套作对大豆苗期茎秆纤维素合成相关糖类物质转化的影响及其与叶片光合的关系
2018-04-18任胜茂邓榆川文凤君SajadHussain蒲全明于晓波刘卫国杨文钰
任胜茂,邓榆川,文凤君,Sajad Hussain,蒲全明,于晓波,刘卫国,杨文钰
套作对大豆苗期茎秆纤维素合成相关糖类物质转化的影响及其与叶片光合的关系
任胜茂1, 2,邓榆川1, 2,文凤君2,Sajad Hussain1,蒲全明2,于晓波2,刘卫国1,杨文钰1
(1四川农业大学农学院/农业部西南作物生理生态与耕作重点实验室,成都 611130;2南充市农业科学院,四川南充 637000)
【】从纤维素合成相关糖类物质转化的角度,阐明玉米大豆套作模式下,大豆苗期茎秆光形态建成的机理。【】在大豆单作和玉米大豆套作两种种植模式下,以强耐荫大豆南豆12和弱耐荫大豆南032-4为试验材料,对叶片光合速率和茎秆总碳、纤维素、可溶性糖、蔗糖、β-1,3-葡聚糖等含量进行测定和分析。【】与单作相比,套作大豆由于受到玉米荫蔽,苗期光合速率显著降低,但材料间对套作的反应程度不同,强耐荫大豆南豆12受套作荫蔽的影响程度相对较小,在套作下表现出较强的光合能力;套作显著降低了大豆叶片和茎秆的总碳含量,但南豆12的降低幅度显著低于南032-4。相关分析表明叶片光合速率与叶片和茎秆的总碳含量、茎秆的纤维素含量均呈极显著正相关(=0.952,0.935,0.825,<0.01),说明荫蔽通过影响大豆叶片的光合速率,减少了光合产物的积累和向茎秆中的分配,导致大豆植株茎秆纤维素含量降低;强耐荫大豆南豆12在荫蔽下的光合速率较高,光合产物积累较多,适合套作种植。在整个苗期,虽然套作大豆茎秆可溶性糖含量均显著低于单作,但β-1,3-葡聚糖和蔗糖含量在大豆出苗后30—51 d却表现为套作显著高于单作,且在套作模式下,两个大豆糖类物质转化率差异显著;同一种植模式下,强耐荫大豆南豆12茎秆可溶性糖、蔗糖和β-1,3-葡聚糖的含量和转化率均显著或极显著高于南032-4。对纤维素沉积方式分析表明,同一大豆材料,单作模式下茎秆纤维素快速累积时间和累积速率要高于套作;同一种植模式下,强耐荫大豆南豆12纤维素快速累积时间要短于南032-4,但差异较小,而累积速率要高于南032-4,最终导致南豆12的茎秆纤维素含量显著高于南032-4。【】套作荫蔽降低了大豆叶片的光合能力,减少了光合产物向茎秆的运输量和茎秆填充物的含量,改变了茎秆纤维素的沉积方式,使得纤维素的含量降低;而强耐荫性大豆南豆12在套作模式下能保持较高光合能力和茎秆纤维素合成能力,具有较强的抗倒伏性。
套作大豆;光合速率;总碳;纤维素;可溶性糖;蔗糖;β-1,3-葡聚糖
0 引言
【研究意义】光除了作为一种能源参与光合作用外,它还以环境信号的形式通过光敏色素等作用途径调节植物生长、发育和形态建成[1]。当植株处于弱光环境中时,主要通过改变形态结构和调整生物量积累分配来适应环境[2-3]。大豆是喜光作物,在整个生育期内对光照的反应都很敏感[4],当大豆处于弱光环境时,会降低光合产物的积累并重新分配,使得茎秆变细,节间过度伸长,增加了倒伏发生的风险[5]。在中国大豆南方种植区,普遍采用玉米大豆带状套作种植[6],在该模式下,大豆苗期与玉米共生,受玉米遮荫而处于弱光照强度和低R﹕FR的光照环境,加之受到玉米对其水分、养分的竞争,导致植株光合能力减弱,生物量积累降低,植株茎秆形态结构发生改变,在初花期前极易发生藤蔓化和倒伏[5, 7-10],这不仅造成大豆严重减产,还导致籽粒品质下降,严重地制约着中国南方大豆产业的可持续发展[6]。因此,以西南地区玉米大豆带状套作种植模式为对象,研究该模式下荫蔽对大豆叶片光合产物积累转移和茎秆理化性质的影响,可为优化茎秆形态,提高植株抗倒性提供重要的理论支持。【前人研究进展】荫蔽会减弱大豆的叶片光合能力,降低光合产物的积累,提高光合产物向茎秆中的分配比例,促进茎秆的伸长生长,从而改变植株的形态建成[5, 11]。而茎秆作为套作大豆形态建成的关键部位,对维持植株较优的形态结构起着重要的作用[12]。研究表明,茎秆中纤维素的含量对提高植株茎秆机械强度和维持较优的茎秆形态具有明显的促进作用[13-14],而光照对套作大豆茎秆中纤维素的合成起着重要的调控作用[15-16]。