF家族表皮蛋白基因,并发现这4个表皮蛋白基因只在蛹期或羽化前期表达,表明其可能与成虫体壁上表皮的形成有关。然而,仍有一些表皮蛋白不属于上面这些家族中的任一家族,被称为其他家族,如Jensen等[12]从死人头蟑螂()中鉴定了4个蜕皮后表皮蛋白(BcNCP1、BcNCP2、BcNCP3和BcNCP4),该类表皮蛋白含有2—3个重复基序,但不含几丁质结合域。该家族表皮蛋白在昆虫表皮中发挥怎样的作用仍不清楚。另外,昆虫表皮蛋白基因的表达受到多种因素的影响,如激素(20E和JH)、转录因子和内含子等,这些因素之间相互作用,使昆虫表皮蛋白基因的表达形成一个多级调控系统[13]。在昆虫蜕皮过程中20E与其受体(EcR)结合通过其响应因子调控表皮蛋白基因的表达[13],如在黑腹果蝇()和烟草天蛾()中研究发现FTZ-F1能够促进表皮蛋白基因的表达[14-15]。在家蚕()中,翅原基表皮蛋白基因受到多个转录因子激活表达,并响应20E的诱导[16-20]。但20E对表皮蛋白基因的诱导调控研究大多集中在CPR家族基因,对其他家族基因的研究较少。在飞蝗中,笔者课题组前期基于转录组数据库获得了81个表皮蛋白基因,根据其保守基序分为CPR家族、CPAPs家族、Tweedle家族、CPF/CPFL家族以及其他家族[21]。随后,对CPR家族的内表皮蛋白基因研究发现其主要在外胚层形成的组织中表达,可能参与内表皮的形成[22];王燕等[23]基于RNAi方法对CPAPs家族表皮蛋白研究,发现其在飞蝗生长发育过程中具有重要作用;Song等干扰Tweedle家族表皮蛋白基因后引起飞蝗蜕皮困难死亡[24],但对其他家族基因的研究尚未见报道。【本研究切入点】目前,关于飞蝗表皮蛋白的研究还比较少,主要集中于表皮蛋白的提取、鉴定和分类。对一些不含几丁质结合域的表皮蛋白的分子特性及其基因表达模式尚缺乏系统研究。另外,该类表皮蛋白基因是否响应20E诱导表达以及在20E介导的蜕皮过程中具有怎样的功能,有待深入研究。【拟解决的关键问题】以飞蝗表皮蛋白基因为对象,研究其分子特性、组织分布和时期表达特性及其响应20E诱导表达情况,并通过RNA干扰结合H&E染色方法分析其生物学功能,为明确其在飞蝗蜕皮过程中的功能提供依据。
1 材料与方法
试验于2016—2017年在山西大学应用生物学研究所完成。
1.1 试验材料
供试昆虫:飞蝗虫卵购自河北沧州蝗虫养殖公司,将购买的飞蝗虫卵置于恒温人工气候培养箱内进行孵化,待其孵化成1龄若虫时移至养虫笼内置于人工气候培养箱中,种植新鲜小麦苗用于饲喂,待其长至3龄时开始辅以麦麸饲喂。人工气候培养箱内设置温度为(30±2)℃,相对湿度为(40±5)%,光周期为光照14 h﹕黑暗10 h。
试验试剂:RNAisoTMPlus,Reverse transcriptase M-MLV(RNaseH)购自TaKaRa公司;SYBR®Green real-time PCR Master Mix购自TaKaRa公司;2×Tag PCR Master Mix购自天根公司;AxyPrepTMPCR Cleanup Kit试剂盒购于AXYGEN公司。
1.2 表皮蛋白基因获得及其分子特性分析
基于课题组飞蝗转录组数据库,获得表皮蛋白基因Unigene3699,结合飞蝗基因组数据库[25],获得其全长ORF序列,克隆验证后根据飞蝗基因组序列对其基因结构进行分析,引物见表1(由上海生工生物有限公司合成)。利用ExPASy网站translate tool对其ORF序列进行翻译、并预测蛋白分子量大小及等电点(pI),利用ExPASy网站ProtParam tool分析其带负电荷的酸性氨基酸(Asp+Glu)和带正电荷的碱性氨基酸(Arg+Lys)、蛋白脂肪系数、不稳定指数以及总水平亲水性系数(grand average of hydropathicity,GRAVY),SignalP4.1 server分析其信号肽,利用SMART分析其结构域,通过WEBLOGO工具对其在各物种间的保守基序进行分析。
1.3 系统进化树分析
