不同透明度水稻籽粒横断面扫描电镜分析
2018-04-09陆彦张晓敏祁琰张昌泉凌裕平刘巧泉
陆彦 张晓敏 祁琰 张昌泉 凌裕平 刘巧泉,*
不同透明度水稻籽粒横断面扫描电镜分析
陆彦1,2张晓敏2,3祁琰2,3张昌泉1凌裕平3刘巧泉1,*
(1扬州大学 农学院 植物功能基因组学教育部重点实验室/江苏省作物遗传生理重点实验室/粮食作物现代产业技术协同创新中心,江苏 扬州 225009;2扬州大学测试中心,江苏 扬州 225009;3扬州大学 园艺与植物保护学院,江苏 扬州 225009;)
【目的】探索单淀粉粒断面观察方法,并阐明稻米透明度与直链淀粉含量的关系以及造成稻米透明度下降的淀粉结构基础。【方法】采用扫描电镜对经过不同处理的具有不同透明度的籼稻和粳稻胚乳横断面进行淀粉粒结构观察研究。【结果】籼稻和粳稻稻米的透明度在直链淀粉含量低于15%的软米中明显下降。通过比较不同籽粒横断面断开方式,发现直接机械断裂法无法断开单个淀粉粒,机械断裂加玻璃刀刮刻法可以断开单个淀粉粒但断面结构变形且无法定量分析。液氮脆断法可以观察到单个断裂的淀粉粒以及其中的空腔。比较籼稻和粳稻胚乳横断面淀粉粒的排列方式和单个淀粉粒内部结构,表明所有品种稻米胚乳淀粉粒均规则且紧凑排列,但所有暗胚乳稻米籽粒横断面中淀粉粒内部存在明显的空腔且糯稻淀粉粒空腔的数目和大小均明显高于暗胚乳稻米。进一步通过梯度烘干实验,证明稻米淀粉粒中的空腔数目和大小随着水分的降低而增多和变大。【结论】液氮直接脆断法是观察水稻单个淀粉粒断面的有效方法。稻米透明度与胚乳淀粉粒的排列紧密程度无关而与稻米的含水量以及单个淀粉粒中间的气腔数目和大小直接相关。此外,直链淀粉含量越低,淀粉粒中间的空腔数目越多,孔径越大。
水稻;淀粉粒;扫描电镜;透明度;气腔
水稻是我国重要的粮食作物之一,稻米外观品质是广大消费者和研究者首要关注的性状,也是稻米商品价值的重要决定因素[1-2]。稻米外观品质主要涉及稻米粒形和透明度(或垩白),其中稻米粒形因不同地区的文化差异而存在一定的偏好性,但对稻米透明度性状的偏好基本一致,都偏爱透明度好的稻米[3-4]。目前我国优质稻米育种在常规稻和杂交稻研究方面都取得了较大进展[5],其中稻米蒸煮食味品质提升的一个重要方式是通过降低稻米表观直链淀粉含量(AAC)途径来实现。但通过该方法培育的优质软米由于AAC普遍在14%以下而造成整粒精米在存放一段时间后因水分降低或老化等原因而出现暗胚乳(或云雾状)的表型[6]。因此,目前软米一般都是真空包装销售且需要保持一定的含水量,散装销售则很快就会出现暗胚乳表型而影响消费者的选择。暗胚乳的极端表型是糯稻,精米呈乳白色,完全不透明。
有关暗胚乳和糯稻不透明性状形成的淀粉结构基础并不清楚,但可以明确它不同于常见的稻米垩白性状,后者主要是由于胚乳中区域性的淀粉粒排列松散而导致存在着一些空腔进而造成的一种光学特性[7]。早期的研究发现在糯稻以及部分暗胚乳突变体的淀粉粒表面存在空腔,但透明度是否与之有关尚不明确[8]。Zhang等[9]通过分析在糯稻基础上构建的具有不同AAC转基因稻米的透明度与胚乳淀粉粒的结构关系发现,糯稻胚乳淀粉粒中间存在很多空腔,并且随着AAC的增加,空腔数目有变少的趋势,因此认为糯稻的不透明和暗胚乳的半透明都是由于淀粉中间的空腔造成的。由于上述研究是利用转基因稻米且在一个水稻品种背景下进行比较分析的,在常规栽培的软米和糯米的水稻品种中这种现象是否存在,并不清楚。因此本研究分别在籼稻和粳稻背景下选取了具有不同AAC和透明度的稻米品种进行了稻米胚乳横断面的扫描电镜(Scanning Electron Microscopy, SEM)观察分析,一方面通过优化稻米胚乳横断面断裂方法来达到最优的观察淀粉粒断面效果;另一方面,利用最佳处理方式来明确不同稻米的透明度是否与淀粉粒中间的空腔有关系以及空腔的大小和数目是否存在差异。
1 材料与方法
1.1 实验材料
供试水稻材料包括5个籼稻品种(清芦占11、9311、滇屯502、鸭血糯、苏御糯)和5个粳稻品种(武育粳3号、日本晴、关东194、广陵香糯、太湖糯)。其中滇屯502、关东194为软米品种,鸭血糯、苏御糯、广陵香糯和太湖糯为糯稻品种。为控制生育期可能对后续试验造成干扰,上述所选粳稻品种间和籼稻品种间的生育期差异均控制在2周之内。上述材料于2016年夏季种植在扬州大学校内实验农场。此外,进一步选取了2017年夏季种植在扬州大学校内实验农场的滇屯502、苏御糯、关东194和广陵香糯进行水分含量对稻米外观和淀粉结构的分析。上述水稻材料的播种到成熟期间均实行普通大田常规栽培管理。收获的成熟水稻种子除了部分品种用于烘干处理外,其他均经过自然干燥,室温保存3个月后进行测试。