套作模式下,大豆冠层光合有效辐射降低,茎秆长粗比增大,纤维素含量降低,机械强度变弱,极易造成倒伏[13-14]。在对小麦、水稻、玉米等作物的研究中也发现茎秆中纤维素含量越高品种的抗倒性也越强[17-19]。【本研究切入点】前人对大豆光合产物积累分配、茎秆形态建成和理化性质的研究都基于单作条件,且集中于施肥调节方面。而对不同栽培模式下大豆苗期茎秆理化性质差异形成的原因却鲜有报道。【拟解决的关键问题】本文通过研究套作大豆苗期茎秆纤维素代谢与叶片光合的关系,分析不同耐荫性的两个大豆材料在单作和套作种植模式下的叶片光合特征、光合产物的转化特性和茎秆生理特征的差异情况,为套作大豆高产栽培和抗倒品种选育提供理论依据。
1 材料与方法
1.1 试验地点和材料
试验于2014年和2015年在四川农业大学雅安校区教学农场进行。试验地为重壤地,有机质29.86 g·kg-1、全氮0.72 g·kg-1、速效氮114.88 mg·kg-1、碱解氮62.29 mg·kg-1、速效磷36.85 mg·kg-1、速效钾136.61 mg·kg-1,pH 7.4。供试大豆材料为弱耐荫的南032-4和强耐荫的南豆12[7]。玉米品种为正红505。
1.2 试验设计
试验采用裂区设计,主区为玉米-大豆带状套作和大豆单作2种种植模式,副区为2个大豆材料,试验重复3次。套作模式中,按照《玉米大豆带状复合种植技术规程(NY/T 2632-2014)》采用宽窄行种植(图1),带宽2 m,宽行1.6 m,窄行0.4 m,玉米种植于窄行,大豆种植于宽行,行距均为0.4 m,每带种2行玉米和2行大豆,每个小区种植6带,带长12 m,小区面积为72 m2;单作模式中,大豆采用等行距种植,行距为0.5 m,长6 m,每小区种植6行,小区面积为18 m2。
玉米分别于2014年4月5日和2015年4月2日育苗,7 d后移栽,穴距0.2 m,每穴1株;大豆分别于2014年6月12日和2015年6月18日播种于玉米宽行内,穴距0.1 m,与玉米行相隔0.6 m,每穴定苗1株。施肥和其他管理措施同大田生产。

实线和虚线分别代表玉米行和大豆行,黑色实心圆和灰色实心圆分别代表单株玉米和大豆,(a)和(b)分别代表套作和单作配置
1.3 光合特性的测定方法
大豆出苗后23 d开始,每隔7 d测定一次叶片光合速率直至玉米大豆共生期结束。在晴朗无风的上午11:00—12:00,按小区选取长势一致的植株5株,利用光合系统测定仪LI-6000(LI-COR,USA)测定第3片复叶的光合速率(Photosynthetic rate,n)。
1.4 植株总碳、茎秆纤维素及其合成相关糖类物质测定
大豆苗期每个重复选取长势一致的植株15株,迅速截取茎秆第2节间和第3片复叶分别装入纸袋,在105℃下杀青0.5 h,并于70℃条件下烘干至恒重后粉碎,过60目筛用于测定以下指标:
用CE-440型元素分析仪测定总碳含量;用蒽酮比色法测定可溶性糖含量[15];用间苯二酚法测定蔗糖含量[15];用分光光度法测定β-1,3-葡聚糖含量[20];用瑞典产FOSS 1020型半自动纤维分析仪测定纤维素含量,均重复测定3次。
1.5 数据统计分析
用Microsoft Excel 2007软件整理数据,用origin 9.0绘图,用SPSS 17.0软件分析数据,采用Duncan氏新复极差法(SSR)进行显著性检验。
2 结果
2.1 套作对大豆苗期光合速率的影响
由图2可知,随着生育进程的不断推进,大豆叶片光合速率不断增加,变化趋势在材料、种植模式和年际间均表现一致。套作使大豆各时期光合速率均显著降低,但受影响的程度因大豆材料而异。与单作相比,套作模式下南032-4光合速率降低34.10%,而南豆12仅降低25.27%(2年不同测定时期平均值)。方差分析表明种植模式(=0.003)和材料(=0.001)均极显著影响大豆叶片光合速率,且互作效应显著(=0.006)。说明强耐荫大豆南豆12受套作荫蔽的影响程度相对较小,在套作下表现出较强的光合能力。

图2 不同种植模式对大豆叶片光合速率的影响