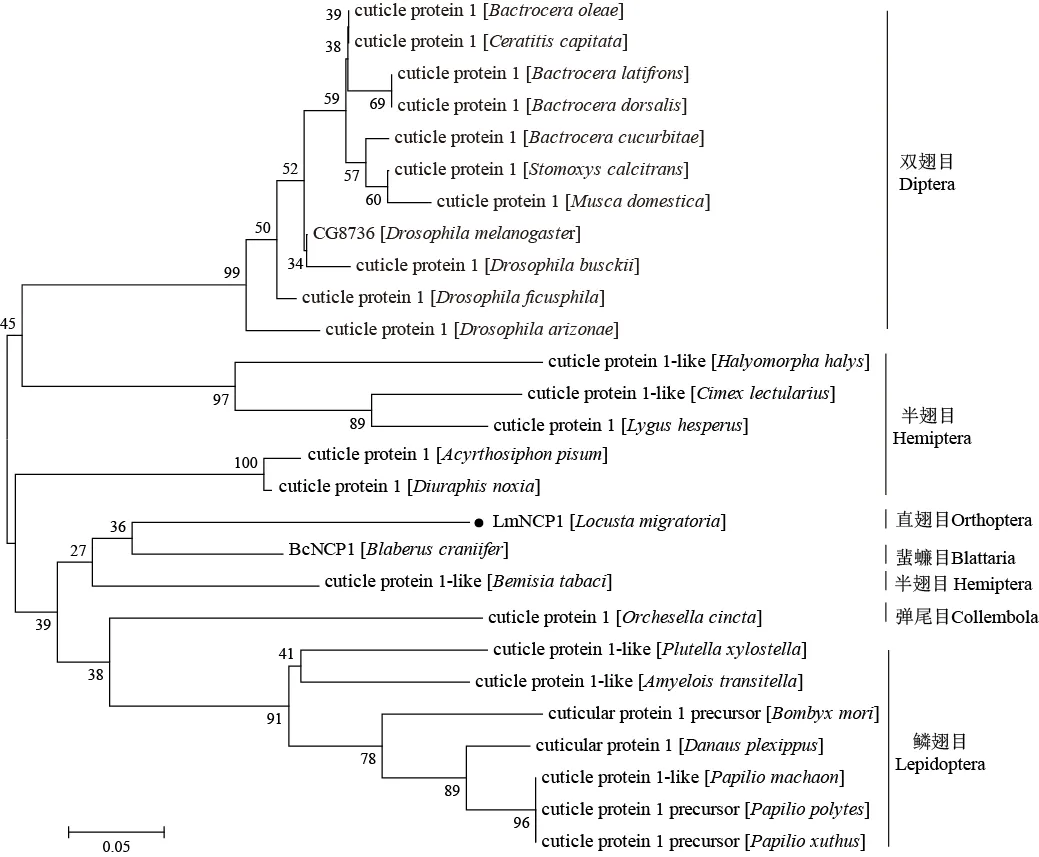
根据Unigene3699氨基酸序列,运用NCBI的BLAST工具搜索其同源序列并进行比对,下载来源于其他物种的同源序列,使用Mega 6.0软件中邻接法(neighbor-joining,NJ),将Unigene3699氨基酸序列与死人头蟑螂、柑橘凤蝶()、黑腹果蝇等物种的同源序列进行聚类分析,独立分析1 000次,数值代表bootstrap估算值。根据聚类结果将其命名为LmNCP1(nymph cuticle protein 1)。用于构建进化树的不同目物种的表皮蛋白(cuticle protein 1,CP1)和飞蝗序列的GenBank登录号见表2。
1.4 mRNA表达特性
1.4.1 表达的组织特异性检测 为了探讨在不同组织的表达特性,选用5龄第2天飞蝗若虫,雌虫和雄虫各半,设置3个生物学重复,每个生物学重复4头试虫,解剖体壁、翅芽、前肠、胃盲囊、中肠、后肠、马氏管和脂肪体8个组织,立即冻存于液氮中。按照TaKaRa公司的试剂说明书,用RNAisoTMPlus提取上述组织的总RNA,将所提取RNA用1%的琼脂糖凝胶电泳检测其质量,然后使用Nanodrop 2000对所取总RNA进行定量,并以1 μg总RNA为模板,根据TaKaRa反转录试剂盒说明书,合成cDNA。
利用RT-qPCR方法检测5龄第2天飞蝗不同组织部位的表达情况,反应体系和程序参考文献[22]。采用ABI Prism 7300 SDS 1.1软件对数据进行分析并记录。每个样本作2个技术重复,以作为内参(KX276642)[26-27],引物由上海生工生物有限公司合成(表1)。
1.4.2 不同发育天数的5龄若虫体壁中的表达量检测 为了探讨表达的时间动态,选取并解剖5龄第1—7天飞蝗若虫的第2—3腹节处体壁并冻存于液氮中,设4个生物学重复,每个生物学重复4头试虫。总RNA提取和cDNA模板合成方法同1.4.1。采用RT-qPCR方法检测在不同发育天数的5龄若虫体壁中的表达情况,引物见表1。
1.5 20E诱导表达分析
根据课题组20E处理的表皮转录组数据库[28],发现与对照组相比,在20E处理组中的表达量显著上升。为了进一步验证这一结果,对5龄第2天若虫进行体内注射20E,具体参考Li等[29]的方法。以注射10%乙醇的若虫为对照组,分别处理1、3、6和12 h后解剖飞蝗若虫第2—3腹节处体壁并冻存于液氮中,设3个重复,每个生物学重复4头试虫。总RNA提取和cDNA模板合成方法同1.4.1。采用RT-qPCR方法检测在20E处理组和对照组中的表达差异,具体方法同上。
1.6 干扰20E核受体后LmNCP1的表达变化
为了进一步检测响应20E诱导表达情况,基于RNA干扰方法合成并向5龄第2天若虫注射10 μg 20E受体基因(GenBank登录号:AAD19828.1)双链RNA(ds),以注射ds(绿色荧光蛋白基因的双链RNA)为对照,ds片段选取位置为和的公共区域,具体参考Liu等[30]的方法。48 h和72 h后取飞蝗若虫第2—3腹节处体壁并冻存于液氮中,设3个重复,每个生物学重复4头虫子。总RNA提取和cDNA模板合成方法同上。采用RT-qPCR方法检测的沉默效率,同时检测在干扰48 h和72 h后的表达变化,具体方法同上,引物见表1。

表1 本研究中所用引物

表2 本研究中用于构建系统进化树的物种和NCP1的GenBank登录号
1.7 基于RNAi的LmNCP1功能分析
扩增基因片段(5′-3′:158—451 bp),利用1.6中方法合成并向4龄第2天若虫注射10 μg该基因双链RNA(ds),以注射ds为对照,24 h后取对照组和处理组飞蝗体壁组织,利用1.6中方法检测的沉默效率,其余若虫正常饲养观察表型。若飞蝗正常蜕皮则于5龄第2天再次注射10 μg ds,正常饲养观察表型。
1.8 H&E染色
