中国李属樱亚属Prunus L.subgenus Cerasus (Mill.) A.Gray的数量分类*
2018-04-02吴保欢黄文鑫石文婷羊海军崔大方
吴保欢,黄文鑫,石文婷,羊海军,崔大方
(1.华南农业大学林学与风景园林学院,广东 广州 510642;2.华南农业大学公共基础课实验教学中心,广东 广州 510642)
李属PrunusL.樱亚属subgenusCerasus(Mill.) A.Gray植物,广泛分布于北半球温带地区,其中尤以中国种类最为丰富[1-2]。
对于樱亚属的分类问题,可以分为3个方面。
首先是它的分类等级的问题。一般存在两种观点,一是将樱类植物处理为桃亚科Amgdaloideae T.& G.下面的一个独立的属CerasusMiller.[1-5];二是把樱类植物作为樱亚属subgen.Cerasus归入李属PrunusL.中[6-8]。《中国植物志》和《Flora of China》都采用了第一种处理,目前在我国仍然有很多学者[2, 9-13]使用。
其次是樱亚属的范围问题。一直以来,多数学者[1-4, 6, 7]都承认樱亚属(或樱属)包含两个类群,典型樱类(Cerasus或Typocerasus)和矮樱类(Microcerasus或Lithocerasus),但是也有少数学者认为矮樱类是一个独立的类群。
第三方面是樱亚属内部的分类问题。不同的学者对樱亚属内部类群的划分存在较大的分歧(详见表1)。Koehne[6]依照萼片反折与否,设置了反萼组sect.Cremastosepalum、 直萼组sect.Pseudocerasoides2个组,组下共设13个亚组(subsection),11个系(series)。俞德浚和李朝銮[1]在《中国植物志》没有按照反萼组和直萼组的划分,而是对Koehne系统的亚组和系进行调整合并后,设为组的等级,共设9个组。王贤荣[2]在《中国樱花品种图志》中,和Koehne一样依据萼片反折与否,将国产的樱亚属植物分为了反萼组(sect.Cerasus)和直萼组(sect.Pseudocerasus),组下不设亚组,只设12个系。

表1 Koehne、俞德浚和李朝銮、王贤荣的樱亚属分类系统Table 1 3 taxonomic systems of subgen.Cerasus by Koehne, Yü & Li and Wang Xianrong
对一些种的界定也存在不同的处理。王贤荣和向其柏[10]将多变樱PrunusvariabilisKoehne、毛筒樱PrunusrehderianaKoehne处理为独立的种,将雕核樱桃PrunuspleiocerasusKoehne并入锥腺樱桃PrunusconadeniaKoehne中,又将细花樱桃PrunuspusillifloraCardot与蒙自樱桃Prunushenryi(C.K.Schneid.) Koehne进行了合并。而作为《中国植物志》的修订版,《Flora of China》[3]对《中国植物志》的种进行了一些调整,将岩樱PrunusscopulorumKoehne并入了樱桃PrunuspseudocerasusLindl.,将西南樱桃PrunusduclouxiiKoehne并入云南樱桃PrunusyunnanensisFranch.,将光叶樱桃Prunusglabra(Pamp.) Koehne并入华中樱桃PrunusconradinaeKoehne,将多变樱与毛筒樱并入微毛樱桃PrunusclarofoliaC.K.Schneid。
随着分子系统学的发展,前两个问题基本得到解决,近20年来涉及李属的分子系统学研究[3]结果都表明广义的李属是一个单系类群,典型樱类与矮樱类都包含在这个单系群中,但是两个类群是互相独立的,矮樱类与桃李杏的关系更近。Shuo Shi 等[19]发表的李属分类系统中,将李属分为了subgen.Padus、subgen.Cerasus和subgen.Prunus3个亚属,其中subgen.Cerasus包含了典型樱类和斑叶樱PrunusmaackiiRupr.和圆叶樱桃PrunusmahalebL.。矮樱类作为一个组sect.Microcerasus被置于subgen.Prunus中。本文所使用的樱亚属定义遵从Shuo Shi等[19]的处理,包含典型樱类、斑叶樱和圆叶樱桃。至于第3个问题,目前仍然还没有得到解决。
