多星韭分布格局对末次盛冰期以来气候的响应
2018-04-08谢登峰周颂东何兴金
白 娟,谢登峰,周颂东,何兴金
(四川大学 生命科学学院,生物资源与生态环境教育部重点实验室,成都 610064)
物种的分布区域受环境因素的制约,其中气候因素是影响物种分布的主要因子,因此气候因素对物种的分布有着极为关键的作用[1]。末次盛冰期产生的冰盖以及地球轴向变化引起的太阳辐射变化,造成了日照的改变和冰盖的撤退,极大地影响了中高纬度地区的气候环境,从而导致了北半球和南半球的植被调整[2]。全新世中期 (6 ka BP) 是暖期高温高湿,而末次盛冰期 (18 ka BP) 是冰期寒冷干燥,这两个时期代表最近地球历史时期全球气候的两个极端状态,所以,它们是研究气候对物种分布变化的关键时期[3]。并且,这种温暖-寒冷气候的反复交替会影响到物种的进化,导致物种的灭绝以及现存物种分布范围的反复变化。此外,纬度和地形对物种的空间分布也有明显的影响,气候的剧烈变化会导致物种大面积灭绝,然后回迁到较高的纬度,或者回迁到位于低纬度具有复杂地形的避难所并改变其生存的海拔高度[4]。因此冰期气候的反复变化对物种分布格局尤为重要。
物种分布模型(species distribution models, SDM)是结合物种出现频率的观测值与环境估计值的数值工具[5]。现在对于物种的分布区预测的模型有很多种,其中最大熵模型(maximum entropy model,MaxEnt)是一种通用的机器学习方法,具有简单而精确的数学公式,适合作为物种分布建模[6]。对于MaxEnt来说,受试者工作特征曲线(receiver operating characteristic curve,ROC)下面积(area under curve,AUC)值总是较高,表明该物种适合与不适合的区域有着较好的区别。最大熵模型是比较常用的物种分布模型,主要应用于保护生物学、生态学、进化生物学以及谱系地理学等领域来观测物种的分布[7-8]。同时MaxEnt模型还可投射于以前的地质时期,如末次间冰期、末次盛冰期和全新世中期等[9]。地理信息系统(GIS)可以与MaxEnt结合分析,进行可视化表达,计算气候变化中分布稳定的区域来进行避难所推测[10]。
多星韭(AlliumwallichiiKunth.),又名石韭菜、山韭菜等,隶属于石蒜科葱亚科葱属粗跟组(Sect.BromatorrhizaEkberg)。其味辛、甘,性平,具有散瘀止痛、祛风止痒的功效,在云南省部分少数民族中常作为药物使用,是彝药的来源之一,全草均可入药。多星韭的叶片可以作为蔬菜食用,是一种药食同源的植物[11]。同时,多星韭花大而艳丽,花色多样,呈现为白色、粉色、紫色、紫红色以及紫黑色等多种花色,四倍体植株的花葶更是高大坚实又粗壮,具有极高的观赏价值和园艺开发利用前景。以多星韭为主要优势种群的贵州省赫章县韭菜坪更是壮丽的自然植被景观,极具旅游观赏价值[12]。此外,作为多年生草本植物,多星韭主要分布于青藏高原东南部的横断山脉地区,云贵高原、南岭山脉、喜马拉雅山脉南麓以及邻近国家(如印度北部、尼泊尔、不丹等地)也有分布。该种占据了灌丛、石壁、草坡、沟边、林下及林缘等多样的生境,海拔一般为(1 700~)2 300~4 800 m,是以二倍体和四倍体为主的多倍体复合体,具有较为丰富的倍性水平[13-14]。多星韭的物种分化时间大约在上新世中晚期,而物种内的主要分支分开于中新世早期[15]。多星韭具有食用、药用以及观赏价值,同时由于种内分化剧烈,具有二倍体和四倍体多种核型,是研究植物多倍化、进化与生态适应等的良好材料,因此该物种具有较高的经济价值和科研价值,本研究对它的当前分布区进行模拟,以便于更好地保护和利用这一种质资源。