为了观察干扰后对飞蝗表皮形成的影响,分别解剖对照组和处理组5龄蜕皮后72 h飞蝗体壁组织。取其一部分腹部(第2、3腹节)表皮,用3%戊二醛固定,4℃过夜以制备石蜡切片,另一部分速冻后提取RNA再次检测沉默效率。固定的样品经石蜡包埋后进行切片(5 μm),然后用苏木精和伊红(H&E)染色,具体方法参照文献[24],封片后利用显微镜(Olympus BX51,Japan)观察表皮结构。
1.9 数据分析
基因相对表达量采用2-△△Ct [31]法进行分析,采用student-test方法进行差异表达分析,*表示在<0.05水平差异显著,**表示在<0.01水平差异显著,***表示在<0.001水平差异显著。
2 结果
2.1 LmNCP1特性分析
根据课题组飞蝗转录组数据库获得一个表皮蛋白基因Unigene3699,结合飞蝗基因组序列获得其cDNA序列并克隆、测序验证,结果显示该基因cDNA序列长457 bp,具有一个306 bp的开放阅读框(ORF);基因结构分析显示该基因含有两个外显子(图1-A),编码101个氨基酸,预测分子量为10.5 kD,等电点为7.5,其编码的蛋白N末端含有一个信号肽,不含几丁质结合域,但具有3个重复基序(图1-B),而且该重复基序在不同物种的同源蛋白中具有高度保守性(图1-C)。该蛋白有3个带负电荷的酸性氨基酸(Asp+Glu),4个带正电荷的碱性氨基酸(Arg+Lys),脂肪系数为69.80%,不稳定指数为66.89。亲/疏水性分析发现,总水平亲水性系数为0.226,属亲水性蛋白。

A:LmNCP1基因结构Gene structure of LmNCP1;B:LmNCP1序列分析Sequence analysis of LmNCP1;C:LmNCP1保守基序分析 Analysis of conserved motifs by Weblogo。下划线为信号肽The underline: signal peptide;方框为重复基序The box: Repeated motif
2.2 LmNCP1系统进化树分析
将获得的飞蝗NCP1氨基酸序列与其他昆虫的CP1氨基酸序列进行聚类分析,聚类结果显示,飞蝗NCP1与蜚蠊目、鳞翅目、半翅目烟粉虱和弹尾目昆虫的CP1聚为一支,其中与蜚蠊目昆虫BcNCP1亲缘关系最近(图2)。因此,根据其同源性将该蛋白命名为LmNCP1。
2.3 LmNCP1组织和时期表达特性分析
组织表达显示,在飞蝗体壁、翅芽、前肠和脂肪体中均有表达,胃盲囊、中肠、后肠和马氏管表达较低或不表达,其中在体壁中表达量最高,约为翅芽和前肠中表达量的8—10倍(图3-A),表明可能参与体壁的形成。
对不同发育天数的5龄若虫体壁组织的RT-qPCR检测结果显示,主要在上一个龄期蜕皮后高表达(N5D1-N5D2),随后表达量逐渐降低(N5D3-N5D6),到下一个龄期蜕皮前表达有所上升(N5D7),其中在蜕皮后第1天(N5D1)的表达量最高,约为N5D6表达量的55倍(图3-B),这一表达模式与内表皮形成时间一致,推测其可能在飞蝗蜕皮过程中参与内表皮形成。蜕皮前后时期表达分析发现,在蜕皮前表达量极低,蜕皮后表达量逐渐升高(图3-C)。
用于构建系统进化树的物种和NCP1的GenBank登录号见表2 Species and GenBank accession number for phylogenetic analysis are listed in table 2

A:LmNCP1在5龄飞蝗不同组织中的表达The relative expression of LmNCP1 in different tissues of 5thinstar nymphs。IN:体壁 Integument;WP:翅芽 Wing pads;FG:前肠 Foregut;GC:胃盲囊 Gastric caeca;MG:中肠 Midgut;HG:后肠 Hindgut;MT:马氏管 Malpighian tubules;FB:脂肪体 Fat body。B:LmNCP1在5龄飞蝗不同发育天数的表达The relative expression of LmNCP1 in different days of 5thinstar nymphs。N5D1—N5D7:5龄第1—7天 1st-7th day of 5thinstar nymphs。C:LmNCP1在4龄到5龄蜕皮过程中的表达The relative expression of LmNCP1 during molting from 4th instar to 5th instar nymphs。BE:蜕皮前Before ecdysis;E:蜕皮中ecdysis;AE:蜕皮后After ecdysis