数量分类学在解决植物分类问题的研究中有较为广泛的应用[20-24]。在樱类植物种类的界定上,C.Chang等[20-24]利用主成分析对Prunusserrulata复合体的分类进行了探讨。本文意在利用数量分类学的方法对中国樱亚属植物进行宏观形态的分析,以期对樱亚属内的分类和种的界定进行探讨。
1 材料与方法
研究材料包括41种典型樱植物,其中原产自我国的有36种,产自日本、韩国等地的有5种。以30个形态性状进行分析,每个种测量份数大于30份,包含花部性状的标本以及包含叶部性状的标本各15份(部分分类单位因馆藏量太少,则以实测为准)。数量性状的测量以每份标本每个性状测定3~5个数据,最后取各数据的均值作为原始数据。定性性状则以性状表现在观测统计中超过60%以上为准。定性性状分为二态和多态,采用二元编码和多元编码将性状编码(表2)和数量性状一起构成分析原始数据。用NTSYSpc versiong 2.1e软件进行聚类和主成分分析。

表2 41种樱亚属植物的形态性状与编码Table 2 Morphological characters and code of 41 species in subgen.Cerasus for numerical classification
2 结果与分析
利用UPGMA (unweighted pair-group method with arithmetic means)方法进行聚类,结果如图1如示。在欧氏距离6.76处,41个种被划分为11个类群,包括6个由单独种组成的分支和A、B、C、D、E 5个较大的分支。
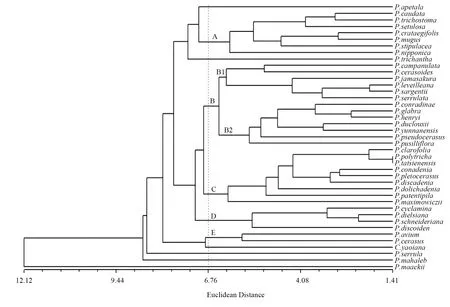
图1 基于30个形态性状的41种樱亚属植物UPGMA聚类分析结果Fig.1 Result of UPGMA cluster analysis on 41 species in subgen.Cerasus based on thirty morphological characters
A支包括尖尾樱桃PrunuscaudataFranch.、川西樱桃PrunustrichostomaKoehne、刺毛樱PrunussetulosaBatalin、山楂叶樱桃PrunuscrataegifoliusHand.-Mazz.、偃樱桃PrunusmugusHand.-Mazz.、托叶樱桃PrunusstipulaceaMaxim.和PrunusnipponicaMatsum.。
B支包括2个小分支,B1和B2,B1包括钟花樱桃PrunuscampanulataMaxim.、高盆樱桃PrunuscerasoidesD.Don、红山樱Prunusjamasakura(Makino) Nakai、PrunussargentiiRehd.和山樱花PrunusserrulataLindl.;华中樱桃、光叶樱桃、蒙自樱桃、西南樱桃、云南樱桃、樱桃和细花樱桃组成B2。
C支包括微毛樱桃、多毛樱桃PrunuspolytrichaKoehne、康定樱桃PrunustatsienensisBatalin、锥腺樱桃、雕核樱桃、盘腺樱桃PrunusdiscadeniaKoehne、长腺樱桃PrunusdolichadeniaCardot、散毛樱桃PrunuspatentipilaHand.-Mazz.和黑樱桃PrunusmaximowicziiRupr.。
D支包括襄阳山樱桃PrunuscyclaminaKoehne、尾叶樱桃PrunusdielsianaC.K.Schneid.、浙闽樱桃PrunusschneiderianaKoehne和迎春樱桃Prunusdiscoidea(T.T.Yu & C.L.Li) Z.Wei & Y.B.Chang。E支包括欧洲酸樱桃PrunuscerasusL.和欧洲甜樱桃Prunusavium(L.) L.。Prunusapetala(Sieb.& Zucc.) Franch.& Sav.、毛瓣藏樱PrunustrichanthaKoehne、西藏樱桃CerasusyaoianaW.L.Cheng、细齿樱桃PrunusserrulaFranch.、圆叶樱桃和斑叶樱桃这6个种独立为一支;其中P.apetala、毛瓣藏樱与A支相近,西藏樱桃与E支相近。