1.2 样品前处理
用于水分含量与外观分析的种子在收获后密封保存,其余分成2份,于40℃下分别烘6 h和12 h,随后用砻谷机(SY88-TH, 韩国)脱去颖壳,并用小型精米机(Kett,日本)研磨60 s,密封保存精米用于观察横断面。用水分分析仪(Mettler Toledo MJ33, 瑞士)测定含水量并拍照分析。
其他样品正常去壳、出糙和出精,用于淀粉提取和磨粉(FOSS, 瑞典)。磨完的米粉用百目筛筛除大颗粒后,转入纸袋,37℃下烘2 d,之后室温平衡2 d,用于其他分析。
1.3 稻米常规品质分析
米粉AAC的测定按照农业部颁发标准NY147-88进行,参比样品购自中国水稻研究所。稻米蛋白质含量用全自动凯氏定氮仪(Kjeltec 2300型, FOSS公司)测定,按半微量凯氏法测定米粉中全氮含量并通过换算按系数6.25计算出样品蛋白质含量,每份样品重复测定3次,取其平均值。
1.4 稻米淀粉分离
参照Zhang等[9]湿磨的淀粉分离方法,并进行一定的改动。具体为称取10 g精米于50 mL离心管中,将精米用3倍体积碱性水溶液(用NaOH调pH值至8.0~8.5)浸泡过夜后用组织匀浆机(IKA-T RCT-Basic,德国)匀浆并过300目筛。匀浆液转至100 mL三角瓶中,加入50 mg/g的碱性蛋白酶和25 μL叠氮化钠溶液(0.04 g/mL)。在42℃下,摇床消化16 h。匀浆液转移至50 mL离心管中,离心并去除上清,刮掉淀粉表层黄色部分后用蒸馏水悬浮沉淀,再次离心,重复4~5次。最后用90%无水乙醇清洗2次并用去脂抽提液(氯仿∶甲醇=1∶1),在45℃下抽提去脂30 min。沉淀随后用无水乙醇再清洗4次,然后在40℃烘箱中烘干。
1.5 稻米断面电镜观察分析
将精米进行3种处理:第一种处理是将完整精米用镊子直接掰断;第二种处理是将完整的精米放至液氮中冷冻3~5 min,然后用镊子对米粒施加外力将其脆断;第三种处理是将直接掰断的米粒断面进一步用刀片刮蹭。将3种方法处理过的精米断面朝上粘在样品台上,用SCD-500离子溅射喷镀仪镀金,在场发射扫描电子显微镜(S-4800ⅡFESEM)上观察拍照。
1.6 稻米淀粉粒的电镜观察与粒径分布测定
取10 mg烘干的淀粉粉末于1.5 mL离心管中,加入500 μL无水乙醇悬浮后,滴加到样品台上,晾干后,置于离子溅射喷镀仪(SCD-500)内喷金镀膜,用场发射扫描电子显微镜(S-4800ⅡFESEM)观察稻米淀粉粒的形貌及颗粒大小,随机选取多个观察区域,拍照。根据放大倍数校正标尺后,每个样品随机选择200粒淀粉,利用Image J软件测量其直径,随后用Excel软件计算其平均值、标准差,并绘制粒径的频数分布图。
1.7 统计分析
上述测量数据的输入和分析由Excel软件整理并由统计软件SPSS 16.0进行单因素方差分析检测不同数据间是否存显著差异。试验数据以平均值±标准差表示。
2 结果与分析
2.1 不同品种稻米的外观与基本理化指标比较
对相同存储条件下不同品种稻米的精米进行外观比较发现,籼稻中清芦占11和9311稻米均为正常的透明籽粒而软米滇屯502则为半透明的暗胚乳表型。两个籼糯品种鸭血糯和苏御糯为完全不透明的乳白色(图1)。粳稻中同样存在类似的区别,武育粳3号和日本晴都为正常的透明籽粒,软米关东194为半透明的暗胚乳外观,而两个糯稻品种广陵香糯和太湖糯稻米则为不透明的乳白色(图1)。
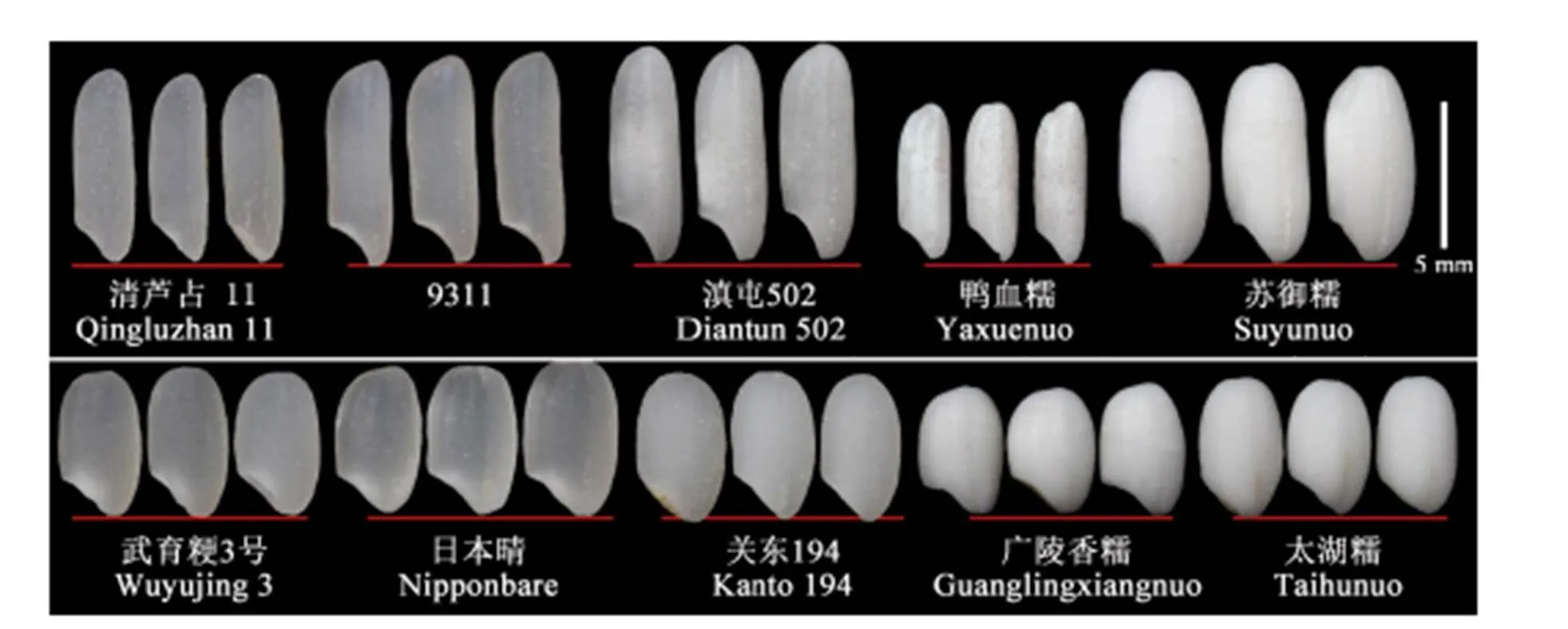
图1 不同品种稻米整体外观
Fig. 1.Rice grain appearance of different rice cultivars.
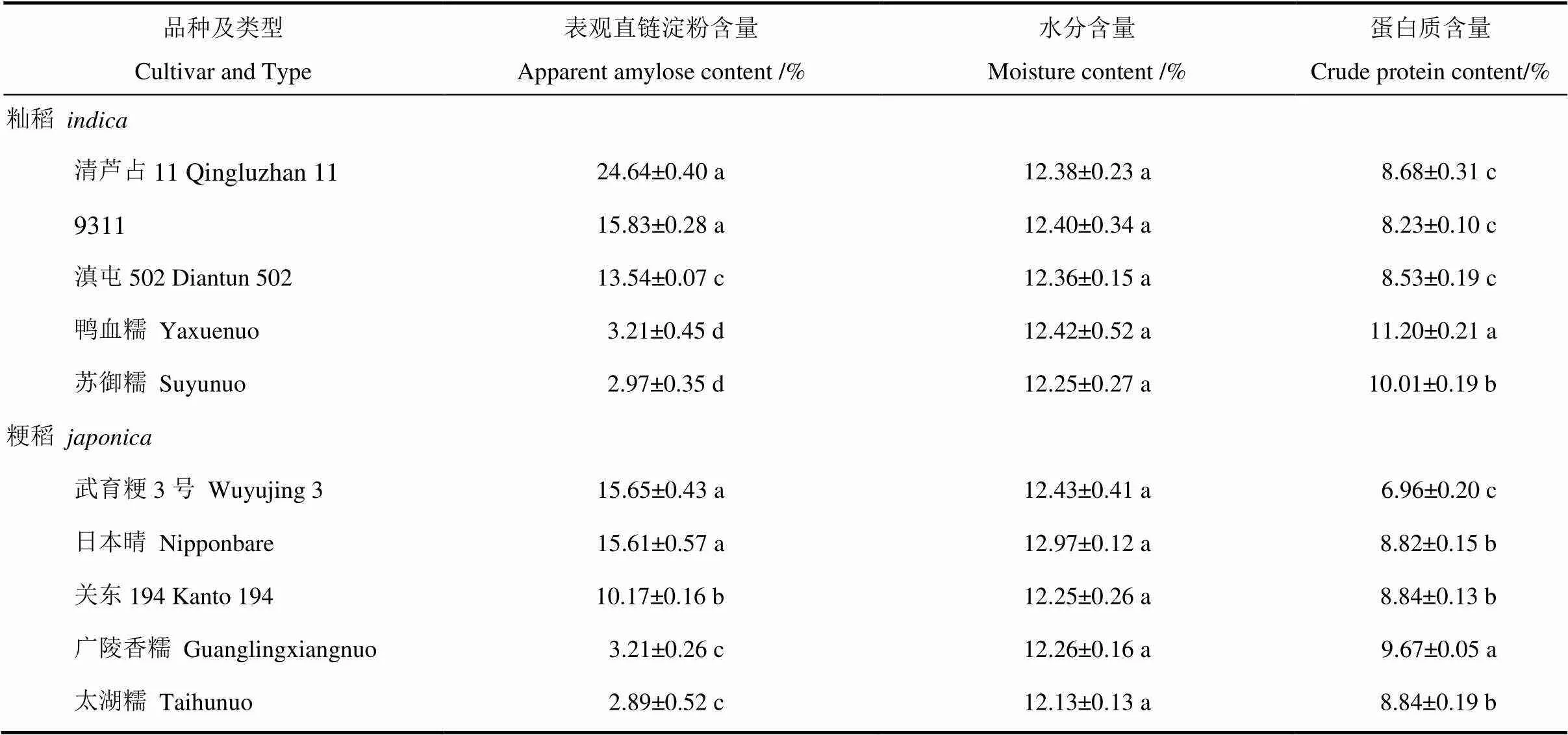
表1 不同品种稻米基本理化指标
同一列中,数据后跟相同小写字母者表示在0.05水平上差异不显著。
In the same column, the data followed by the same lowercase indicates that the difference is not significant at the 0.05 level.