2.4 20E诱导表达
为了检测是否响应20E诱导表达,选取5龄第2天飞蝗若虫进行体内20E诱导,RT-qPCR检测在20E诱导1、3、6和12 h后的表达变化。结果如图4所示,与对照组相比,在20E诱导1 h和3 h后表达量显著上调,约为对照组表达量的1.3倍和0.9倍,表明其响应20E诱导表达。
2.5 干扰20E受体基因LmEcR后LmNCP1的表达变化
为了进一步检测响应20E信号通路的调控,利用RNAi方法向5龄第2天若虫注射20E受体基因的双链RNA,48 h和72 h后检测表达变化情况。结果显示,注射ds48 h和72 h后的表达被显著抑制,其沉默效率为86%和82%(图5-A),同时的表达量均显著下调,分别降低了71%和87%(图5-B),表明受介导的20E信号通路调控。

图4 20E诱导后不同时间LmNCP1的相对表达量
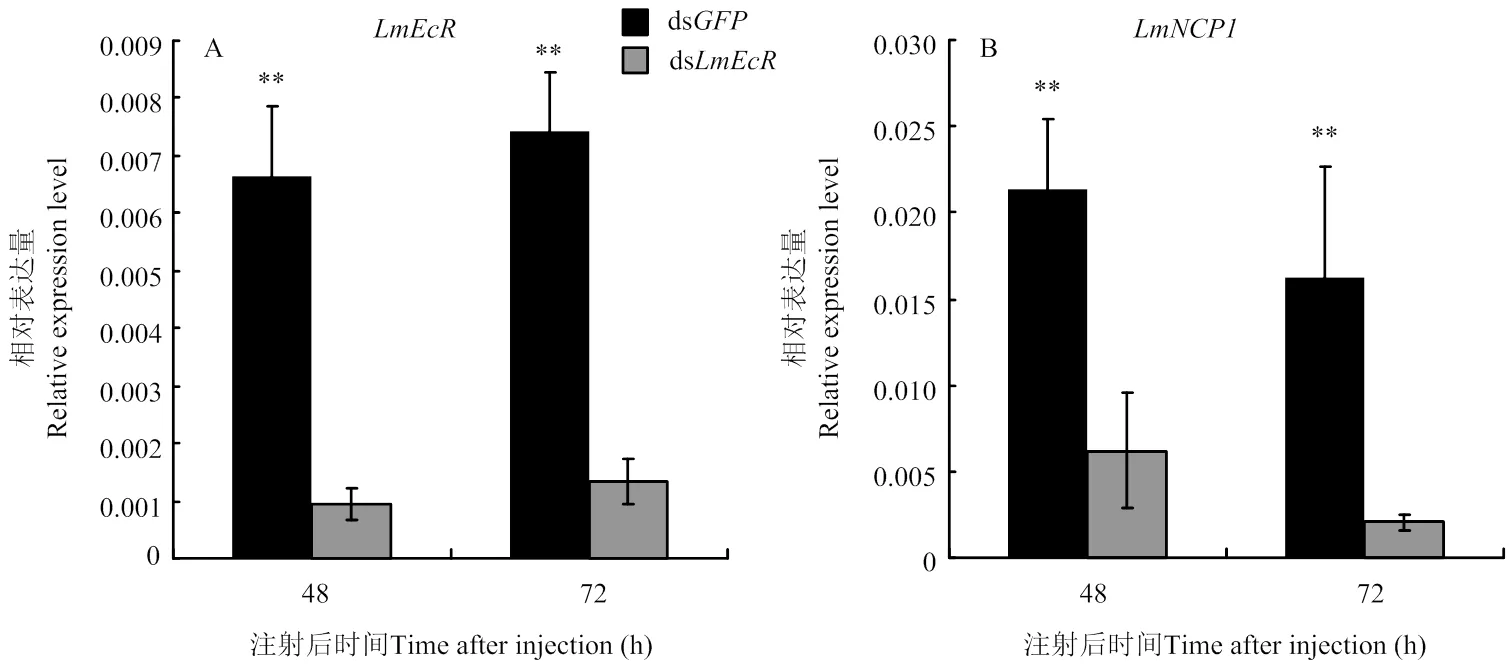
A:LmEcR沉默效率检测The relative expression of LmEcR determined by RT-qPCR after injected with dsLmEcR;B:干扰LmEcR后LmNCP1表达检测The relative expression of LmNCP1 determined by RT-qPCR after injected with dsLmEcR
2.6 基于RNAi的LmNCP1生物学功能分析
为了进一步研究对表皮形成的影响,首先向4龄第2天若虫注射10 μg ds,以注射ds为对照。结果发现,与对照组相比,注射ds的飞蝗的表达能够被显著抑制,但飞蝗均能够正常蜕皮进入5龄。随后于正常蜕皮的5龄第2天飞蝗再次注射10 μg ds,尽管与对照组相比,处理组的表达量下降了98%(图6-A),但飞蝗仍能够正常蜕皮变为成虫。为了观察该基因的缺失是否对飞蝗表皮结构产生影响,分别解剖对照组和处理组5龄蜕皮后72 h的成虫体壁组织,制作石蜡切片进行H&E染色。结果显示对照组原表皮结构正常,具有明显的外表皮和内表皮(图6-B和6-B’),但处理组原表皮层只含有外表皮,几乎没有内表皮形成(图6-C和6-C’),厚度测量统计分析显示处理组原表皮层相对于对照组显著变薄,表皮层厚度减少了33%(图6-D),表明参与了飞蝗蜕皮过程中内表皮的形成。

A:LmNCP1沉默效率检测The relative expression of LmNCP1 determined by RT-qPCR after injected with dsLmNCP1。B、B’:注射dsGFP后,表皮显微结构(B)以及选择放大区域(B’)After injection of dsGFP, microstructure (B) and magnification (B’) of cuticle。C、C’:注射dsLmNCP1后,表皮显微结构(C)以及选择放大区域(C’)After injection of dsLmNCP1, microstructure (C) and magnification (C’) of cuticle;红色为细胞质及细胞外基质,深蓝色为细胞核cytoplasm and extracellular matrix are stained red with eosin, nuclei are stained dark blue with hematoxylin (Scale bars, 20 μm);Pro:原表皮Procuticle;Exo:外表皮Exocuticle;Endo:内表皮Endocuticle。D:对照组与处理组原表皮层厚度(n=8)Relative thickness of procuticle between dsGFP and dsLmNCP1