2.2 套作对大豆苗期叶片和茎秆总碳、茎秆纤维素及其合成相关糖类物质的影响
2.2.1 叶片和茎秆总碳 由图3可知,随着生育进程的不断推进,大豆叶片和茎秆总碳含量不断增加,变化趋势在材料、种植模式和年际间均表现一致。2种大豆材料叶片和茎秆总碳差异主要表现在总碳的含量和降低幅度,套作使大豆各时期叶片和茎秆总碳含量均显著降低,但受影响的程度因大豆材料而异。与单作相比,套作模式下南032-4叶片和茎秆总碳含量分别降低5.84%、5.55%,而南豆12降低了3.88%、4.11%(2年不同测定时期平均值)。方差分析表明种植模式(=0.000)和材料(=0.000)均极显著影响大豆茎秆总碳,且互作效应显著(=0.043)。说明套作会降低大豆碳素化合物的积累,而强耐荫大豆南豆12具有较高的总碳含量,表现出较强的碳素同化能力和转移能力,是大豆较优形态建成的物质基础。
2.2.2 茎秆可溶性糖 从图4可看出,随着生育进程的不断推进,大豆茎秆可溶性糖含量不断降低,变化趋势在材料、种植模式和年际间均表现一致。2种大豆材料茎秆可溶性糖差异主要表现在可溶性糖的含量和转化率(各生育期茎秆可溶性糖含量较大豆出苗23 d茎秆中减少的量占大豆出苗23 d时含量的百分比[21])。套作使大豆各时期茎秆可溶性糖含量均显著降低,但受影响的程度因大豆材料而异,与单作相比,套作模式下南032-4茎秆可溶性糖含量降低5.84%,而南豆12仅降低3.88%(2年不同测定时期平均值),不同种植模式间差异极显著(=0.008)。而同一种植模式下,材料间茎秆可溶性糖含量均表现为南豆12显著高于南032-4。在共生期结束时,单作模式下南豆12和南032-4的转化率分别为49.09%和42.80%,套作模式为44.35%和40.80%,种植模式间差异也极显著(=0.033)。
2.2.3 蔗糖 由图5可知,单作模式下大豆茎杆中蔗糖的含量随着大豆生育时期的推进而不断增加,峰值出现在大豆出苗后58 d,材料间表现为南豆12显著高于南032-4;套作模式下大豆茎秆中蔗糖含量呈“升-降-升”的变化趋势,峰值出现在大豆出苗后44 d和51 d。大豆出苗后44 d前,由于玉米的遮光效果严重,大豆叶片通过光合作用积累的光合产物大部分用于植株的茎秆生长,在茎秆中积累较多的蔗糖。44 d后,随着玉米成熟,遮荫程度逐渐减弱,光合产物供应茎秆生长的相对量减少,而蔗糖又迅速转化大豆茎秆中的纤维素,蔗糖的含量降低。51 d后,由于玉米遮荫程度的降低,大豆叶片的光合速率迅速增加,茎秆蔗糖来源充足。同一种植模式下,两大豆材料茎秆蔗糖含量均为南豆12高于南032-4。
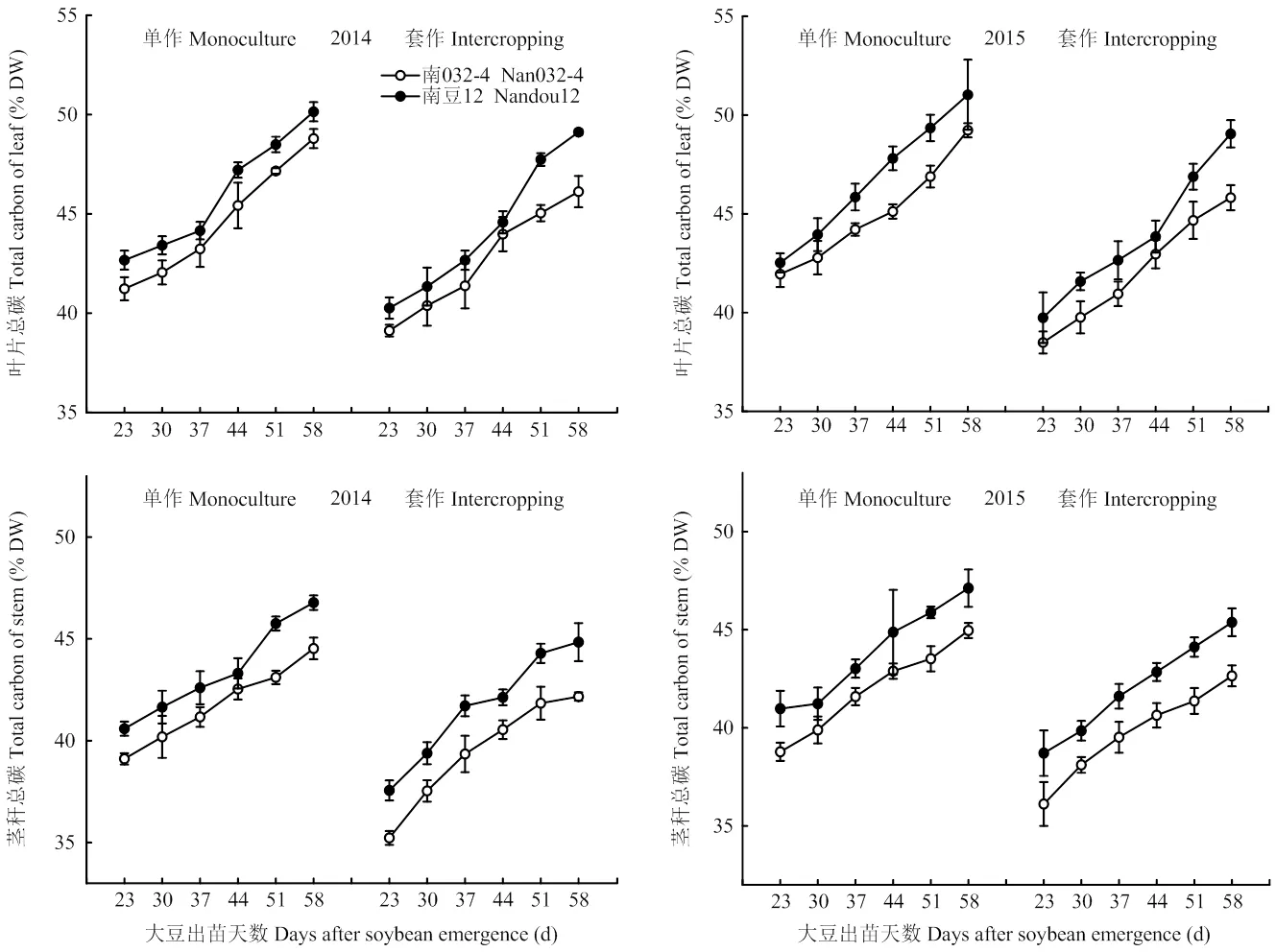
图3 不同种植模式对大豆茎秆总碳的影响

图4 不同种植模式对大豆茎秆可溶性糖的影响
。

图5 不同种植模式对大豆茎秆蔗糖的影响
2.2.4 β-1,3-葡聚糖 β-1,3-葡聚糖的沉积和降解对纤维素的合成具有一定的调控和促进作用[22],从图6可看出,随着生育进程的不断推进,大豆茎秆β-1,3-葡聚糖含量不断降低,变化趋势在材料、种植模式和年际间均表现一致。2种大豆材料茎秆β-1,3-葡聚糖差异主要表现在β-1,3-葡聚糖的含量及其转化率。同一种植模式下,大豆茎秆β-1,3-葡聚糖含量均表现为南豆12显著高于南032-4。同一大豆材料,在出苗后30—51 d表现为套作显著高于单作,其余生育时期表现为套作低于单作;大豆出苗44 d前,单作大豆茎秆β-1,3-葡聚糖的转化率显著高于套作,44 d后表现为套作显著高于单作,而共生期结束时,套作模式下南豆12的转化率为62.04%;南032-4为59.03%,种植模式间差异极显著(=0.041)。
2.2.5 纤维素 由图7可知,随着生育进程的不断推进,大豆茎秆纤维素含量不断增加,变化趋势在材料、种植模式和年际间均表现一致。套作使大豆各时期茎秆纤维素含量均显著降低,但受影响的程度因大豆材料而异。与单作相比,套作模式下南032-4茎秆纤维素含量降低9.55%,而南豆12仅降低6.78%(2年不同测定时期平均值)。方差分析表明,种植模式(=0.014)和材料(=0.027)均显著影响茎秆纤维素含量,且互作效应显著(=0.007),强耐荫大豆南豆12在套作环境下能保持较高水平的茎秆纤维素含量,表现出较优的茎秆力学性能。