稻米的外观表现受多种因素影响,如含水量、表观直链淀粉含量(AAC)和蛋白质含量等。水分含量是影响稻米外观的一个重要因素,一般水分含量高的稻米透明度好。为了分析不同透明度稻米间含水量是否存在差异,对不同精米样品的含水量进行了测定,发现所有样品的水分含量均在12%左右(表1),无显著差异。通过分析不同样品的AAC,发现籼稻品种中AAC有高、中、低和糯四类(表1)。中高AAC稻米如清芦占11和9311的外观均表现透明,而中低AAC如滇屯502为暗胚乳;在粳稻中,AAC存在中、低和糯三类,其中中间类型AAC稻米如武育粳3号和日本晴的外观正常,而低AAC的关东194也为暗胚乳表型。进一步分析了不同样品的蛋白质含量,发现籼稻中糯稻蛋白质含量明显高于非糯稻品种,而非糯稻米如清芦占11、9311和滇屯502的蛋白质含量较为接近(表1);在粳稻中,广陵香糯稻米的蛋白质含量最高,武育粳3号最低(表1)。上述结果说明不同品种稻米的透明度差异可能与AAC是直接相关的,而与水分含量和蛋白质含量没有直接关系。
2.2 不同处理对稻米断面电镜观察的影响
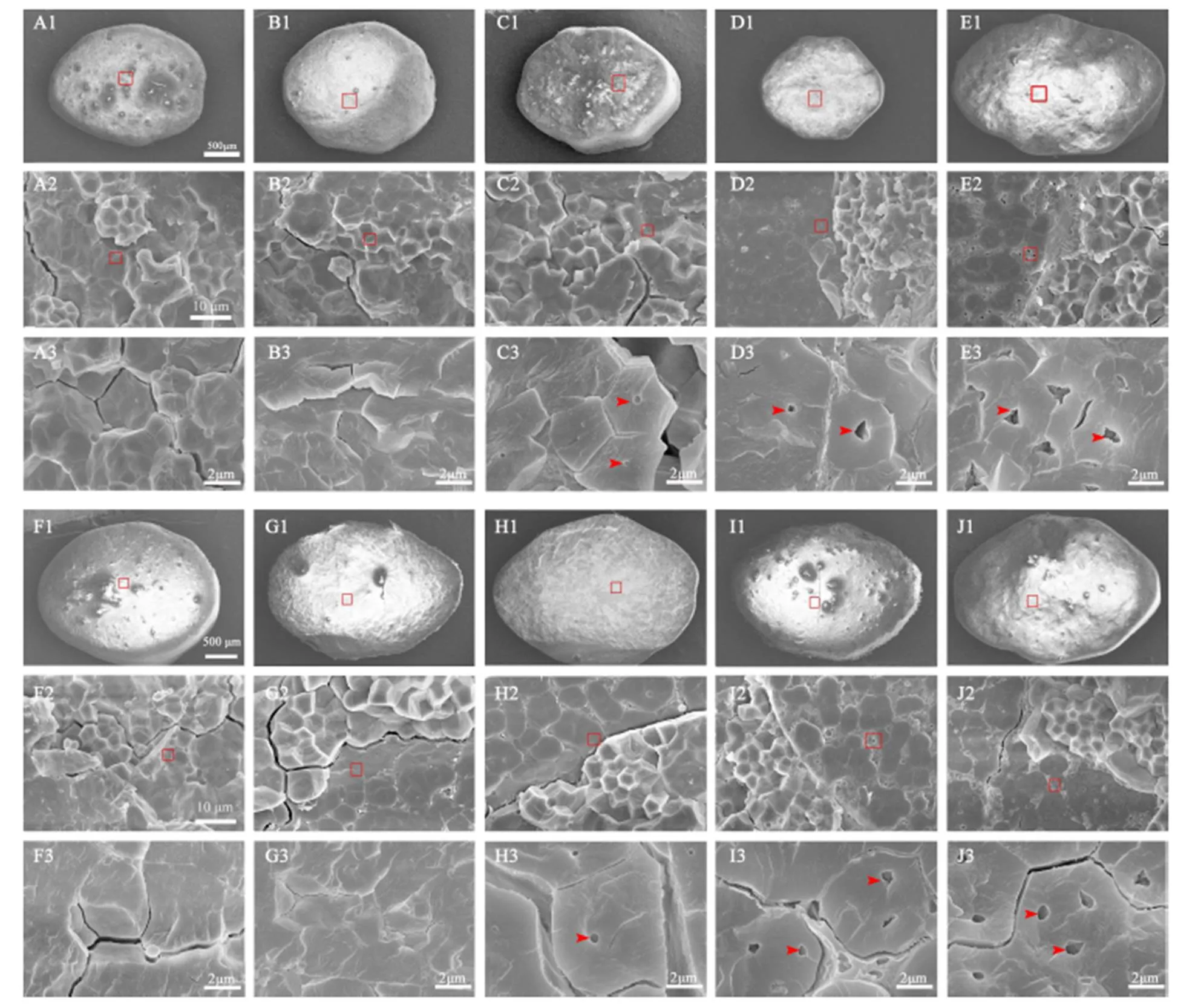
为了观察稻米淀粉粒中是否存在气腔,对稻米横断面的断裂方式进行了优化。通过比较10个水稻品种稻米三种断裂方法的扫描结果发现,采用常规自然断裂方法能观察到淀粉粒规则的外部形态,不同品种间在胚乳断面的正常区域(非垩白区)的淀粉粒排列紧密,单个淀粉粒形状规则(图2-A1~E1, 图3-A1~E1)。尽管该方法对于观察具有完整结构的淀粉粒非常方便,但是很难观察到单个断裂的淀粉粒。而利用液氮脆断法,籼稻(图2-A2~E2)和粳稻(图3-A1~E1)样品中尽管也能在少数区域观察到具有完整淀粉粒的区域,但是,在多数区域都能清楚观察到单个淀粉粒断裂的情况(图2、图3)。通过分析,发现无论是籼稻还是粳稻背景,籽粒透明的稻米中,单个淀粉粒的断面都比较光滑,几乎没有孔洞,而暗胚乳和糯稻的单个淀粉粒的断面都能观察到孔洞,且糯稻中孔洞多而大。考虑到制样的方便性,我们又比较了常规自然断裂后用刀片对断面刮蹭处理的方法,发现处理后样品断面刀痕非常明显(图2-A3~E3,2-A3~E3),尽管在糯稻中能够模糊观察到淀粉粒中的空腔(图2-D3~E3,图4-I3~J3),但图片非常粗糙,无法进行细节分析。因此,通过比较认为,液氮脆断稻米胚乳是一种有效的观察单淀粉粒断面结构的方法。