前人对樱亚属的分类中,重齿系ser.Phyllopodium和P.serrulata复合体,以及圆叶系ser.Eumahaleb,芽鳞系ser.Cerasus都得到较好的识别。聚类的结果与前人对樱亚属分类的不同之处主要表现在3个方面:
1)Koehne和王贤荣都以萼片反折与否将樱类植物区分为两大类群。不过在对41种典型樱植物的形态聚类中,这两个类群并没有被识别出来。
2)B2支是一个由不同系或组的种组成的混杂的类群。以王贤荣的系统为例(为了方便行文,下文如无特殊说明,均以王贤荣的系统来讨论),华中樱桃属于红果系ser.Conradinia,光叶樱桃属于裂瓣系ser.Cyclaminium,蒙自樱桃属于小苞系ser.Pseudomahaleb,但这3个种首先聚为一支;而裂瓣系的西南樱桃和小苞系的云南樱桃也首先聚到一起;这2个分支的种类都属于反萼组,但是2个分支聚到一起后,首先和直萼组展萼系ser.Pseudoceraoides的樱桃聚在一起,最后才是同组的细花樱桃。
3)C支由反萼组的总状系ser.Macradenium和伞形系ser.Phyllocerasus组成。伞形系3个种(微毛樱桃、多毛樱桃、康定樱桃)和总状系的3个种(锥腺樱桃、雕核樱桃、盘腺樱桃)首先聚在一起,而后与总状系的长腺樱桃、散毛樱桃依次聚类。
对41种樱亚属植物的形态数据进行主成分分析,结果如表3所示。主成分的累积贡献率增加缓慢,前3个主成分的贡献率分别为19.095 6%、17.334 3%和11.888 2%,累积贡献率仅48.318 0%。这表明在30个形态性状中,没有明显的主成分,以主成分对樱亚属植物进行分析的话,信息会有较大的丢失,因此我们没有进行更进一步的分析。

表3 基于30个形态性状的樱亚属植物主成分分析Table 3 Principal component analysis on 41 species in subgen.Cerasus based on thirty morphological characters
3 讨论与结论
Kohne和王贤荣按萼片反折与否划分的直萼组和反萼组没有在聚类中识别出来,这表明以萼片反折与否将樱亚属划分为两个大的类群是缺少足够的依据的。
B2支与前人的分类处理较为不同,分别归属不同系乃至不同组的种类聚在一起,但这一支的种类在分类上原先就有一定的争议。《中国植物志》将襄阳山樱桃、尾叶樱桃、浙闽樱桃、光叶樱桃、樱桃、崖樱桃、西南樱桃放在裂瓣组,将云南樱桃、蒙自樱桃、细花樱桃放在小苞组sect.Pseudomahaleb,但又认为不同组的细花樱桃和樱桃形态接近,有时两种不易区分。王贤荣[2]认为在Koehne对蒙自樱桃的描述与细花樱桃极为相似,只在花序的花数量和花序长短上有所区别,但是两者的变异范围相近,因此应予以合并,这个处理没有得到Li & Bartholomew[3]的认可。《Flora of China》将光叶樱桃归并入不同组的华中樱桃中,又将西南樱桃归并入不同组的云南樱桃中。从前人对这些种的处理看来,这几个种的形态应该是比较接近的,这和形态聚类的结果相一致。结合形态聚类的结果,如果前人对这些种的处理都正确的话,那么这些种似乎更应该归并为一个种。从种的分布范围来看,樱桃分布于我国大部分地区,蒙自樱桃与细花樱桃分布在云南,西南樱桃分布于四川、云南,云南樱桃分布于云南、四川、广西,光叶樱桃分布于湖北、四川,华中樱桃分布于陕西、河南、湖南、湖北、四川、贵州、云南、广西。华中樱桃和樱桃覆盖了其余5个种的分布地,这使得7个种的分布呈现连续的状态。这7个种似乎存在是一个种的可能。但应该看到,聚类中各个种之间是存在较大距离的,以此做归并是不够科学的。王贤荣将细花樱桃并入蒙自樱桃中,但是从聚类的结果来看,相较光叶樱桃和华中樱桃,蒙自樱桃与细花樱桃的关系要更远一些。而《Flora of China》将光叶樱桃并入华中樱桃,西南樱桃并入云南樱桃的处理,在一定程度上得到聚类的支持。但是光叶樱桃在聚类中首先与蒙自樱桃聚在一起,其次才是华中樱桃。而西南樱桃与云南樱桃虽然聚在一起,但是仍然有一定的距离。这些种的处理,应该在更细致的研究后再进行考虑。