图6 不同种植模式对大豆茎秆β-1, 3-葡聚糖的影响
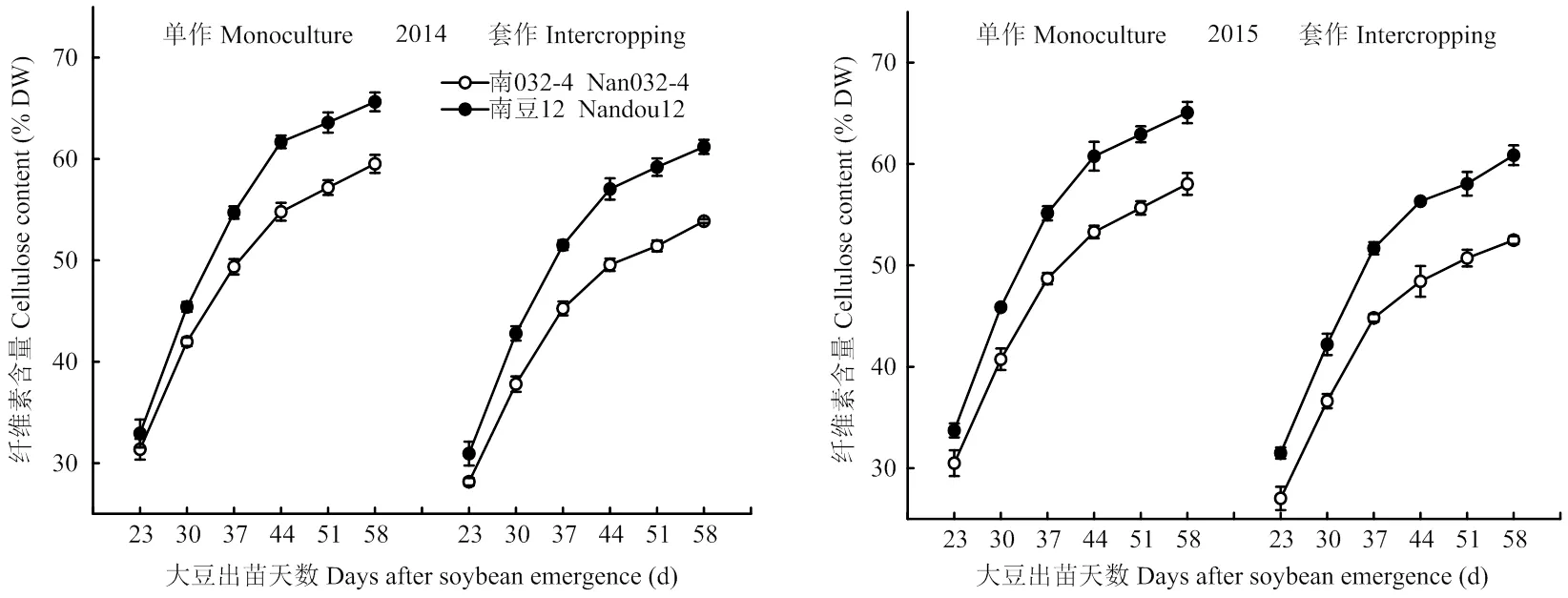
图7 不同种植模式对大豆茎秆纤维素的影响
用Logistic模型[23]对纤维素的累积进行模拟,回归系数均达极显著水平(表1)。进一步分析表中的特征值发现,同一种植模式下2种大豆材料茎秆纤维素沉积特性的差异主要表现在纤维素快速累积时间的长短和累积速率的高低上。2年试验结果均表现为强耐荫大豆南豆12茎秆纤维素快速累积时间较弱耐荫大豆南032-4开始较晚,持续时间较短,但累积速率较大,表现出较快的纤维素积累过程;不同种植模式对2种大豆材料茎秆纤维素沉积特性的差异主要表现在累积速率上,单作模式下2种大豆材料茎秆纤维素的累积速率高于套作。说明在套作模式下强耐荫大豆南豆12具有较强的光合能力和物质转化能力,能在相对较短的时间内将茎秆中的光合产物转化成细胞壁结构物质——纤维素。

表1 不同种植模式对大豆茎秆纤维素积累特征的影响
T、t1、t2、Vmax分别表示纤维素快速累积时间、纤维素快速累积起始时间、纤维素快速积累截止时间、纤维素累积速率;*,<0.05;**,<0.01。下同
T, t1, t2and Vmaxrepresent the rapid accumulation time of cellulose, initial time of rapid accumulation of cellulose, rapid accumulation of cellulose, and the accumulation rate of cellulose; *,<0.05; **,<0.01. The same as below
2.3 纤维素累积特征值与纤维素合成相关物质转化率的相关性
纤维素累积特征值中与纤维素合成相关性最为密切的是纤维素快速累积时间和纤维素累积速率[24]。相关分析表明(表2),大豆茎秆纤维素累积特征值与相关物质转化率的相关系数存在着大小和正负的差异。综合2年的数据表明,大豆茎秆纤维素快速累积时间、累积速率与相关物质转化率的相关性均达到显著或极显著水平(除2015年单作模式下纤维素快速累积时间与可溶性糖转化率相关性不显著外),前者与物质转化率呈极显著正相关,而后者与物质转化率呈显著负相关。大豆茎秆纤维素合成相关物质转化率越高,纤维素快速累积时间越短,累积速率增加,越有利于高强纤维的形成。
2.4 大豆茎秆纤维素及其合成相关物质与叶片光合的相关分析
综合2年的数据,相关分析表明(表3),大豆叶片光合速率与叶片总碳、茎秆总碳、茎秆纤维素含量均呈极显著正相关。说明大豆苗期叶片光合能力与茎秆纤维素含量密切相关,光合能力越强,积累的碳素同化物也越多,而强耐荫大豆南豆12还表现出较强的茎秆光合产物转移能力,为纤维素的合成提供了必要的能量基础。纤维素含量与蔗糖含量呈极显著正相关,蔗糖不仅作为光合产物的运输形式也作为茎秆纤维素合成的直接底物,而较强的蔗糖转移能力是提高套作大豆茎秆纤维素含量,优化植株形态建成的物质基础;与可溶性糖和β-1,3-葡聚糖呈极显著负相关,可溶性糖和β-1,3-葡聚糖纤维素合成的间接底物,共同作用调节纤维素的合成。
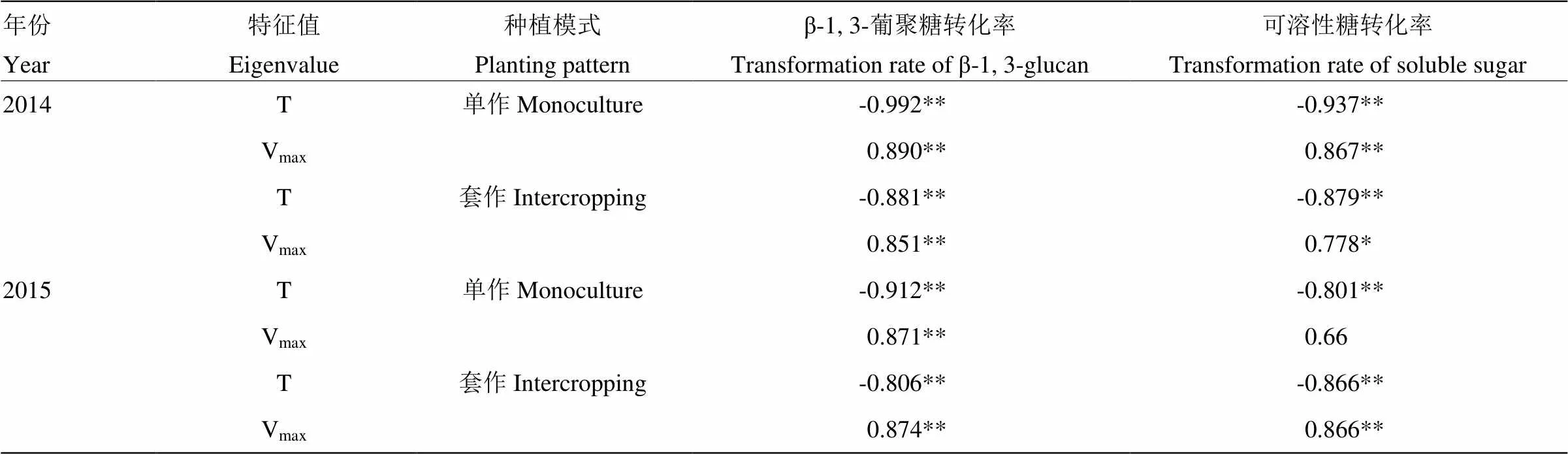
表2 不同种植模式对大豆茎秆纤维素累积特征值与物质转化率的相关分析

表3 纤维素含量及相关糖类物质与叶片光合的相关性分析
3 讨论
3.1 套作对大豆苗期叶片光合的影响