2.3 不同稻米断面空腔分析
利用液氮脆断法分别对5个籼稻和5个粳稻品种稻米横断面淀粉粒中的空腔有无、数目和大小进行了分析。在籼稻背景下,两个透明稻米的淀粉粒断面都看不到孔洞(图4-A1~A3, B1~B3),而在极少数淀粉粒表面发现有较小的孔洞,大多位于淀粉与储藏蛋白结合处。具有暗胚乳的滇屯502胚乳断面中可以观察到明显的孔洞分布,几乎所有的淀粉粒断面处都有孔洞(图4-C1~C3)。在两个糯稻中,只要有淀粉粒断裂的断面,就能观察到明显的孔洞分布,且部分淀粉粒中存在1~2个明显较大的孔洞(图4-D1~D3, E1~E3)。在粳稻背景下,具有正常透明度的武育粳3号和日本晴淀粉粒断面也未观察到孔洞(图4-F1~F3, G1~G3)。而在软米关东194的胚乳断面,可见明显散布的孔洞,单个淀粉粒一般存在1个孔洞(图4-H1~H3)。与籼糯相似,2个粳糯品种胚乳淀粉粒断面可见明显散布的孔洞,且单淀粉粒孔洞数目明显增加,一般存在2个以上的较大的孔洞(图4-I1~I3, J1~J3)。
为定量比较不同品种间孔洞数目和大小的差异,详细分析了不同稻米横断面单个断裂淀粉粒中孔洞的数目和大小。籼稻滇屯502淀粉粒中平均孔洞数目在1.1个左右,平均孔洞直径为275 nm;而两个籼糯淀粉粒的可见孔洞数目最多,接近2,平均孔洞直径也极显著大于滇屯502(表2)。此外,在粳稻关东194淀粉粒中平均孔洞数目为1,平均直径318 nm;而两个粳糯淀粉粒中的平均孔洞数目接近2,空腔的直径也极显著大于关东194(表2)。通过上述分析发现,不管是籼稻还是粳稻,单粒淀粉中孔洞个数与直径大小呈明显的正相关,表现为孔洞数量越多,孔洞直径越大。
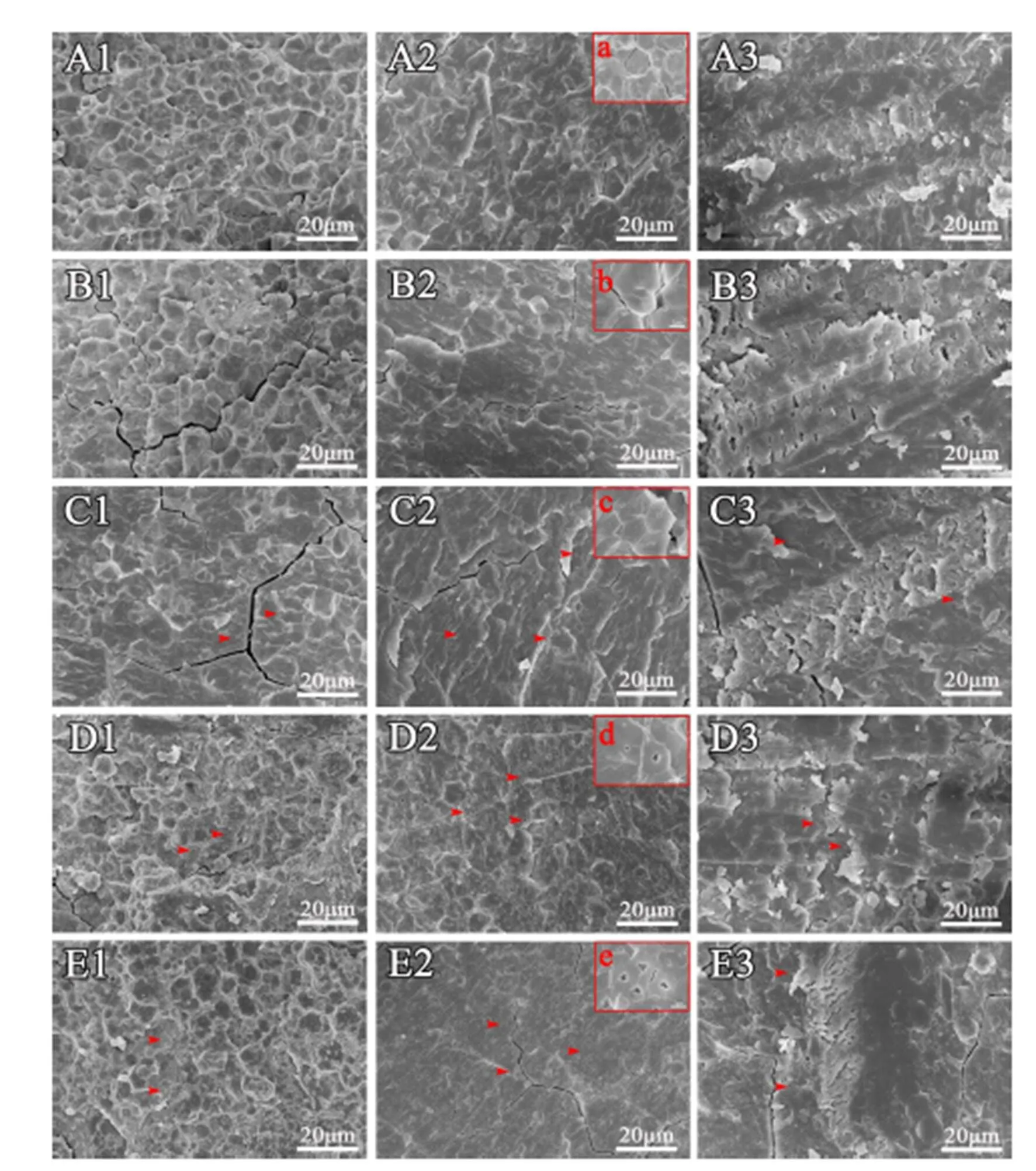
A-清芦占11; B-9311; C-滇屯502; D-鸭血糯; E-苏御糯。1-正常掰断处理; 2-液氮冷冻处理; 3-正常掰断后双面刀片处理。箭头示淀粉粒内孔洞。标尺=20 μm。
Fig. 2. Scanning electron microscopic images ofrice cross-section obtained by different methods.