Koehne、《中国植物志》、王贤荣都将康定樱桃、微毛樱桃、多毛樱桃放在一个组或系中,雕核樱桃、锥腺樱桃、散毛樱桃、黑樱桃等总状花序的种类放在一个组或系中。形态聚类识别出了康定樱桃、多毛樱桃、微毛樱桃一支,而锥腺樱桃、散毛樱桃一支没有得到支持。锥腺樱桃、雕核樱桃、盘腺樱桃聚为一支,表明这3个种具有较近的形态特征,但这一支与康定樱桃等种形态更为接近,将锥腺樱桃与散毛樱桃放在一起的总状系或总状组没有得到支持。
形态聚类中,微毛樱桃、康定樱桃、多毛樱桃在欧氏距离2.12处聚为一支,显示了十分接近的形态,而康定樱桃和多毛樱桃在形态上更是无法区分。这3个种一直被认为是相近的种类,主要的区别是毛被和苞片边缘的腺体。《中国植物志》认为康定樱桃苞片边缘的腺体呈盘状,与微毛樱桃锥状或头状的腺体可以区别,《Flora of China》和《中国樱花品种图志》都遵循了这一处理。但是,Batalin[3]在发表康定樱桃时描述其苞片边缘的腺体是头状的,并不是盘状的,模式标本也未见苞片边缘的腺体是盘状的。嫩枝、叶和花的毛被一直被认为是多毛樱桃与近缘种的区别,而前人的描述中康定樱桃和微毛樱桃的嫩枝、叶和花的毛被是无或疏被柔毛的。从查阅标本和野外考察的情况来看,毛被的多少是存在过渡的(如图2),并且这3个种的分布是互相重叠的,将它们作为3个独立的种处理,在鉴定时不免让人感到十分困难。因此,我们建议这3个种应该合并为一个种,PrunustatsienensisBatalin发表在先,具有优先权。

图2 康定樱桃叶背与小枝条毛被的变化Fig.2 The variation of the hair on the leaves abaxial surface and branchlets of Prunus tatsienensis Batalin (图中1、2、3为采自四川省阿坝州黑水县沙石多乡居群的标本的图片,采集号为吴保欢等PHS1010、PHS1007、PHS1009;4、5、6为采自四川省阿坝州马尔康县大朗脚沟居群的标本的图片,采集号为吴保欢等PMX1014、PMX1022、PMX1017。上述标本存于CANT)
Koehne[6]发表雕核樱桃和锥腺樱桃时,认为雕核樱桃相比锥腺樱桃的叶片更狭长,腺体有较明显的凹陷。王贤荣认为两者的模式标本不存在本质上的区别,而从多数标本来看,叶较狭长、锯齿较浅、苞片较小这些性状在2个种之间是连续的,而果核的纹饰与成熟度相关,应该给予合并。形态聚类的结果显示雕核樱桃、锥腺樱桃、盘腺樱桃3个种的形态是相接近的,但是仍有一定的距离,现有证据下,仍支持《Flora of China》将雕核樱桃和锥腺樱桃作为独立的种处理。
致谢:中国科学院植物研究所标本馆杜玉芬老师和林祁研究员以及华南植物园标本馆曾飞燕老师,中国科学院昆明植物研究所标本馆王静华老师,浙江农林大学夏国华老师,美国University of California,Davis Dr.Daniel Potter,University of California,Berkeley Andrew S.Doran,Missouri Botanical Garden Dr.Libing Zhang,Dr.Zhang Liang,Herbarium of the Arnold Arboretum,Harvard University Dr.David E.Boufford,Dr.Anthony R.Brach在查阅标本过程中给予的帮助,标本查阅中也受益于中国科学院成都生物研究所植物标本馆网络标本馆(http:∥cdbi.cib.ac.cn/biology-mis/page/sample.jsf)的便利,在此一并致谢。
参考文献:
[1]俞德浚,李朝銮. 中国植物志第38卷 [M] . 北京 : 科学出版社 , 1986: 41-89.