刘卫国等[7, 9]、王竹等[10]研究表明,在玉米大豆带状套作种植模式中,玉米的遮荫和对自然光谱的选择性吸收,降低了大豆冠层光合有效辐射强度和红光与远红光的比例,使大豆容易倒伏。通过对不同种植模式下大豆叶片光合速率的测定分析发现,套作模式下,大豆叶片光合速率降低,而在弱光环境中导致光合速率降低的主要原因除能量不足外,还与叶片气孔导度变小有关。虽然在弱光环境中大豆叶片胞间较高浓度的二氧化碳保证了光合作用的供应,有利于植物光合作用,但气孔导度小却限制了二氧化碳的扩散,从而使光合速率降低[5]。研究表明,大豆对荫蔽的响应与不同基因型植株叶片光合特性和光合产物积累分配代谢等密切相关[5, 25-26]。本试验进一步研究发现,大豆苗期叶片光合速率与叶片总碳和茎秆总碳呈极显著正相关(=0.952,0.935,=0.005)。荫蔽下倒伏发生严重可能是大豆受弱光胁迫,改变了光合产物的积累分配,生理代谢受阻,影响包括茎秆在内的光合形态建成[5]。虽然本文中2种大豆参试材料的光合速率均降低,但强耐荫大豆材料南豆12降低程度较南032-4低,表现出较强的光合与抗倒性特性,适合套作种植。
3.2 套作对大豆苗期茎秆纤维素合成代谢的影响
茎秆中可溶性糖的转化是纤维素合成的前提和物质基础[26],其中蔗糖降解产生的UDPG是纤维素合成的直接底物[27-28],β-1,3-葡聚糖的沉积和降解对微纤丝的结晶和纤维素的合成具有调控和促进作用[29],因此茎秆中可溶性糖含量及其转化对纤维素累积具有重要影响。本试验研究表明,随着生育时期的不断推进,大豆茎秆可溶性糖转化率逐渐增加,含量逐渐降低,并受光环境和基因型的共同作用。强耐荫性大豆材料具有较强的可溶性糖转化率,这是其茎秆抗倒形态建成的基础。蔗糖作为纤维素合成的直接底物,其变化情况对纤维素合成的影响最为直接[28]。本试验与Liu等[16]的研究结果相一致,蔗糖含量越多,纤维素累积量越大,荫蔽环境促进茎秆生长的同时,增加了光合产物向茎秆的转移量,从而维持蔗糖的稳定增长,保证纤维素的合成。当茎秆细胞进入次生壁加厚发育期时,除纤维素开始快速累积外,细胞壁内侧还合成大量的非纤维素物质β-1,3-葡聚糖[30],β-1,3-葡聚糖主要是对纤维素的合成起调控和促进作用。本试验表明,大豆出苗44 d前,单作大豆茎秆β-1,3-葡聚糖的转化率显著高于套作,44 d后表现为套作显著高于单作。而强耐荫大豆材料具有较强的β-1,3-葡聚糖转化率,相关分析表明,β-1,3-葡聚糖含量与纤维素的含量和叶片光合速率呈极显著负相关(=-0.863,-0.811,= 0.004),说明β-1,3-葡聚糖是茎秆纤维素合成的重要代谢物质之一,对套作大豆茎秆纤维素合成起着重要的调控作用。
3.3 套作对大豆苗期茎秆纤维素累积的影响
纤维素累积方式是影响纤维素积累量的最直接原因[24]。张文静等[22]研究表明,纤维素快速累积时间和累积速率与纤维含量的关系必须在一定条件下才成立,而并非纤维素快速累积历时长、累积速率越平缓,纤维含量就越高。本试验中,2年的数据表明,不同种植模式下,单作大豆茎秆纤维素快速累积时间和累积速率要高于套作大豆,强耐荫大豆南豆12纤维素快速累积时间要长于弱耐荫大豆南032-4,但差异较小,而累积速率要高于后者,最终表现为南豆12的茎秆纤维素含量显著高于南032-4。说明荫蔽会改变纤维素的积累方式,降低纤维素的最大累积时间和累积速率,而强耐荫性大豆材料具有较强的荫蔽适应能力,在相差较小的纤维素最大累积时间下具有较高的积累速率,表现出较强的纤维素合成能力。
4 结论
在玉米大豆带状套作模式下,荫蔽导致大豆叶片光合速率、叶片和茎秆中总碳含量降低,茎秆纤维素含量和与纤维素合成相关的可溶性糖、蔗糖、β-1,3-葡聚糖含量降低,可溶性糖、β-1,3-葡聚糖的转化率也降低,但不同材料对荫蔽的反应程度不同。与单作相比,套作模式下强耐荫性材料光合能力较强,积累的碳素同化物较多,茎秆中的可溶性糖、蔗糖、β-1,3-葡聚糖含量较高,运输和转化能力也较强,纤维素含量高,为减少苗期倒伏创造了条件。
[1] 吴能表, 谈锋, 肖文娟, 王小佳. 光强因子对少花桂幼苗形态和生理指标及精油含量的影响. 生态学报, 2005, 25(5): 1159-1164.
Wu N B, Tan F, Xiao W J, Wang X J. Effects of light intensity on morphologic and physiological indexes and safrol content of., 2005, 25(5): 1159-1164. (in Chinese)