由于草本植物的进化速率快于木本植物,对气候的变化更加敏锐,更适于观测植物的分布区与气候变化之间的关系。同一物种,多倍体个体大多拥有比二倍体个体更高的杂合性[16],在短时间内可以产生广泛的遗传多样性[17],这些特性都在气候动荡时期缓冲了近亲交配和遗传漂变[18]。因而多倍体个体可以容忍更广的生态位,适应新的生境。多星韭作为拥有多倍体的高海拔草本植物,可以抵御较为恶劣的气候环境。本次研究主要是通过对多星韭的分布格局进行模拟,推测多星韭对冰期以来气候变化的响应。
1 材料和方法
1.1 数据来源
物种分布数据主要来源于野外实地调查和中国数字植物标本馆(CVH)。部分标本有比较详细的地理位置信息但缺乏经纬度信息,通过Google Earth定位获取经纬度,剔除重复数据,共获取到89个多星韭在中国的详细点位分布信息(图1)。
气候变量数据从WorldClim数据库(www.worldclim.org)中下载,获取末次盛冰期(LGM)、全新世中期(Mid-Holocene)、当前(Current,2000年)以及未来(Future,2080年)4个时期的气候变量数据。气候变量数据包括19个全球环境因子变量,空间分辨率为2.5 arcmin,每个栅格面积约为22 km2。将多星韭分布点位数据和气候变量数据输入MaxEnt模型中进行分布模拟及检验。

其中黑色为标本数据,粉色为野外采集数据图1 多星韭当前分布范围及详细分布点位The black point stood for the berbarium distribution data; the pink point stood for the distribution data of vegeation surveyFig.1 The distribution range of Allium wallichii in the current and distribution points
1.2 模型检验
将多星韭当前分布数据和气候环境数据导入MaxEnt,参照Roberto等[19]研究的参数选择,随机选取75%的多星韭分布点位数据用于建立模型的训练集(Training Data),剩余25%的分布点用于模型验证(Test Data),选择刀切(Jackknife)法,设定模型运算次数1 000次,进行10次重复运算,其他参数为模型默认值,同时采用MaxEnt模型的受试者工作特征曲线(Receiver Operating Characteristic Curve,ROC)下面积AUC(Area Under Curve)值评估分析模型模拟的准确性。AUC的数值范围在0.5~1之间,AUC大小表示预测精确度,AUC>0.9表示模拟结果非常精确。
1.3 分布面积计算
利用ArcGIS10.2计算多星韭不同时期的分布面积变化。本研究的面积计算均在ArcGIS10.2中的“SDM toolbox”[20]完成。首先在ArcGIS10.2中将多星韭4个时期的分布数据ASCII文件转换为二进制文件。然后依次计算各个时期之间(LGM-Mid-Holocene、Mid-Holocene-Present、Present-Future)的分布面积变化,得到扩张的区域、稳定的区域以及收缩的区域。本研究使用的是ArcGIS10.2“SDM Tools”中的“Universal Tools”子目录中的“Distribution Changes Between Binary SDMs”工具。然后选取1个合适的等面积投影,输出Tiff(.tif)文件格式。
2 结果与分析
2.1 模型适合度
在MaxEnt模型中对当前(Current,1960~1990年)的环境因子和多星韭现在的地理分布信息进行模拟,测试数据和训练数据的AUC值高达0.98以上(图2),这说明预测结果极为准确。而对过去(Past)时期的末次盛冰期(LGM)、全新世中期(Mid-Holocene)以及未来(Future,2080年)的模拟中,AUC值至少高达0.979以上,这说明这一预测结果可信度极高,可以用于讨论多星韭分布区变化与迁移路线对第四纪冰期巨大气候变化的响应。
2.2 生态因子的讨论