3 讨论
本文根据课题组转录组数据库获得一个表皮蛋白基因,结合飞蝗基因组序列获得其全长ORF序列,并通过基因结构和序列分析发现,该基因只有2个外显子,编码的蛋白不具有几丁质结合域,但具有3个重复基序(C-P-x-P-x-C),而且这3个重复基序在物种间高度保守。前人研究也有类似发现,如Jensen等[12]从蟑螂若虫体壁中提取并鉴定出4个蜕皮后表皮蛋白,其中BcNCP1含有3个重复基序,根据其序列特征推测其可能特异性地与金属离子结合或与其他蛋白质相互作用。然而基于笔者课题组飞蝗转录组数据库仅发现一个该类型的表皮蛋白LmNCP1。系统进化树分析发现,LmNCP1与蜚蠊目蟑螂BcNCP1之间的亲缘性最近。
5龄飞蝗组织特异性表达分析发现,在飞蝗体壁中高表达,表明其可能与体壁的形成有关;时期表达分析发现,在5龄早期(上一个龄期蜕皮后)高表达(N5D1-N5D2),之后表达量逐渐降低(N5D3-N5D6),到下一个龄期蜕皮前表达有所上升(N5D7)。蜕皮前后时期表达分析发现,在蜕皮前表达量极低,蜕皮后表达量逐渐升高。昆虫的内表皮在蜕皮后逐渐形成,约在蜕皮后72 h完全形成[2],这种表达模式与昆虫内表皮形成时间一致,推测其可能参与内表皮结构的形成。目前,RNAi技术是研究基因生物学功能的有效工具,而且飞蝗对RNAi具有高度敏感性。因此,基于RNAi方法沉默该基因后,与对照组相比,飞蝗能够正常蜕皮进入下一龄期,然而H&E染色发现干扰的虫体内表皮形成减少,与对照组相比原表皮层极显著变薄(减少33%),表明其在飞蝗蜕皮过程中参与了内表皮结构的形成。随着昆虫基因组学和转录组学的发展,越来越多的表皮蛋白基因被鉴定,然而关于表皮蛋白基因的功能研究,尤其是内表皮蛋白基因的功能研究报道较少。Xiong等从家蚕体形突变体中鉴定出一个表皮蛋白基因,研究发现其参与家蚕内表皮的形成[32],然而其作用机制有待深入研究。
研究表明,20E在昆虫蜕皮过程中具有重要作用,而表皮蛋白是结构蛋白,已有研究表明表皮蛋白基因的表达在昆虫表皮形成中受20E诱导调控[13]。根据笔者课题组前期20E处理的表皮转录组数据库[28],发现与对照组相比,在20E处理组中的表达量显著上升,暗示其可能响应20E诱导调控。为进一步研究其是否受20E信号调控,首先选取5龄第2天若虫注射20E进行不同时间诱导,结果发现在20E诱导后1 h和3 h均能够上调表达;随后,通过RNAi方法沉默20E受体基因后发现,与对照组相比,表达显著下调,表明响应介导的20E信号通路调控。然而,时期表达模式与20E滴度变化并不一致[30]。有研究发现,一些表皮蛋白基因,其转录水平的上调需要20E调控,但这些表皮蛋白的基因转录只有在20E信号降低时才会启动[33-34]。因此,笔者推测可能响应低浓度的20E诱导,但其受20E信号通路调控的具体机制以及在表皮形成中的作用机制还有待进一步研究。
4 结论
飞蝗LmNCP1具有3个物种间高度保守的重复基序,其编码基因主要在飞蝗体壁中高表达,暗示其参与体壁的形成。表达具有时期特异性,其表达模式与内表皮形成时期一致。基于RNAi的生物学功能分析显示该基因在飞蝗4龄和5龄若虫期均能被有效沉默,虽无致死表型,但影响内表皮的形成。体内20E诱导及其受体基因RNAi结果表明,受介导的20E信号通路调控。
[1] Vincent J F, Wegst U G. Design and mechanical properties of insect cuticle., 2004, 33(3): 187-199.