2.4 水分含量对稻米外观和淀粉结构的影响
水分含量对稻米外观具有重要的影响,通常水分含量高的稻米透明度好。因此,为分析不同水分含量对上述稻米尤其是软米和糯米外观及淀粉粒内部结构的影响,选取了籼稻软米滇屯502、糯稻苏御糯和粳稻软米关东194、糯稻广陵香糯4种稻米进行梯度烘干处理分析。如图5-A所示,刚收获的稻米水分含量非常高,外观透明。随着烘干时间的增加,稻米水分含量显著下降,胚乳逐渐变暗。烘6 h后两个糯稻稻米彻底变为乳白色,软米滇屯502和关东194的胚乳均开始变暗;烘12 h后,两个软米呈现典型的暗胚乳表型。
为分析不同水分含量下稻米透明度是否与胚乳淀粉粒中的孔洞数目和大小存在相关性,针对不同处理的样品进行了籽粒横断面的SEM观察和淀粉粒孔洞数目和大小的分析(图5)。如图5-C所示,在高水分含量下的滇屯502,其部分淀粉粒中央仍可以观察到少量孔洞存在(图5-C1),分析认为,由于样品在喷金镀膜处理和扫描电镜观察时均处于真空环境中,导致样品失水,从而使淀粉内出现空洞。此外,随着水分的减少,能够观察到的孔洞概率明显增多(图5-C2,C3)。定量分析表明,高水分条件下可观察到的单淀粉粒中孔洞数基本为1个,孔洞直径较小;随着水分含量下降,单淀粉粒中可观察的孔洞数目增加,可见2个孔洞,且孔洞直径也明显增大(图5-B)。对糯稻而言,在高水分条件下可观察到部分淀粉粒内存在孔洞(图5-D1),且随着水分含量降低,单淀粉粒中的孔洞数目有明显增多的趋势,多数淀粉粒内可见2~3个孔洞(图5-B,图5-D2,D3)。
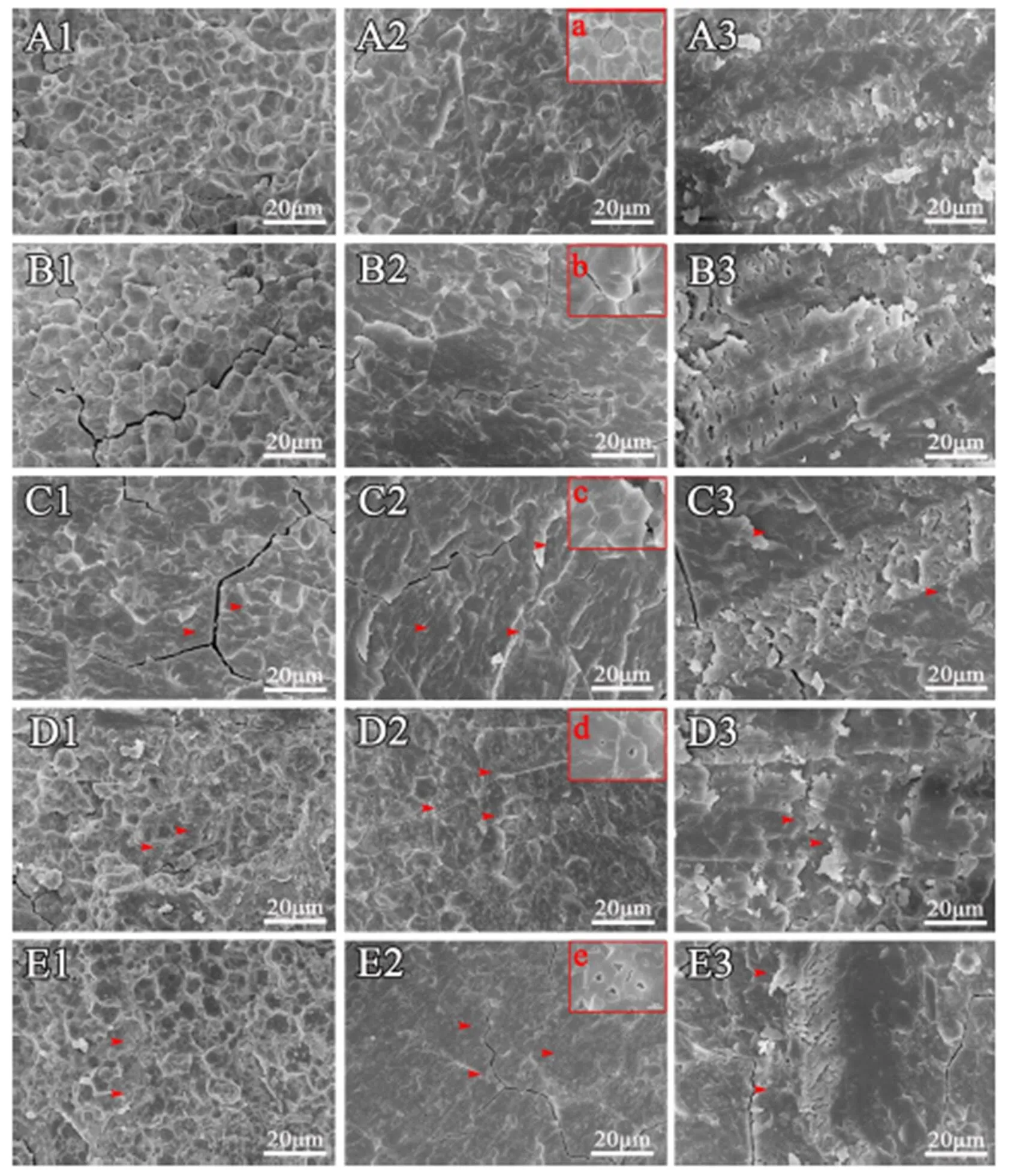
A-武育粳3号; B-日本晴; C-关东194; D-广陵香糯; E-太湖糯。1-正常掰断处理; 2-液氮冷冻处理; 3-正常掰断后双面刀片处理。箭头示淀粉粒内孔洞。标尺=20 μm。
Fig. 3. Scanning electron microscopic images ofgrain cross-section obtained by different methods.
此外,关东194和广陵香糯样品也表现出相似的趋势,即随着水分含量的降低,单淀粉粒中的孔洞数目和直径有增大的趋势(图5-B~F)。该烘干处理实验表明,对于软米和糯米而言,高水分含量条件下稻米淀粉粒中的空腔数目少且体积小,因此表现为外观透明,而随着水分的减少,淀粉粒中的空腔数目增多,体积变大,因此造成了暗胚乳或蜡质表型的出现。
2.5 不同稻米淀粉粒比较
稻米胚乳断面观察无法系统分析淀粉粒的完整形态结构和粒径分布,为明确不同稻米淀粉粒表面结构和粒径是否存在差异,进一步分离了不同品种稻米的淀粉,并对淀粉粒进行了SEM分析。通过比较发现,无论是籼稻还是粳稻,不同品种稻米的淀粉粒都非常规则,表面存在较浅的小孔(图6小箭头所示),分析认为这些孔洞是在胚乳发育后期,淀粉体中蛋白体由于受挤压而陷入淀粉粒所形成的[10]。另外,暗胚乳样品中偶尔可以观察到开裂的淀粉粒,尤其是糯稻来源的淀粉粒中间有明显的孔洞(图6-C~E, H~J大箭头所示),这与籽粒断面的扫描结果非常吻合,进一步证明了上述的观察结果。此外,部分样品中存在淀粉粒碎片,分析与研磨过度有关,该现象在品种间没有显著的差异。进一步统计了不同品种淀粉粒的大小和粒径分布,发现不同品种间淀粉粒平均直径存在一定差异,平均直径从4.5 μm到6.5 μm不等,但是这些差异与品种的外观以及AAC没有明显的相关性。从淀粉粒径分布曲线来看,籼稻背景下几个品种间的粒径分布相差较大(图6-K),而5个粳稻淀粉的粒径分布较为集中(图6-L)。总体来看,不同品种间淀粉粒大小和分布没有明显的规律性。
A-清芦占11; B-9311; C-滇屯502; D-鸭血糯; E-苏御糯; F-武育粳3号; G-日本晴; H-关东194; I-广陵香糯; J-太湖糯。箭头示淀粉粒内孔洞。2和3分别为1和2图中方框的放大图片。
Fig. 4. Scanning electron microscopic images of rice grain cross-section obtained by the liquid nitrogen freezing treatment.