贡献率较高的生态因子是最暖季度降水量、海拔、年均温变化范围和温度季节性变化标准差,贡献率依次为28.7%、22.7%、14.1%和10.3%,总计超过75%,其他因子的贡献率较低(表1)。这一结果显示在气候因子中温度对多星韭的分布有很大的影响。
2.3 多星韭不同时期的分布情况
模拟结果显示(图2),比起当前分布区,末次盛冰期(LGM)的分布区向南扩张,在中南半岛、喜马拉雅山脉南麓以及印度半岛一带均有分布,也向东部略微扩张,中国南岭地区甚至日本岛附近也有分布,但主要分布区仍是横断山脉,以中国云南省为主。
全新世中期(Mid-Holocene)气候温暖湿润,与当前的气候差异不大,多星韭分布区整体位于当前分布区的南部,主要分布于中国云南省、四川省南部、西藏林芝、昌都地区以及喜马拉雅山脉南麓。
当前分布区(Current)在云南省南部不再有分布,仍然集中分布在横断山脉地区,呈现以横断山脉为中心向两边沿山脉扩散分布的状态。通过最大熵模型预测,在未来(Future)的80年里,多星韭的分布区整体向西移动,横断山脉东部分布区也略微收缩,南岭一带分布的可能性明显降低,而在青藏高原地区,多星韭分布区略微扩张。这一现象的出现可能是由于全球温度升高所导致。
3 讨 论
3.1 重要生态因子的讨论
研究结果表明在19个气候因子中,贡献率最高的生态因子是最暖季度降水量、年均温变化范围和温度季节性变化标准差,由此可以看出温度比水分对多星韭的影响更为显著。除了气候因素,也存在其他很多因素的影响,例如地形、土壤、其他群落之间的相互作用等。在多星韭的分布区预测中,海拔拥有相当高的贡献率,高达22.7%,这是因为多星韭生长受到海拔的影响较为明显,一般生长在海拔(1 700~)2 300~4 800 m的地区。纬度相同的地区海拔越高温度越低,多星韭显然更适宜生长在相对寒冷的条件下。
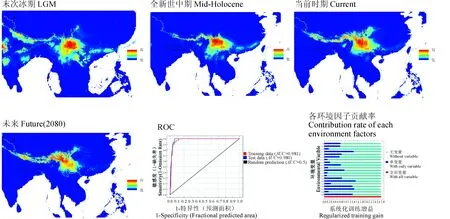
图2 基于MaxEnt及GIS的末次冰期(LGM)、全新世中期(Mid-Holocene)、当前时期(Current)及2080年(Future)多星韭的潜在分布图和当前时期的ROC值和各环境因子贡献率Fig.2 A. wallichii potential distribution map in LGM, mid-Holocene, current period and 2080 (Future), ROC value and contribution rate of each environment factors of current period based on MaxEnt model and GIS
3.2 迁移路线推测
横断山脉地区(HDM)地形地貌复杂,山脉河谷众多,形成了多种生境[21]。地理屏障丰富,导致小气候变化不大,这一地区在第四纪冰期时(< 2 Ma)还充当了很多温带物种的冰期避难所[22-23]。多星韭的主要分布区是横断山脉地区,由于山脉的阻挡,小气候稳定,冰期对多星韭的影响可能不大。黄德清等也认为多星韭的种内分化与冰期关系不大[15]。在本次研究中,多星韭在各个时期下,横断山脉的分布区均十分稳定(图3),这与推断的结果吻合。
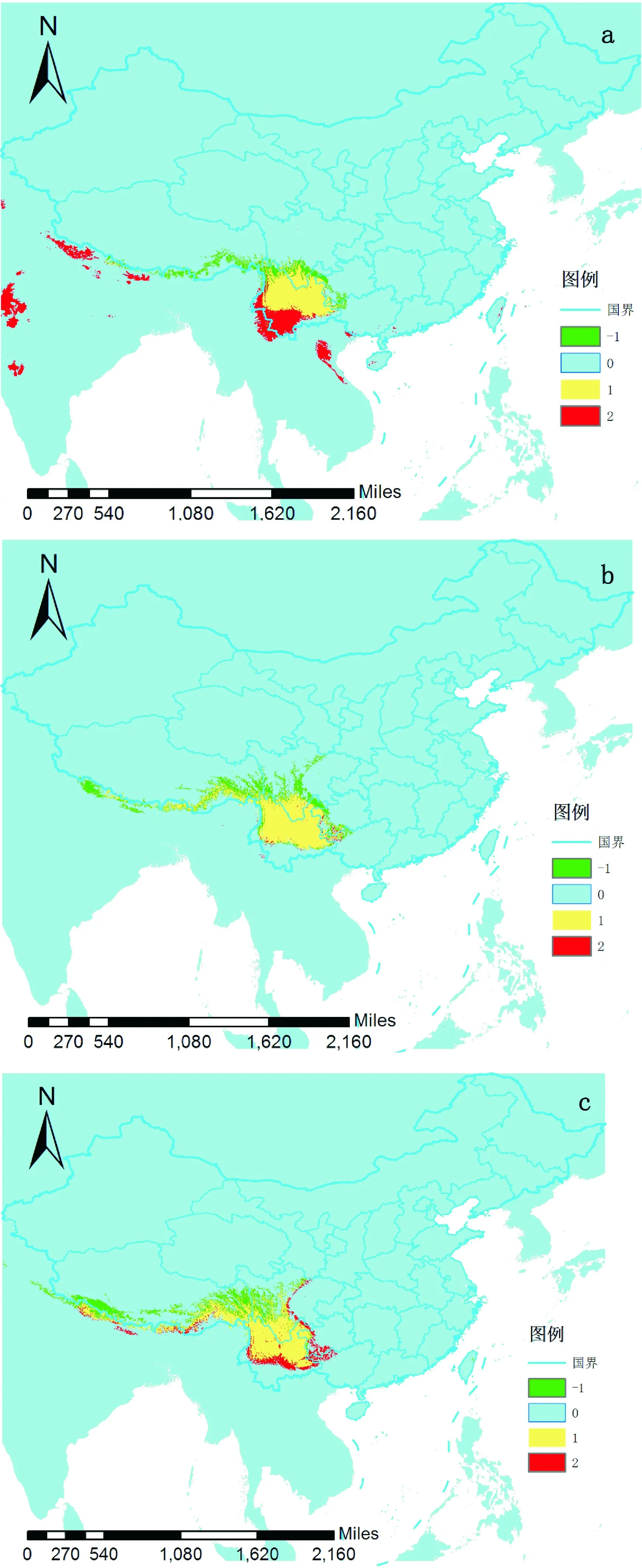
a是末次盛冰期到全新世中期(LGM-Mid-Holocene)分布变化,b是全新世中期到当前(Mid-Holocene-Current)的分布变化,c是现在到2080年(Current-Future)的分布变化。