[2] VALLADARES F, NIINEMETS Ü. Shade tolerance, a key plant feature of complex nature and consequences., 2008, 39 (1): 237-257.
[3] Matos F S, Wolfgramm R, Goncalves F V, Cavatte P C, Ventrella M C, DaMatta F M. Phenotypic plasticity in response to light in the coffee tree., 2009, 67 (2): 421-427.
[4] 韩天富, 盖钧镒, 王金陵, 周东兴. 大豆开花逆转现象的发现. 作物学报, 1998, 24(2): 168-171.
HAN T F, GAI J Y, WANG J L, ZHOU D X. Discovery of flowering reversion in soybean plants., 1998, 24(2): 168-171. (in Chinese)
[5] 刘婷, 刘卫国, 任梦露, 杜勇利, 邓榆川, 邹俊林, 方萍, 杨文钰. 遮荫程度对不同耐荫性大豆品种光合及抗倒程度的影响. 中国农业科学, 2016, 49(8): 1466-1475.
LIU T, LIU W G, REN M L, DU Y L, DENG Y C, ZHOU J L, FANG P, YANG W Y. Effects of shade degrees on photosynthesis and lodging resistance degree of different shade tolerance soybean., 2016, 49(8): 1466-1475. (in Chinese)
[6] 杨文钰, 雍太文, 任万军, 樊高琼, 牟锦毅, 卢学兰. 发展套作大豆, 振兴大豆产业. 大豆科学, 2008, 27(1): 1-7.
YANG W Y, YONG T W, REN W J, FAN G Q, MOU J Y, LU X L. Develop relay-planting soybean, revitalize soybean industry., 2008, 27(1): 1-7. (in Chinese)
[7] Liu W, Zou J, Zhang J, Yang F, Wan Y, Yang W. Evaluation of soybean () stem vining in maize-soybean relay strip intercropping system., 2015, 18(1): 69-75.
[8] 刘卫国, 宋颖, 邹俊林, 张云松, 梅林森, 蒋涛, 杨文钰. LED灯模拟作物间作套种群体内光环境的设计与应用. 农业工程学报, 2011, 27(8): 288-292.
LIU W G, SONG Y, ZOU J L, ZHANG Y S, MEI L S, JIANG T, YANG W Y. Design and effect of LED simulated illumination environment on intercropping population., 2011, 27(8): 288-292. (in Chinese)
[9] 刘卫国, 蒋涛, 佘跃辉, 杨峰, 杨文钰. 大豆苗期茎秆对荫蔽胁迫响应的生理机制初探. 中国油料作物学报, 2011, 33(2): 141-146.
LIU W G, JIANG T, SHE Y H, YANG F, YANG W Y. Preliminary study on physiological response mechanism of soybean () stem to shade stress at seedling stage., 2011, 33(2): 141-146. (in Chinese)
[10] 王竹, 杨文钰, 伍晓燕, 吴其林. 玉米株型和幅宽对套作大豆初花期形态建成及产量的影响. 应用生态学报, 2008, 19(2): 323-329.