插画要有其独特的审美价值,《彼得兔》作为全球畅销绘本经久不衰并不是意外,可以说是天时地利人和的结合。天时:工业革命彩色印刷技术的创新进步;地利:出生在当时的伦敦的作者可以接触到很多大师的优秀作品提高其鉴赏能力;人和;作者本身对绘画以及讲故事的理解和努力,形成了其独有的画风和有趣的故事。无一不是促进了《彼得兔》这一作品的诞生和畅销成为绘本中的经典之作。
[2]王贤荣. 中国樱花品种图志 [M] . 北京 : 科学出版社 , 2014.
[3]LI Chaoluan, BARTHOLOMEW B.Cerasusflora of China (Volume 9)[M] . Beijing : Science Press, 2003:404-420.
[4]POYARKOVA A I.CerasusFlora of USSR (Volume 10)[M] . Jerusalem : Israel Program for Scientific Translations Ltd, 1971 : 10, 547-575.
[5]MILLER P. Gard dict abr[M].4th ed. London : By Author , 1754:289-302.
[6]KOEHNE E.PrunusL. Plantae Wilsonianae [M] . Cambridge : Cambridge University Press , 1912:196-282.
[7]SCHNEIDER C K. Illustriertes handbuch der laubholzkunde [M] . Jena : G Fischer , 1905:602-620.
[8]REHDER A. Manual of cultivated trees and shrubs [M]. New York : The Macmillan Company, 1940.
[9]李朝銮,蒋舜媛. 杏属和樱属植物新组合 [J] . 植物分类学报, 1998, 36 (4): 80-85.
LI Chaoluan, JIANG Shunyuan. New combinations inArmeniacaMill. andCerasusJuss. (Rosaceae) [J] . Acta Phytotaxonomica Sinica, 1998, 36 (4) : 367-372.
[10]王贤荣,向其柏. 樱属植物分类研究 [J] . 南京林业大学学报, 1999, 23 (6) : 61-64.
WANG Xianrong, XIANG Qibai. Notes on some species of the genusCerasusMiq. (Rosaceae) [J] . Journal of Nanjing Forestry University, 1999, 23 (6) : 61-64.
[11]王贤荣,向其柏. 樱属植物分类订正 [J] . 南京林业大学学报, 1998, 22 (4) : 60-62.
WANG Xianrong, XIANG Qibai. Notes on some species of the genusCerasusMiq. [J] . Journal of Nanjing Forestry University, 1998, 22 (4) : 60-62.
[12]王贤荣,伊贤贵,谢春平. 武夷山樱属一新变种——武夷红樱(英文) [J] . 云南植物研究, 2007,29 (6): 616.
WANG Xianrong, YI Xiangui, XIE Chunping,Cerasuscampanulatavar.wuyiensis, a new variety of Rosaceae in Wuyi Mountain [J] . Acta Botanica Yunnanica, 2007, 29 (6) : 616.
[13]郑维列. 西藏樱属(蔷薇科)一新种 [J] . 植物分类学报, 2000, 38 (2) : 195-197.