[2] Noh M Y, Muthukrishnan S, Kramer K J, Arakane Y. Development and ultrastructure of the rigid dorsal and flexible ventral cuticles of the elytron of the red flour beetle,., 2017, 91: 21-33.
[3] Moussian B, Seifarth C, Muller U, Berger J, Schwarz H. Cuticle differentiation duringembryogenesis., 2006, 35(3): 137-152.
[4] Andersen S O. Studies on proteins in post-ecdysial nymphal cuticle of locust,, and cockroach., 2000, 30: 569-577.
[5] Behr M, Hoch M. Identification of the novel evolutionary conservedmultigene family in invertebrates., 2005, 579(30): 6827-6833.
[6] Jasrapuria S, Specht C A, Kramer K J, Beeman R W, Muthukrishnan S. Gene families of cuticular proteins analogous to peritrophins (CPAPs) inhave diverse functions., 2012, 7(11): e49844.
[7] Petkau G, Wingen C, Jussen L C, Radtke T, Behr M. Obstructor-A is required for epithelial extracellular matrix dynamics, exoskeleton function, and tubulogenesis., 2012, 287(25): 21396-21405.
[8] Pesch Y Y, Riedel D, Behr M. Obstructor A organizes matrix assembly at the apical cell surface to promote enzymatic cuticle maturation in., 2015, 290(16): 10071-10082.
[9] Guan X, Middlebrooks B W, Alexander S, Wasserman S A. Mutation of Tweedle D, a member of an unconventional cuticle protein family, alters body shape in., 2006, 103(45): 16794-16799.
[10] Andersen S O, Rafn K, Roepstorff P. Sequence studies of proteins from larval and pupal cuticle of the yellow meal worm,., 1997, 27(2): 121-131.
[11] Togawa T, Augustine Dunn W, Emmons A C, Willis J H. CPF and CPFL, two related gene families encoding cuticular proteins ofe and other insects., 2007, 37(7): 675-688.
[12] Jensen U G, Rothmann A, Skou L, Andersen S O, Roepstorff P, HØjrup P. Cuticular proteins from the giant cockroach,., 1997, 27(2): 109-120.