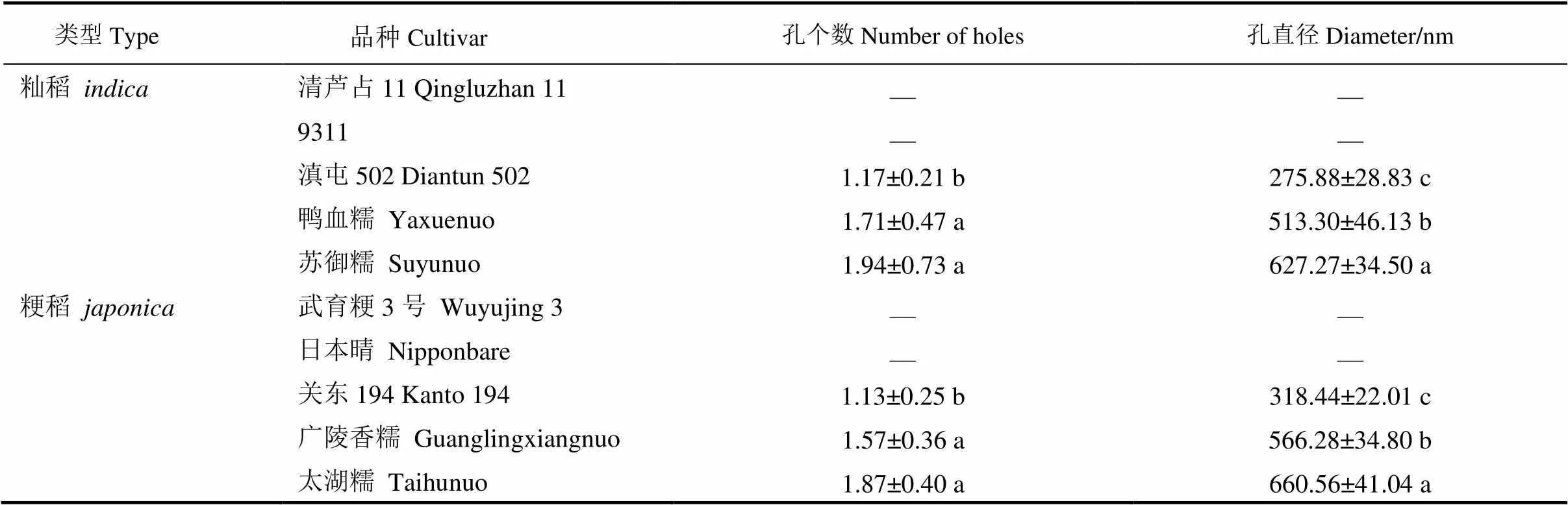
表2 不同品种水稻稻米单个淀粉粒断面孔洞比较
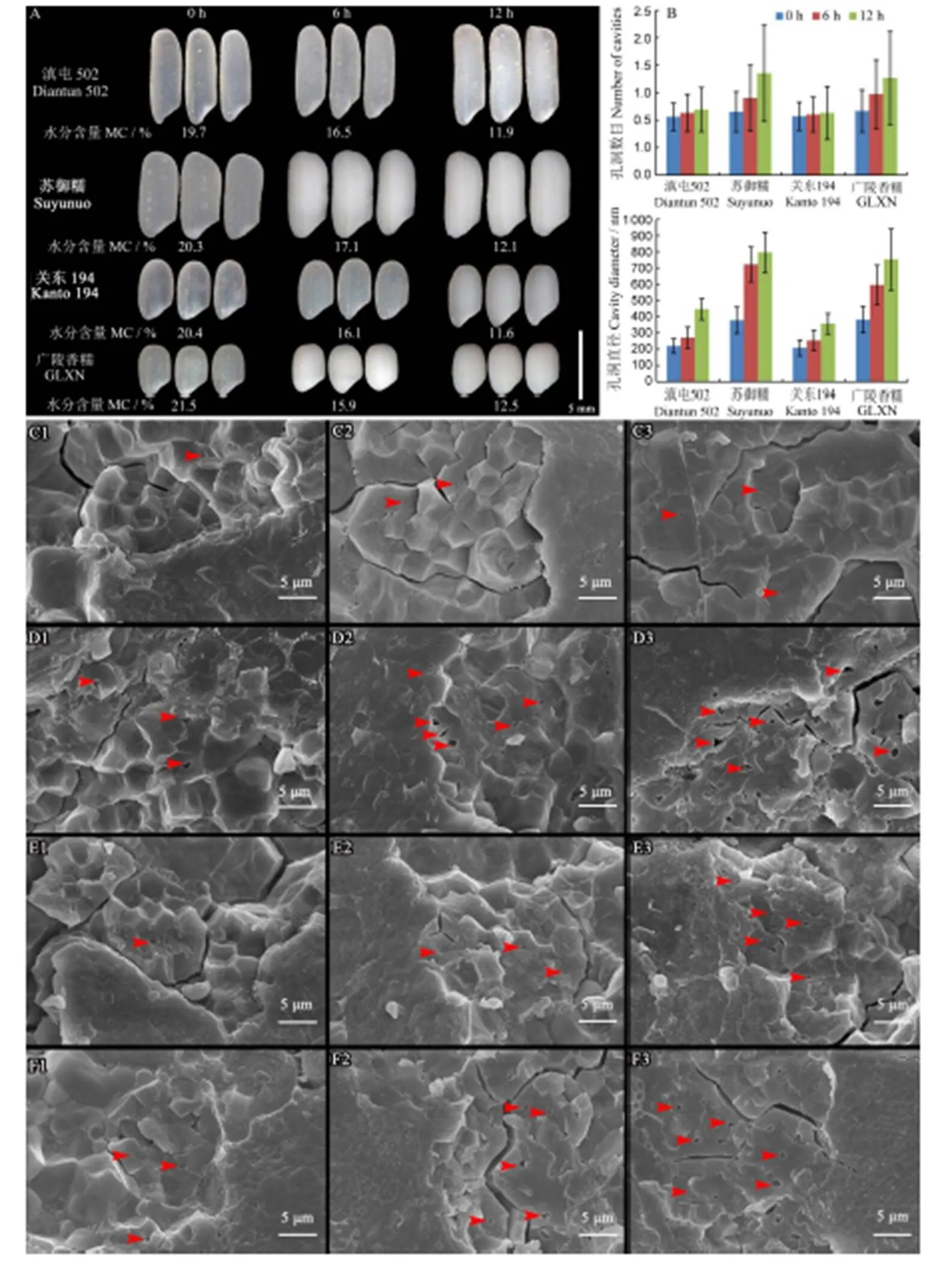
A-不同烘干处理的稻米外观。B-不同烘干处理的淀粉粒孔洞数目和孔洞大小; C-滇屯502; D-苏御糯; E-关东194; F-广陵香糯。
Fig. 5.Rice grain appearance and scanning electron microscopic images of grain cross-section.
3 讨论
稻米透明度是育种专家和广大消费者关注的性状,也是优质稻米定级的一个重要指标,该性状受多种因素影响,如含水量、粒形、垩白度、生长环境等[11-13]。一般常规的粳稻和籼稻稻米在透明度评价中都能够达到优质稻米的要求,然而对于近年来大量上市的软米而言,在水分含量较低的情况下由于稻米透明度极显著下降而往往达不到优质稻米标准。关于暗胚乳稻米乃至糯稻稻米透明度的形成存在不同的观点,本研究通过优化稻米横断面断裂方式明确了液氮脆断法是观察单个淀粉粒断面的有效方式,同时,通过在籼稻和粳稻背景下对具有不同透明度的稻米胚乳淀粉粒排列方式以及内部结构和大小等指标进行了系统的分析,明确了暗胚乳和糯稻中淀粉粒中间的空腔是暗胚乳形成的主要原因,这与垩白稻米中部分不透明区域的松散淀粉粒排列方式明显不同[14]。目前在稻米外观品质研究方面主要集中在稻米垩白粒率和垩白度方面,一般认为垩白度高的稻米透明度较差,在遗传调控方面已经克隆了系列相关基因或QTL,并且对其形成的机制有了一定的了解[15-16]。然而在稻米透明度的遗传调控和形成机制方面研究较少,且没有明确的直接调控基因被鉴定到。与之相反的是在对于一些暗胚乳突变的研究表明,GBSSⅠ的一些低酶活自然突变或者转基因修饰的低酶活蛋白,由于合成较少的直链淀粉(13%以下)而使稻米表现出半透明的表型,说明两者具有直接的相关性[17-19]。此外,通过对一些水稻粉质突变体的研究,发现胚乳中储藏蛋白的错误转运也会导致稻米不透明[20-21],当然也有部分粉质突变体(包括高垩白突变体)与胚乳的灌浆和直链淀粉的含量有关[22-24],但通过胚乳横断面的扫描电镜观察发现,上述粉质突变或高垩白突变的不透明表型主要是是由于淀粉粒的排列松散所造成的,而与本研究中所观察到的淀粉粒中的孔洞没有关系。
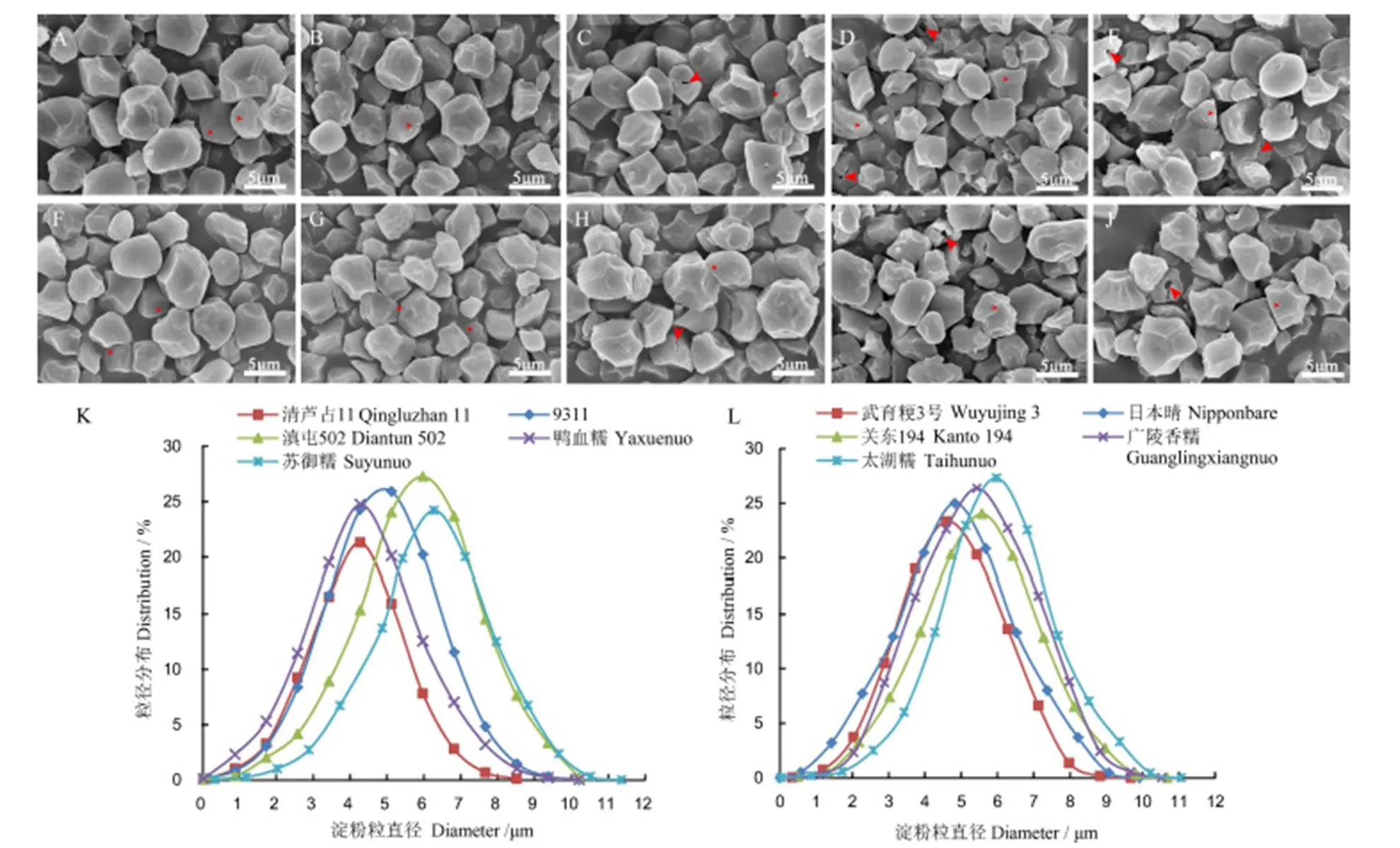
A-清芦占11; B-9311; C-滇屯502; D-鸭血糯; E-苏御糯; F-武育粳3号; G-日本晴; H-关东194; I-广陵香糯; J-太湖糯。K-籼米粒径分布; L-粳米粒径分布。小箭头示淀粉粒表面小孔,大箭头示淀粉粒内孔洞。
Fig. 6. Scanning electron microscopic images of isolated starch from different cultivars.
有关淀粉粒孔洞结构的发现实际上早在1981年就有报道,但仅在糯稻中有发现,且其是否与透明度和直链淀粉含量有关并不清楚[8]。最近,本课题组通过对不同人工修饰的GBSSⅠ转基因材料分析发现,转基因稻米的AAC越低,稻米透明度越差,稻米淀粉粒中间的空腔也越多[7],因此,认为AAC与稻米透明度以及淀粉粒中的空腔数目有着直接联系。然而在常规栽培品种中这种关联是否存在并不清楚。为明确这种相关性,本研究通过选用具有不同AAC的籼稻和粳稻品种,通过方法优化和统计分析,进一步明确了这种现象在普通栽培稻中普遍存在,并且发现稻米透明度随着淀粉粒中央孔洞的数目和尺寸的增加而下降,说明淀粉粒中的空腔是造成暗胚乳和蜡质胚乳的直接原因。空腔的存在不仅造成了较差的外观,而且直接造成了胚乳中可储藏淀粉总量的减少,从而影响稻米潜在的产量和品质。