“-1”代表面积扩张的区域,“0”代表不存在物种分布的区域,“1”代表分布没有发生变化的区域,“2”代表面积收缩的区域图3 多星韭的不同时期分布变化比较The distribution was changed from the last glacial maximum to the middle Holocene in picture a; The distribution was changed from the middle Holocene to the current in picture b; The distribution was changed from the current to 2080 in picture c; “-1” stood for area expansion region; “0”stood for “no species” region; “1” stood for region distribution was not changed; “2” stood for contraction regionFig.3 Comparison of the distribution changes of different period based on MaxEnt model and ArcGIS
多星韭分布区在末次盛冰期呈现扩张趋势,分布区扩张到低纬度。在之后的时期,多星韭的分布区又开始缓慢地向北回迁。因此,横断山脉地区便存在冰期前的居群以及回迁的居群,它们会发生二次接触[24],极易出现多倍体,同时恶劣气候环境会促进未减数分裂的配子生成[25],从而加剧多倍体的出现,这一地区会形成四倍体二倍体混生居群。前人的研究[14]也证实了在横断山脉地区四倍体二倍体混生居群较多。同时由于多倍体可以适应较为恶劣的生存环境,加上葱属植物具有较强的无融合繁殖方式,其变异居群或个体尤其是三倍体和五倍体等在横断山区依靠无融合(珠芽)或营养繁殖(鳞茎或根状茎)得以保存其变异居群或个体,当其度过了冰期前或冰期恶劣气候时期后,在冰后期多星韭得以向南扩张[15]。
3.3 分布区变化
分布在青藏高原的物种在末次盛冰期来临时,一部分会退缩到青藏高原边缘地区(尤其是横断山区)度过冰期后再回迁,还有一部分物种会停留在原地的微型避难所度过冰期[26]。由于青藏高原并没有像北半球其他的一些地区形成统一的大冰盖,加之横断山脉的特殊走向,在横断山脉地区形成了许多大小不同的温和小环境,为多星韭等物种的较近祖先提供了良好的冰期避难所,从而冰期气候并未对青藏高原地区的所有植物造成不利影响或者限制作用[27]。停留在原地的物种大多是高山植物[26],适宜于低温干燥的环境,拥有比较广泛的生态位,如林下、灌丛和草坡等,且存在除种子繁殖以外的多种繁殖方式,在冰期来临时这些植物拥有多样的避难所[27]。因而冰期气候的动荡对这些植物的影响较小,甚至一些耐寒耐旱的植物比如金露梅(PotentillafruticosaL.)、红砂(Reaumuriasongarica(Pall.) Maxim.)等,通常在冰期发生扩张[28],与大部分物种对冰期的应答截然不同。通过ArcGIS计算结果显示(图3),根据MaxEnt模拟结果多星韭在末次盛冰期发生扩张,这与高山植物垫紫草(Chionocharishookeri(Clarke) Johnst.)类似[27]。冰期过后的全新世中期是一个暖期,这使得更倾向生长于寒冷环境的多星韭的分布区向北或者向高海拔迁移,以便更适宜于在低温和相对干旱的环境条件下生长。多星韭在横断山脉地区的分子证据显示在同一居群被不同支系共享,这可能是由于第四纪冰期的气候导致再次接触[15]。而在本研究结果中显示,多星韭位于横断山脉的分布区在冰期前后的确存在重叠,在一定程度上支持前人的分子结果[15]。在未来的模拟预测中,多星韭的分布区向西北移动,但主要适生区仍位于横断山脉一带。由于温室效应导致全球气温升高,多星韭则会向高纬度和高海拔的低温干旱区域迁移。
参考文献:
[1]MANTHEY M, BOX E O. Realized climatic niches of deciduous trees: comparing western Eurasia and eastern North America [J].JournalofBiogeography, 2007,34(6): 1 028-1 040.