WANG Z, YANG W Y, WU X Y, WU Q L. Effects of maize plant type and planting width on the early morphological characters and yield of relay planted soybean., 2008, 19(2): 323-329. (in Chinese)
[11] 范元芳, 杨峰, 何知舟, 王锐, 刘沁林, 袁小琴, 雍太文, 武晓玲, 杨文钰. 套作大豆形态、光合特征对玉米荫蔽及光照恢复的响应. 中国生态农业学报, 2016, 24(5): 608-617.
FAN Y F, YANG F, HE Z Z, WANG R, LIU Q L, YUAN X Q, YONG T W, WU X L, YANG W Y. Effects of shading and light recovery on soybean morphology and photosynthetic characteristics in soybean-maize intercropping system., 2016, 24(5): 608-617. (in Chinese)
[12] 任梦露, 刘卫国, 刘婷, 杜勇利, 邓榆川, 邹俊林, 袁晋, 杨文钰. 荫蔽胁迫下大豆茎秆形态建成的转录组分析. 作物学报, 2016, 42(9): 1319-1331.
REN M L, LIU W G, LIU T, DU Y L, DENG Y C, ZOU J L, YUAN J, YANG W Y. Transcriptome analysis of stem morphogenesis under shade stress in soybean., 2016, 42(9): 1319-1331. (in Chinese)
[13] 向达兵, 郭凯, 雷婷, 于晓波, 罗庆明, 杨文钰. 磷钾营养对套作大豆茎秆形态和抗倒性的影响. 中国油料作物学报, 2010, 32(3): 395-402.
XIANG D B, GUO K, LEI T, YU X B, LUO Q M, YANG W Y. Effects of phosphorus and potassium on stem characteristics and lodging resistance of relay cropping soybean., 2010, 32(3): 395-402. (in Chinese)
[14] 吴其林, 王竹, 杨文钰. 苗期遮荫对大豆茎秆形态和物质积累的影响. 大豆科学, 2007, 26(6): 868-872.
WU Q L, WANG Z, YANG W Y. Seedling shading affects morphogenesis and substance accumulation of stem in soybean., 2007, 26(6): 868-872. (in Chinese)
[15] 邓榆川, 刘卫国, 袁小琴, 袁晋, 邹俊林, 杜俊波, 杨文钰. 套作大豆苗期茎秆纤维素合成代谢与抗倒性的关系. 应用生态学报, 2016, 27(2): 469-476.
DENG Y C, LIU W G, YUAN X Q, YUAN J, ZOU J L, DU J B, YANG W Y. Relationship between cellulose synthesis metabolism and lodging resistance in intercropping soybean at seedling stage., 2016, 27(2): 469-476. (in Chinese)
[16] Liu W, Deng Y, Hussain S, Zou J, Yuan J, Luo L, Yang C, Yuan X, Yang W. Relationship between cellulose accumulation and lodging resistance in the stem of relay intercropped soybean [(L.) Merr.]., 2016, 196: 261-267.
[17] 王健, 朱锦懋, 林青青, 李晓娟, 滕年军, 李振声, 李滨, 张爱民, 林金星. 小麦茎秆结构和细胞壁化学成分对抗压强度的影响. 科学通报, 2006, 51(6): 679-685.
WANG J, ZHU J M, LIN Q Q, LI X J, TEN N J, LI Z S, LI B, ZHANG A M, LIN J X. Effect of wheat stalk structure and chemical composition of cell wall on compressive strength., 2006, 51(6): 679-685. (in Chinese)
[18] 黄海. 群体密度对玉米茎秆及根系抗倒伏特性的影响[D]. 长春: 吉林农业大学, 2013.
HUANG H. Effect of population density on lodging-resistancecharacteristic of stalk and root in maize[D]. Changchun: Jilin Agricultural University, 2013. (in Chinese)
[19] 杨世民, 谢力, 郑顺林, 李静, 袁继超. 氮肥水平和栽插密度对杂交稻茎秆理化特性与抗倒伏性的影响. 作物学报, 2009, 35(1): 93-103.
YANG S M, XIE L, ZHENG S L, LI J, YUAN J C. Effects of nitrogen rate and transplanting density on physical and chemical characteristics and lodging resistance of culms in hybrid rice., 2009, 35(1): 93-103. (in Chinese)
[20] KOHLE H, JEBLICK W, POYEN F, BLASCHEK W, KAUSS H. Chitosan-elicited callose synthesis in soybean cells as a Ca2+- dependent process., 1985, 77(3): 544-551.
[21] RUAN Y L, XU S M, WHITE R, FURBANK R T. Genotypic and developmental evidence for the role of plasmodesmatal regulation in cotton fiber elongation mediated by callose turnover., 2004, 136(4): 4104-4113.
[22] 张文静, 胡宏标, 陈兵林, 王友华, 李文峰, 周治国. 棉铃对位叶生理特性的基因型差异及其与铃重形成的关系. 棉花学报, 2007, 19(4): 296-303.
ZHANG W J, HU H B, CHEN B L, WANG Y H, LI W F, ZHOU Z G. Relationship between genotypic difference of physiological characteristics in leaf subtending boll and boll weight forming., 2007, 19(4): 296-303. (in Chinese).
[23] 束红梅, 王友华, 陈兵林, 胡宏标, 张文静, 周治国. 棉花纤维素累积特性的基因型差异及与纤维比强度形成的关系. 作物学报, 2007, 33(6): 921-926.
SHU H M, WANG Y H, CHEN B L, HU H B, ZHANG W J, ZHOU Z G. Genotypic differences in cellulose accumulation in cotton fiber and its relationship with fiber strength., 2007, 33(6): 921-926. (in Chinese)
[24] 周蓉, 王贤智, 张晓娟, 沙爱华, 吴学军, 涂赣英, 邱德珍, 周新安. 大豆种质倒伏抗性评价方法研究. 大豆科学, 2007, 26(4): 484-489.