从空腔的形成来看,梯度烘干实验表明在高水分含量条件下,淀粉粒中存在极少的空腔且空腔体积小,说明单淀粉粒在高水分含量时其中的淀粉分子分布比较均匀,而在干燥失水的过程中,可能由于淀粉粒中直链和支链淀粉之间的结合不够紧密,或者结合态水在低直链淀粉含量的情况下更容易释放,而造成淀粉粒内部结构的不均一,从而拉伸出现空腔。从目前优质稻米的选育来看,培育中低直链淀粉含量水稻品种是当下培育优良食味稻米的重要途径,尤其是一些软米品种的大面积推广使得通过提高AAC来改善稻米透明度变得难以实现。当然,从目前软米的销售来看,通过对高水分含量稻米的真空包装可以保证软米的较好外观,但在开袋食用后会造成水分含量降低从而不可避免的出现暗胚乳表型。从胚乳淀粉粒孔洞出现的位置来看,淀粉粒的早期发育过程可能与孔洞的形成密切相关,目前水稻中尚未有相关基因被克隆,但在拟南芥中发现淀粉合成酶SS4与淀粉粒导向蛋白PTST(protein targeting to starch)协同起始淀粉粒的合成[25-26]。因此,在水稻中是否可以通过修饰淀粉粒起始合成相关基因来改变淀粉粒内部淀粉分子间的组织结构来提高稻米透明度可能是今后培育透明且具优良食味稻米新品种的一个努力方向。
[1] 张昌泉, 赵冬生, 李钱峰, 顾铭洪, 刘巧泉. 稻米品质性状基因的克隆与功能研究进展. 中国农业科学, 2016, 49(22): 4267-4283.
Zhang C Q, Zhao D S, Li Q F, Gu M H, Liu Q Q. Study on the cloning and function of rice quality trait genes., 2016, 49(22): 4267-4283. (in Chinese with English abstract)
[2] Fitzgerald M A, McCouch S R, Hall R D. Not just a grain of rice: The quest for Quality., 2009, 14(3): 133-139.
[3] Sreenivasulu N, Butardo V M J, Misra G, Cuevas R P, Anacleto R, Kavi Kishor P B. Designing climate-resilient rice with ideal grain quality suited for high-temperature stress., 2015, 66 (7): 1737-1748.
[4] Wang Y J, WangY L, Sun X D, Cai Z M, Yang J B, Cui D, Cao G L, Ma X D, Han B, Xue D Y, Han L Z. Influence of ethnic traditional cultures on genetic diversity of rice landraces under on-farm conservation in southwest China., 2016, 12(1): 51.
[5] Zeng D L, Tian Z X, Rao Y C, Dong G J, Yang Y L, Huang L C, Leng Y J, Xu J, Sun C, Zhang G H, Hu J, Zhu L, Gao Z Y, Hu X M, Gao L B, Xiong G S, Wang Y H, Li J Y, Qian Q. Rational design of high-yield and superior-quality rice., 2017, 3: 17031.
[6] 吴殿星, 夏英武, 李旭. 水稻胚乳外观云雾性状形成基础及其快速识别条件分析. 中国水稻科学, 2001, 15(3): 192-196.