[2]COHMAP MEMBERS.Climatic changes of the last 18 000 years: observations and model simulations [J].Science, 1988, 241: 1 043-1 052.
[3]中国第四纪孢粉数据库小组.中国中全新世 (6 ka BP)和末次盛冰期(18 ka BP)生物群区的重建[J].植物学报,2000,42(11):1 201-1 209.
Members of China Quaternary Pollen Data Base. Pollen-based biome reconstruction at Middle Holocene (6 ka BP) and Last Glacial Maximum (18 ka BP) in China [J].ActaBotanicaSinica,2000,42(11):1 201-1 209.
[4]HEWITT G M. Genetic consequences of climatic oscillations in the Quaternary[J].PhilosophicalTransactionsoftheRoyalSocietyB:BiologicalSciences, 2004, 359: 183-195.
[5]ELITH J, LEATHWICK J R. Species distribution models; Ecological explanation and prediction across space and time [J].AnnualReviewofEcologyEvolutionandSystematics, 2009, 40: 677-697.
[6]PHILLIPS S J, ANDERSON R P, SCHAPIRE R E. Maximum entropy modeling of species geographic distribution [J].EcologicalModelling, 2006, 190: 231-259.
[7]徐军,曹博,白成科.基于MaxEnt濒危植物独叶草的中国潜在适生分布区预测[J].生态学杂志,2015,34(12):3 354-3 359.
XU J, CAO B, BAI C K. Prediction of potential suitable distribution of endangered plantKingdoniauniflorain China with MaxEnt [J].ChineseJournalofEcology,2015,34(12):3 354-3 359.
[8]柳鑫,杨艳芳,宋红萍,等.基于MaxEnt和ArcGIS的黄连生长适宜性区划研究[J].中国中药杂志,2016,41(17):3 186-3 193.
LIU X, YANG Y F, SONG H P,etal. Cultural regionalization forCoptischinensisbased on 3S technology platformⅠ. Study on growth suitability forCoptischinensisbased on ecological factors analysis by Maxent and ArcGIS model [J].ChinaJournalofChineseMateriaMedaica,2016,41(17):3 186-3 193.
[9]胡忠俊,张镱锂,于海彬.基于MaxEnt模型和GIS的青藏高原紫花针茅物种分布格局模拟[J].应用生态学报,2015,26(2):505-511.
HU Z J, ZHANG Y L, YU H B. Simulation ofStipapurpureadistribution pattern on Tibetan Plateau based on MaxEnt model and GIS [J].ChineseJournalofAppliedEcology,2015,26(2):505-511.
[10]CHAN L M, BROWN J L, YODER A D. Integrating statistical genetic and geospatial methods brings new power to phylogeography [J].MolecularPhylogeneticsandEvolution,2011, 59: 523-537.
[11]刘盼盼,许云章,王静霞,等.UPLC法同时测定彝药多星韭中四种核苷类成分的含量[J].中药材,2015,8(38):1 618-1 621.
LIU P P, XU Y Z, WANG J X,et al. Determining the content of four nucleoside components inAlliumwallichiiKunth which is a medicine in the Yi nationality [J].JournalofChineseMedicinalMaterials,2015,8(38):1 618-1 621.