[13] Charles J P. The regulation of expression of insect cuticle protein genes., 2010, 40(3): 205-213.
[14] Murata T, Kageyama Y, Hirose S, Ueda H. Regulation of thegene by FTZ-F1 during metamorphosis in., 1996, 16(11): 6509-6515.
[15] Rebers J E, Niu J, Riddiford L M. Structure and spatial expression of theMSCP14.6 cuticle gene., 1997, 27(3): 229-240.
[16] Nita M, Wang H B, Zhong Y S, Mita K, Iwanaga M, Kawasaki H. Analysis of ecdysone-pulse responsive region ofin wing disc of., 2009, 153(1): 101-108.
[17] Wang H B, Iwanaga M, Kawasaki H. Activation ofpromoter and regulation by BR-C Z2 in wing disc of., 2009, 39(9): 615-623.
[18] Wang H B, Nita M, Iwanaga M, Kawasaki H.FTZ-F1 and broad-complex positively regulate the transcription of the wing cuticle protein gene,, in wing discs of., 2009, 39(9): 624-633.
[19] Deng H, Zheng S, Yang X, Liu L, Feng Q. Transcription factors BmPOUM2 and BmFTZ-F1 are involved in regulation of the expression of the wing cuticle protein genein the silkworm,., 2011, 20(1): 45-60.
[20] Deng H, Zhang J, Li Y, Zheng S, Liu L, Huang L, Xu W H, Palli S R, Feng Q. Homeodomain POU and Abd-A proteins regulate the transcription of pupal genes during metamorphosis of the silkworm,., 2012, 109(31): 12598-12603.
[21] Zhao X, Gou X, Qin Z, Li D, Wang Y, Ma E, Li S, Zhang J. Identification and expression of cuticular protein genes based ontranscriptome., 2017, 7: 45462.
[22] 赵小明, 贾盼, 勾昕, 刘卫敏, 马恩波, 张建珍. 飞蝗内表皮蛋白基因的表达与功能分析. 中国农业科学, 2017, 50(10): 1817-1826.
ZHAO X M, JIA P, GOU X, LIU W M, MA E B, ZHANG J Z. Expression and functional analysis of endocuticle structural glycoprotein genein., 2017, 50(10): 1817-1826. (in Chinese)
[23] 王燕, 李大琪, 刘晓健, 李涛, 马恩波, 范仁俊, 张建珍. 飞蝗表皮蛋白家族基因的分子特性及基于RNAi的功能分析. 中国农业科学, 2015, 48(1): 73-82.
WANG Y, LI D Q, LIU X J, LI T, MA E B, FAN R J, ZHANG J Z. Molecular characterization and RNAi-based functional analysis offamily genes in., 2015, 48(1): 73-82. (in Chinese)
[24] Song T Q, Yang M L, Wang Y L, Liu Q, Wang H M, Zhang J, Li T. Cuticular protein LmTwdl1 is involved in molt development of the migratory locust., 2016, 23(4): 520-530.
[25] Wang X, Fang X, Yang P, Jiang X, Jiang F, Zhao D, Li B, Cui F, Wei J, Ma C, Wang Y, He J, Luo Y, Wang Z, Guo X, Guo W, Wang X, Zhang Y, Yang M, Hao S, Chen B, Ma Z, Yu D, Xiong Z, Zhu Y, Fan D, Han L, Wang B, Chen Y, Wang J, Yang L, Zhao W, Feng Y, Chen G, Lian J, Li Q, Huang Z, Yao X, Lv N, Zhang G, Li Y, Wang J, Wang J, Zhu B, Kang L. The locust genome provides insight into swarm formation and long-distance flight., 2014, 5: 2957.
[26] Liu X, Zhang H, Li S, Zhu K Y, Ma E, Zhang J. Characterization of a midgut-specific chitin synthase gene () responsible for biosynthesis of chitin of peritrophic matrix in., 2012, 42: 902-910.
[27] Yang Q, Li Z, Cao J, Zhang S, Zhang H, Wu X, Zhang Q, Liu X. Selection and assessment of reference genes for quantitative PCR normalization in migratory locust(Orthoptera: Acrididae)., 2014, 9(6): e98164.
[28] Han P, Han J, Fan J, Zhang M, Ma E, Li S, Fan R, Zhang J. 20-Hydroxyecdysone activatesmediated immune response in., 2017, 72: 128-139.
[29] Li D, Zhang J, Wang Y, Liu X, Ma E, Sun Y, Li S, Zhu K Y, Zhang J. Two chitinase 5 genes from: Molecular characteristics and functional differentiation., 2015, 58: 46-54.