ZHENG Weilie, A new species ofCerasus(Rosaceae) from Xizang (Tibet), China [J] . Acta Phytotaxonomica Sinica, 2000, 38 (2) : 195-197.
[14]BORTIRI E, HEUVEL B V, POTTER D. Phylogenetic analysis of morphology inPrunusreveals extensive homoplasy [J] . Plant Systematics and Evolution, 2006, 259 (1) : 53-71.
[15]BORTIRI E, OH S, JIANG J, et al. Phylogeny and systematics ofPrunus(Rosaceae) as determined by sequence analysis of ITS and the chloroplast trnL-trnF spacer DNA [J] . Systematic Botany, 2001, 26 (4):797-807.
[16]CHIN S, SHAW J, HABERLE R, et al. Diversification of almonds, peaches, plums and cherries-Molecular systematics and biogeographic history ofPrunus(Rosaceae) [J] . Molecular Phylogenetics and Evolution, 2014, 76 : 34-48.
[17]LEE S, WEN J. A phylogenetic analysis ofPrunusand the Amygdaloideae (Rosaceae) using ITS sequences of nuclear ribosomal DNA [J] . Am J Bot, 2001, 88 (1): 150-160.
[18]POTTER D, ERIKSSON T, EVANS R C, et al. Phylogeny and classification of Rosaceae [J] . Plant systematics and evolution, 2007, 266 (1/2) : 5-43.
[19]SHI S, LI J, SUN J, et al. Phylogeny and classification ofPrunussensulato(Rosaceae) [J] . J Integr Plant Biol, 2013, 55 (11) : 1069-1079.
[20]李超,李娟,张明理. 淫羊藿属植物的数量分类学研究 [J] . 西北植物学报, 2013, 33 (11) : 2339-2345.
LI Chao, LI Juan, ZHANG Mingli. A numerical taxonomical study ofEpimediumL. [J] . Acta Botanica Boreali-Occidentalia Sinica, 2013, 33 (11) : 2339-2345.
[21]任珺,陶玲. 甘肃省白刺属植物的数量分类研究 [J]. 西北植物学报, 2003, 23 (4) : 572-576.
REN Jun, TAO Ling. A numerical taxonomy of the genusNitrariafrom Gansu Province, China [J] . Acta Botanica Boreali-Occidentalia Sinica, 2003, 23 (4) : 572-576.
[22]张永增,孙文光,蒋鑫,等. 瑞香属和荛花属的数量分类研究 [J] . 广西植物, 2016, 36 (1) : 61-72.
ZHANG Yongzeng, SUN Wenguang, JIANG Xin, et al. Numerical taxonomy of the generaDaphneandWikstroemia[J] . Guihaia, 2016, 36 (1) : 61-72.
[23]郑彦超,沐先运,李研南,等. 卫矛属刺果卫矛组的数量分类学分析 [J] . 植物分类与资源学报, 2012, 34 (3) : 271-286.
ZHENG Yanchao, MU Xianyun, LI Yannan, et al. A numerical taxonomic study of theEuonymussectionEchinococcus(Celastraceae) [J] . Plant Diversity and Resources, 2012, 34 (3) : 271-286.
[24]彭焱松,陈丽,李建强. 中国栎属植物的数量分类研究 [J] . 武汉植物学研究, 2007, 25 (2) : 149-157.
PENG Yansong, CHEN Li, LI Jianqiang. Study on Numerical Taxonomy ofQuercusL. (Fagaceae) in China [J] . Journal of Wuhan Botanical Research, 2007, 25 (2) : 149-157.
[25]CHANG K S, CHANG C S, PARK T Y, et al. Reconsideration of thePrunusserrulatacomplex (Rosaceae) and related taxa in eastern Asia [J] . Botanical Journal of the Linnean Society, 2007, 154 (1) : 35-54.
[26]BATALIN A.Notae de plantis Asiaticis[J]. Trudy Imperatorskago S.-Peterburgskago Botanicheskago Sada,1895,14 (2) : 322-323.