[12]姜克,葛敦.赫章县多星韭植物资源及其群落生态特征初报[J].宁夏农林科技,2014,55(10):38-39,51.
JIANG K, GE D. Preliminary report on ecological characteristics ofAlliumwallichiiKunth plant resources and its community in Hezhang county[J].NingxiaJournalofAgricultureandForestryScienceandTechnology,2014,55(10):38-39,51.
[13]晏一祥,黄瑞复,魏蓉城,等.葱属粗根组5种材料的核型研究[J].植物分类学报,1990,28(3):177-184.
YAN Y X, HUANG R F,WEI R C,etal. Studies on the karyotype of 5 sample ofAlliumsect. Bromatorrhiza Ekberg [J].ActaPhytotaxonomicaSinica, 1990,28(3),177-184.
[14]翟艳红,杨莹,星耀武,等.云贵高原多星韭二倍体—四倍体分布格局研究[J].植物科学学报,2011,29(1):50-57.
ZHAI Y H, YANG Y, XING Y W,etal. Diploid and tetraploid distribution ofAlliumwallichiiKunth (Ailiaceae) in the Yunnan-Guizhou Plateau [J].PlantScienceJournal,2011,29(1):50-57.
[15]HUANG D Q, LI Q Q, ZHOU C J,etal. Intraspecific differentiation ofAlliumwallichii(Amaryllidaceae) inferred from chloroplast DNA and internal transcribed spacer fragments [J].JournalofSystematicsandEvolution, 2014,52(3): 341-354.
[16]SOLTIS P S, SOLTIS D E. The role of genetic and genomic attributes in the success of polyploids [J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 2000,97(13): 7 051-7 057.
[17]SONG K, LU P, TANG K,etal. Rapid genome change in synthetic polyploids ofBrassicaand its implications for polyploid evolution [J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 1995,92(17): 7 719-7 723.
[18]BROCHMANN P C, BRYSTING A K, ALSOS I G,etal. Polyploidy in arctic plants [J].BiologicalJournaloftheLinneanSociety, 2004,82(4): 521-536.
[19]ROBERTO M, ZZMORA R, MOLINA J R,etal. Predictive modeling of microhabitats for endemic birds in South Chilean temperate forests using Maximum entropy (Maxent) [J].EcologicalInformatics, 2011,6(6): 364-370.
[20]BROWN J L. SDMtoolbox: a python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses [J].MethodsinEcologyandEvolution, 2014,5(7): 694-700.
[21]WU Y Q, CUI Z J, LIU G N,etal. Quaternary geomorphological evolution of the Kunlun Pass area and uplift of the Qinghai-Xizang (Tibet) Plateau [J].Geomorphology, 2001, 36: 203-216.
[22]GAO L M, MOLLER M, ZHANG X M,etal. High variation and strong phylogeographic pattern among cpDNA haplotypes inTaxuswallichiana(Taxaceae) in China and North Vietnam [J].MolecularEcology, 2007, 16: 4 684-4 698.
[23]CHEN S Y, WU G L, ZHANG D J,etal. Molecular phylogeography of alpine plantMetagentianastriata(Gentianaceae) [J].JournalofSystematicsandEvolution, 2008, 46: 573-585.
[24]STEBBINS G L. Polyploidy and the distribution of the arctic-alpine flora: new evidence and a new approach [J].BotanicaHelvetica, 1984, 94: 1-13.
[25]RAMSEY J, SCHEMSKE D W. Pathways, mechanisms and rates of polyploid formation in flowering plants [J].AnnualReviewofEcology, 1998, 29: 467-501.
[26]QIU Y X, FU C X, COMES H P. Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora [J].MolecularPhylogeneticsandEvolution, 2011, 59: 225-244.
[27]LUO D, YUE J P, SUN W G,etal. Evolutionary history of the subnival flora of the Himalaya-Hengduan Mountains: first insights from comparative phylogeography of four perennial herbs [J].JournalofBiogeography, 2016, 43: 31-43.
[28]于海彬,张镱锂.青藏高原及其周边地区高山植物谱系地理学研究进展[J].西北植物学报,2013,33(6):1 268-1 278.
YU H B, ZHANG Y L. Advances in phylogeography of alpine plants in the Tibetan Plateau and adjacent regions [J].ActaBotanicaBoreali-OccidentallaSinica, 2013,33(6):1 268-1 278.