Wu D X, Xia Y W, Li X. Formation foundation of rice endosperm appearance and its rapid identification condition., 2001, 15(3): 192-196. (in Chinese with English abstract)
[7] Zhang C Q, Chen S J, Ren X Y, Lu Y, Liu D R, Cai X L, Li Q F, Gao J P, Liu Q Q. Molecular structure and physicochemical properties of starches from rice with different amylose contents resulting from modification of OsGBSSI activity., 2017, 65(10): 2222-2232.
[8] Satoh H, Omura T. New endosperem mutations induced by chemical mutagens in riceL., 1981, 31(3): 316-326.
[9] Zhang C Q, Zhu L J, Shao K, Gu M H, Liu Q Q. Toward underlying reasons for rice starches having low viscosity and high amylose: Physiochemical and structural characteristics., 2013, 93(7): 1543-1551.
[10] 郑彦坤, 曾德二, 魏和平, 许远, 顾蕴洁, 王忠. 水稻胚乳组织的结构观察. 中国水稻科学, 2017, 31(1): 91-98.
Zheng Y K, Zeng D E, Wei H P, Xu Y, Gu Y J, Wang Z. Structure observation of rice endosperm tissue., 2017, 31(1): 91-98. (in Chinese with English abstract)
[11] Wang S K, Wu K, Yuan Q B, Liu X Y, Liu Z B. Control of grain size, shape and quality byin rice., 2012, 44(8): 950.
[12] Li Y B, Fan C C, Xing Y Z, Yun P, Luo L J, Yan B, Peng B, Xie W B, Wang G W, Li X H, Xiao J H, Xu C G, He Y Q.encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice., 2014, 46(4): 398-404.
[13] Zhang C Q, Zhou L H, Zhu Z B, Lu H W, Zhou X Z, Qian Y T, Li Q F, Lu Y, Gu M H, Liu Q Q. Characterization of grain quality and starch fine structure of tworice () cultivars with good sensory properties at different temperatures during the filling stage., 2016, 64(20): 4048-4057.
[14] Lin Z M, Zheng D Y, Zhang X C, Wang Z X, Lei J C, Liu Z H, Li G H, Wang S H, Ding Y F. Chalky part differs in chemical composition from translucent part of japonica rice grains as revealed by a notched-belly mutant with white-belly.,2016, 96(11): 3937-3943.
[15] Gao Y, Liu C L, Li Y Y, Zhang A P, Dong G J, Xie L H, Zhang B, Ruan B P, Hong K, Xue D W, Zeng D L, Guo L B, Qian Q, Gao Z Y. QTL analysis for chalkiness of rice and fine mapping of a candidate gene for., 2016, 9(1): 41.
[16] Wang X Q, Pang Y L, Wang C C, Chen K, Zhu Y J, Shen C C, Ali J, Xu J L, Li Z K. New candidate genes affecting rice grain appearance and milling quality detected by genome-wide and gene-based association analyses., 2016, 7: 1998.
[17] Mikami I, Uwatoko N, Ikeda Y, Yamaguchi J, Hirano H Y, Suzuki Y, Sano Y. Allelic diversification at thelocus in landraces of Asian rice., 2008, 116(7): 979-989.
[18] Sato H, Suzuki Y, Okuno K, Hirano H, Imbe T. Genetic analysis of low-amylose content in a rice variety, ‘Milky Queen’., 2001, 3: 13-19.
[19] Liu D, Wang W, Cai X. Modulation of amylose content by structure-based modification of OsGBSS1 activity in rice (L.)., 2014, 12(9): 1297-1307.
[20] Tian L H, Dai L L, Yin Z J, Fukuda M, Kumamaru T, Dong X B, Xu X P, Qu L Q. Small GTPase Sar1 is crucial for proglutelin and α-globulin export from the endoplasmic reticulum in rice endosperm., 2013, 64(10): 2831-2845.
[21] Wang Y H, Ren Y L, Liu X, Jiang L, Chen L M, Han X H, Jin M N, Liu S J, Liu F, Zhou K N, Su N, Bao Y Q, Wan J M. OsRab5a regulates endomembrane organization and storage protein trafficking in rice endosperm cells., 2010, 64(5): 812-824.
[22] Zhong H S, Peng F F, San F L, Gui A J, Li H X, Pei S H, Shao Q T, Xiang J W. Phenotype of rice floury endosperm mutantand fine mapping of mutated Gene., 2015, 22(4): 162-170.
[23] Zhang D P, Wu J G, Zhang Y J, Shi C H. Phenotypic and candidate gene analysis of a new floury endosperm mutant () in rice., 2012, 30(6): 1303-1312.
[24] Wu Y P, Pu C H, Lin H Y, Huang H Y, Huang Y C, Hong C Y, Chang M C, Lin Y R. Three novel alleles of() confer dull grains with low amylose content in rice., 2015, 233: 44-52.
[25] Pfister B, Zeeman S C. Formation of starch in plant cells., 2016, 73(14): 2781-2807.
[26] Seung D, Boudet J, Monroe J, Schreier T B, David L C, Abt M, Lu K J, Zanella M, Zeeman S C. Homologs of PROTEIN TARGETING TO STARCH control starch granule initiation inleaves., 2017, 29(7): 1657-1677.
Scanning Electron Microscopic Analysis of Grain Cross-section from Rice with Different Transparency
LU Yan1,2, ZHANG Xiaomin2,3, QI Yan2,3, ZHANG Changquan1, LING Yuping3, LIU Qiaoquan1,*
(Jiangsu Key Laboratory for Crop Genetics and Physiology / Key Laboratory of Plant Functional Genomics of Ministry of Education / Co-Innovation Center for Modern Production Technology of Grain Crops, Yangzhou University, Yangzhou, China;2Instrumental Analysis Center, Yangzhou University, Yangzhou 225009, China;3College of Horticulture and Plant Protection, Yangzhou University, Yangzhou 225009, China;)
【Objective】 The objective of this study was to set up an ideal method of observation of a single starch granule cross-section, to clarify the relationship between rice transparency and amylose content and to find the structure foundation that caused the poor rice transparency. 【Method】 The pre-treated rice grain cross-section and isolated starch were observed under a scanning electron microscopy.【Result】The transparency of bothandsoft rice with amylose contentbelow 15%was significantly decreased. By comparingdifferent methods of grain cross-section pre-treatments, we found that the direct mechanical fracture method can’t break a single starch granule and the mechanical fracture with glass scraping method could break down an individual starch granule, however, it will cause rough surface and thus difficult to be analyzed quantitatively. As for the liquid nitrogen fracture method, we found it’s an ideal way to break a single starch granule and the cavity in starch granule can be easily observed. We then further analyzed the starch granule arrangement and the internal structure of an individual starch granule in paddy rice from bothandrice cultivars. We found that all the starch granules arranged tightly and obvious air containing space was observed in an individual starch granule from grains of all dark endosperm rice. Also, we found that the number and area of starch granule cavity in glutinous rice was significantly higher than those in dark endosperm rice. Furthermore, we proved that the number and area of starch granule cavity increased as the grain moisture content decreased.【Conclusion】The direct liquid nitrogen fracture treatment of rice grain cross-section is an ideal method for the single starch granule cross-section observation. Besides, the rice grain transparency showed a close relationship with the moisture content and also the number and size of cavity in the middle of a starch granule but not the starch granule arrangement. Moreover, the size and number of cavities increased as the amylose content decreased.
L.; starch; scanning electron microscopy; transparency; cavity
Corresponding author, E-mail: qqliu@yzu.edu.cn
Q944.6; S511.01
A
1001-7216(2018)02-0189-11
2017-09-01;
2017-11-15。
江苏省自然科学基金资助项目(BK20160464);江苏省高校自然科学研究面上项目(16KJB210011);国家重点研发计划资助项目(2016YFD0100501);国家转基因专项(2016ZX08001006);国家自然科学基金资助项目(31561143008; 31401354)。
通讯联系人, E-mail: qqliu@yzu.edu.cn
10.16819/j.1001-7216.2018.7107