ZHOU R, WANG X Z, ZHANG X J, SHA A H, WU X J, TU G Y, QIU D Z, ZHOU X A. Evaluation method of lodging resistance in soybean germplasm., 2007, 26(4): 484-489. (in Chinese)
[25] 黄中文, 赵团结, 喻德跃, 陈受宜, 盖钧镒. 大豆抗倒伏性的评价指标及其QTL分析. 作物学报, 2008, 34(4): 605-611.
HUANG Z W, ZHAO T J, YU D X, CHEN S Y, GAI J Y. Lodging resistance indices and related QTLs in soybean., 2008, 34(4): 605-611. (in Chinese)
[26] MEINERT M C, DELMER D P. Changes in biochemical composition of the cell wall of the cotton fiber during development., 1977, 59(6): 1088-1097.
[27] AMOR Y, HAIGLER C H, JOHNSON S, WAINSCOTT M, DELMER D P. A membrane-associated form of sucrose synthase and its potential role in synthesis of cellulose and callose in plants., 1995, 92(20): 9353-9357.
[28] Haigler C H, Datcheva M I, Hogan P S, Salnikov V V, Hwang S, Kirt M, Delmer D P. Carbon partitioning to cellulose synthesis., 2001, 47(1/2): 29-51.
[29] Ruan Y L, Xu S M, White R, FURBANK R T. Genotypic and developmental evidence for the role of plasmodesmatal regulation in cotton fiber elongation mediated by callose turnover., 2004, 136(4): 4104-4113.
[30] Scheible W R, Pauly M. Glycosyltransferases and cell wall biosynthesis: Novel players and insights., 2004, 7(3): 285-295.
(责任编辑 杨鑫浩)
Effects of Intercropping on the Transformation of Carbohydrate Related Substances in Stem of Soybean Seedling Stage and Its Relationship with Leaf Photosynthesis
REN ShengMao1, 2, DENG YuChuan1, 2, WEN FengJun2, Sajad Hussain1, Pu QuanMing2, YU XiaoBo2, LIU WeiGuo1, YANG WenYu1
(1College of Agronomy, Sichuan Agricultural University/Key Laboratory of Crop Ecophysiology and Farming System in Southwest China, Ministry of Agriculture, Chengdu 611130;2Nanchong Academy of Agriculture Sciences, Nanchong 637000, Sichuan)
【】The purpose of this study was to clarify the mechanism of photosynthetic morphology of soybean at seedling stage in maize/soybean intercropping from the perspective of transformation of related sugars and cellulose synthesis.【】Under soybean monoculture and maize/soybean intercropping systems, the strong shade tolerant soybean (Nandou12) and weak shade tolerant soybean (Nan 032-4) were used as the experimental materials. The photosynthetic rate of leaf and the total carbon, cellulose, soluble sugar, sucrose, and β-1, 3-glucan of stem were measured and analyzed. 【】Compared with soybean monoculture, the leaf photosynthetic rate of intercropping soybean was significantly decreased due to maize shading, but the degree of response was different with different soybean cultivar. In intercropping system, the decreasing degree of strong shade tolerance soybean Nandou 12 was relatively small, and Nandou 12 showed stronger photosynthetic ability than Nan 032-4. The total carbon contents in the leaf and stem of shaded soybean decreased, but the reduction degree of Nandou12 was significantly lower than that of Nan 032-4. Correlation analysis showed that the photosynthetic rate of leaf was highly significantly positive correlation with the total carbon content in the leaf and stem, and the cellulose content in the stem (=0.952, 0.935, 0.825, respectively,<0.01). The result explained that shade of intercropping affected the photosynthesis rate, reduced the accumulation and distribution of photosynthetic products in the stem. Finally, the cellulose content in soybean stem was decrease. But the strong shade tolerance soybean Nandou 12, with the higher photosynthetic rate and accumulation of photosynthetic product, was more suitable planting in intercropping. At seedling stage, the soluble sugar content of intercropping soybean stem was significantly lower than that of monoculture soybean. The contents of β-1, 3-glucan and sucrose in intercropped soybean stem from 30-51 d after germinating were significantly higher than that in monoculture, and in intercropping the carbohydrate conversion rate of two soybean was significantly different. Under the same planting pattern, the content and transformation rate of soluble sugar, sucrose and β-1,3-glucan of soybean stem with strong shade tolerance soybean Nandou 12 were significantly higher than those in Nan 032-4. The analysis of cellulose deposition modes showed that, for the same soybean material, the rapid accumulation time and accumulation rate in monoculture system were higher than that in intercropping system. In the same planting pattern, the cellulose accumulated rate in the stem of Nandou 12 was shorter than that of Nan 032-4, but the difference was small, and the accumulation rate was higher than that in Nan 032-4. It resulted in that the cellulose content in the stem of Nandou12 was significantly higher than that in Nan 032-4.【】Shading from maize in intercropping system decreases the photosynthetic capacity of soybean leaves, slow the transportation of photosynthate from leaf to stem, and reduce content of stuffing in stem stuffing. Shading changes the accumulation modes of cellulose in soybean stem, and makes the cellulose content decrease. In intercropping system, the strong shade tolerance soybean Nandou12 can maintain higher photosynthetic capacity and stronger cellulose synthesis ability in the stem, so its’ lodging resistance is strong.
intercropping soybean; photosynthetic rate; total carbon; cellulose; soluble sugar; sucrose; β-1, 3- glucan
2017-08-25;
2017-11-09
国家自然科学基金(31671626)、四川省育种攻关(2016NYZ0031)
任胜茂,E-mail:1802475325@qq.com。邓榆川,E-mail:515271439@qq.com。任胜茂和邓榆川为同等贡献作者。
刘卫国,E-mail:lwgsy@126.com。通信作者杨文钰,E-mail:mssiyangwy@sicau.edu.cn