[30] Liu X J, Sun Y W, Li D Q, Li S, Ma E B, Zhang J Z. Identification ofas a 20-hydroxyecdysone response gene in the chitin biosynthesis pathway from the migratory locust,., 2016, DOI 10.1111/1744-7917. 12406.
[31] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCtmethod., 2001, 25: 402-408.
[32] Xiong G, Tong X, Gai T, Li C, Qiao L, Monteiro A, Hu H, Han M, Ding X, Wu S, Xiang Z, Lu C, Dai F. Body shape and coloration of silkworm larvae are influenced by a novel cuticular protein., 2017, 207(3): 1053-1066.
[33] Apple R T, Fristrom J W. 20-Hydroxyecdysone is required for, and negatively regulates, transcription ofpupal cuticle protein genes., 1991, 146(2): 569-582.
[34] Hiruma K, Hardie J, Riddiford L M. Hormonal regulation of epidermal metamorphosis: control of expression of a larval-specific cuticle gene., 1991, 144(2): 369-378.
(责任编辑 岳梅)
Molecular characteristics and Function Analysis of Cuticle Protein Gene
YANG YaTing1,2, ZHAO XiaoMing1,QIN ZhongYu1,2, LIU WeiMin1, MA EnBo1, ZHANG JianZhen1
(1Research Institute of Applied Biology, Shanxi University, Taiyuan 030006;2College of Life Science, Shanxi University, Taiyuan 030006)
【】Theobjective of this study is to search and clone afrom() based on transcriptome database, analyze its sequence and expression characteristics. Then its expression was determined after 20-hydroxyecdysone (20E) induction and interference 20E receptorin individuals by RNAi, respectively. Meanwhile, the function ofwas analyzed by H&E staining based on RNAi with ds. The results will provide a theoretical basis for the pest control.【】A cuticle protein gene was obtained according to the transcriptome database of locusts, and the length of the cDNA sequence was cloned by reverse-transcription PCR (RT-PCR) and sequenced. The gene structure and sequence characteristics were analyzed by using the genome sequence of the locusts. Using the MEGA 6.0 software, the neighbor-joining (NJ) method was used to construct evolutionary tree with the homologous sequences of NCP1 from other insects. Expression profiles ofat different developmental days and in different tissues at day 2 of 5th instar nymphs were assayed using reverse-transcription quantitative PCR (RT-qPCR).Using RT-qPCR, the expression ofwas detected after treated with 20Efor 1, 3, 6, 12 h and interfering with 20E receptor geneby RNAi for 48 and 72 h, respectively. The biological function ofwas analyzed by combination of RNAi and H&E staining method. 【】 A cuticle protein gene was obtained by searching the transcriptome database of. The length of the cDNA is 457 bp and full length of its ORF is 306 bp, encoding 101 amino acids. Amino acid sequence analysis showed that the protein encoded by the gene contains one signal peptide and three repeated motifs, but no chitin binding domain, and the repeating motifs were conserved among species by Weblogo analysis. Phylogenetic tree analysis showed that the protein has a close genetic relationship with BcNCP1 of. Thus, the protein was named as LmNCP1 (GenBank accession number: MF326211) according to the result of phylogenetic tree. The results of RT-qPCR showed thatwas highly expressed in integument at day 2 of 5th instar nymphs, and lower expression in wing pads, foregut and fat body, but low or no expression in other tested tissues. The expression ofwas high at the early stages (N5D1 and N5D2) of 5th instar nymphs, and then decreased (N5D3-N5D6), following a increase at before ecdysis of next instar (N5D7), which is coincidence with the formation of endocuticle and exocuticle. Compared with the control group, the expression ofsignificantly increased by 1.3 and 0.9 times after injection with 20E for 1 h and 3 h. After 48 h and 72 h ofinterference by RNAi, the expression ofsignificantly decreased by 71% and 87%, respectively, comparedwiththe control group. After declining the expression ofby RNAi at day 2 of 4th and 5th instar nymphs, the locusts could normally molt and no obvious phenotype was observed. However, the endocuticle of the locusts was thinner than that of the control by H&E staining.【】 A cuticle protein genewas obtained according to the transcriptome database of. The protein encoded bydoes not contain chitin binding domain, but contains three repeated motifs.was mainly expressed in integument among all the tested tissues. Furthermore,responds to the regulation of-mediated 20E signaling pathway. The results of RNAi suggested thatwas involved in the formation of endocuticle duringmolting.
; cuticle protein;; 20-hydroxyecdysone (20E)
2017-10-09;
2018-01-05
国家自然科学基金(31702067,31640075,31672364)、山西省青年科学基金(201601D021102)
杨亚亭,Tel:0351-7018871;E-mail:2291272205@qq.com。
赵小明,Tel:0351-7018871;E-mail:zxming@sxu.edu.cn。通信作者张建珍,Tel:0351-7018871;E-mail:zjz@sxu.edu.cn
