迁地保育林麝行为多样性及影响因素研究
2018-03-20韩红金乔佳伦蔡永华程建国付文龙孟秀祥
韩红金,乔佳伦,蔡永华,程建国,付文龙,孟秀祥,3※
(1.中国人民大学环境学院,北京 100872;2.四川养麝研究所,四川 都江堰 611830;3.山西农业大学动物科技学院,山西 太谷 030801)
动物行为指动物在一定环境条件下为了满足自身生理等需求而以一定姿势完成的一系列动作。每种动物的行为是该物种进化过程中的特定环境塑造的结果[1]。动物的行为格局与其灭绝的原因以及人们对其实施保护得难易有密切关系,圈养实践表明,任何野生动物的迁地保育、人工驯养和繁殖成功都与人们对其行为的认识和理解程度是分不开的[2]。加强动物行为格局及影响因素的研究,便于充分认识制约动物驯养和繁殖的关键因素,然后有针对性地加以干预,最终实现迁地保育种群的扩繁和养殖效益的提高。行为多样性(Behavioral diversity)基于分析动物行为流(Behaviorflow)的总行为元素和发生频次,可更直接、更包容地反映动物的行为特征[3],逐渐成为圈养野生动物及家畜行为生态学的研究热点。
麝是我国一级重点保护动物,被列为CITES附录Ⅱ物种。成年雄麝所分泌麝香是中医药的珍稀药材,具有开窍醒神、活血通络、消肿止痛等功效,用于治疗热病神昏、中风痰厥、痛肿瘰疬、跌扑伤痛等常见病、多发病和疑难病症[4]。麝的圈养和驯化作为迁地保育的重要手段以及满足麝香资源可持续供给的可行措施,正发挥着愈来愈突出的作用。目前,关于麝的行为多样性研究多集中在马麝(Moschussifanicus)。研究者们建立并定义了麝类行为谱[5],提出了马麝行为多样性的测算方法体系[6],并探究了圈养雌性马麝性活跃度与行为多样性的关系[7]以及行为频次与生殖潜能之间的相关[8],并初步建立了圈养雄性马麝繁殖性能的行为判别指标[9]。
林麝(Moschusberezovskii)是麝属动物中分布最广泛的麝种,也是最早开始人工圈养的麝种之一。截至2011年底,全国圈养麝群中83%~87%为林麝[10,11]。关于林麝的行为生态研究多集中在生境选择[12,13]、种群动态[14]、分娩[15]、哺乳[16]和等级序位[17]等方面,迄今尚未系统研究圈养林麝的行为多样性及影响因素。对圈养林麝开展行为多样性格局的量化研究,探究性别、年龄、雌麝繁殖成效和雄麝泌香量等因素与其行为多样性的关系,相关结果有助于濒危林麝的成功驯养、迁地保育和可持续利用。
1 研究地区自然概况
本研究在四川养麝研究所所属的马尔康林麝繁育场(以下简称麝场)进行。马尔康麝场位于四川省阿坝藏族羌族自治州,地处高原,空气稀薄;辐射强烈,年日照时数2 214h,日照率50%;气候高寒,年均温度为5℃~9℃,昼夜温差大;年均降雨760mm,多夜雨。马尔康的森林资源十分丰富,是国家重要木材生产基地;动物资源也十分丰富,大型草食性动物主要有牛羚(Budorcas taxicolor)、藏原羚(Procapra picticaudata)等。
2 研究方法
2.1 样本动物及分组
四川马尔康麝场现有存栏圈养林麝700头左右,分别饲养在9个饲养区当中,每个饲养区中有5~8个并排圈舍,每个圈舍由4~7个小舍(每个小舍面积2m2~4m2,混凝土地面基底)及1个公用的活动场(面积100m2)组成,活动场中央位置是水泥凉棚(6m2左右),凉棚四周设置2个饮水槽和2个料槽。活动场为硬化的泥质基底,有天然杂草植被和榆树等落叶乔木。每个饲养区固定由1名专职饲养员负责饲喂和日常管理。饲料提供为每日2次,在相对固定时间(夏季:06:00和19:00;冬季:08:00和17:00)分别投饲干、湿树叶和精饲料,并更换饮水。
本研究样本106头,其中雄麝36头,雌麝70头。林麝按照发育时期可分为亚成体(1岁,59头)、成体(2岁~8岁,32头)和老年个体(≥9岁,7头);依据2016年度的繁殖情况,雌麝分为亚成体雌麝(1岁,31头)和育龄雌麝(≥2岁,27头,其中繁殖成功雌麝14头、繁殖失败雌麝13头);根据当年麝香分泌量,雄麝可分为高泌香量个体(≥10g,9头)、较高泌香量个体(5g~10g,14头)、低泌香量个体(≤5g,15头)和未泌香亚成体(21头)。
2.2 数据收集
参照戚文华等[18]的夏季林麝行为谱和孟秀祥等[6]的麝类行为谱,结合取样现场的实际观察,构建圈养林麝的夏季取样行为谱。行为观察于圈养林麝的早、晚活动高峰期(06:00~08:00;18:00~20:00)进行,确保行为观察和取样的全面性及代表性。首先依据耳号和个体特征做到个体识别,用10 22°望远镜协助肉眼进行焦点取样(Focal sampling),记录10min内各行为型的行为发出者、行为对象、行为频次和行为持续时间等变量。如果林麝在10min内走出视野,此次记录中断。
2.3 数据整理及分析方法
计算样本行为流的绝对行为多样性指数H(Absolute diversity index)及特定亚总体的最大行为多样性指数Hmax,再计算相对多样性指数r(Relativediversity index)[19]。本研究用行为频次参与计算:
H=∑(Pilog21/Pi),Pi=fi/∑f。其中,fi为样本行为流里第i种行为的发生频次,f为每次取样观察记录的总频次;
Hall=log2N,其中,N为圈养林麝的行为谱所含行为型个数;Hall代表圈养林麝的最大行为多样性指数,即行为谱的N种行为均概率出现;
r=H/Hall,即相对行为多样性指数,反映了特定环境下或特定类群的具体行为多样性指数H和Hall的相对关系。
整理行为数据,采用Kolmogorov-Smirnov Test检验数据正态性,如数据呈正态分布,则用ANOVA和独立样本T检验检测性别、年龄等因素对行为多样性的效应;如数据及标准化后的数据均呈非正态分布,选用Mann-WhitneyU和Kruskal-WallisTest分析各因子对林麝行为多样性的影响。设定差异显著性标准为P=0.05,数据分析通过SPSS19.0进行。
3 结果
3.1 行为多样性指数分布
在马尔康养麝场的调查共记录到43种行为,行为多样性指数r呈正态分布(K-S:Z=1.176,P=0.146>0.05,n=277)。
3.2 性别对行为多样性的效应
性别对林麝行为多样性的影响不具有显著差异(ANOVA:df=1,F=1.215,P=0.271),雄麝行为多样性(r=0.26±0.016,n=94)略低于雌麝(r=0.29±0.012,n=175)。
3.3 年龄对行为多样性的影响
年龄对林麝行为多样性的影响不显著(ANOVA,df=9,F=0.906,P=0.521)。分组计算后(图1)发现,亚成体行为多样性(r=0.316±0.036,n=137)高于成体(r=0.301±0.013,n=100)和老年个体(r=0.295±0.017,n=21),但各年龄组行为多样性的递减趋势不显著(ANOVA,df=2,F=0.032,P=0.968)。
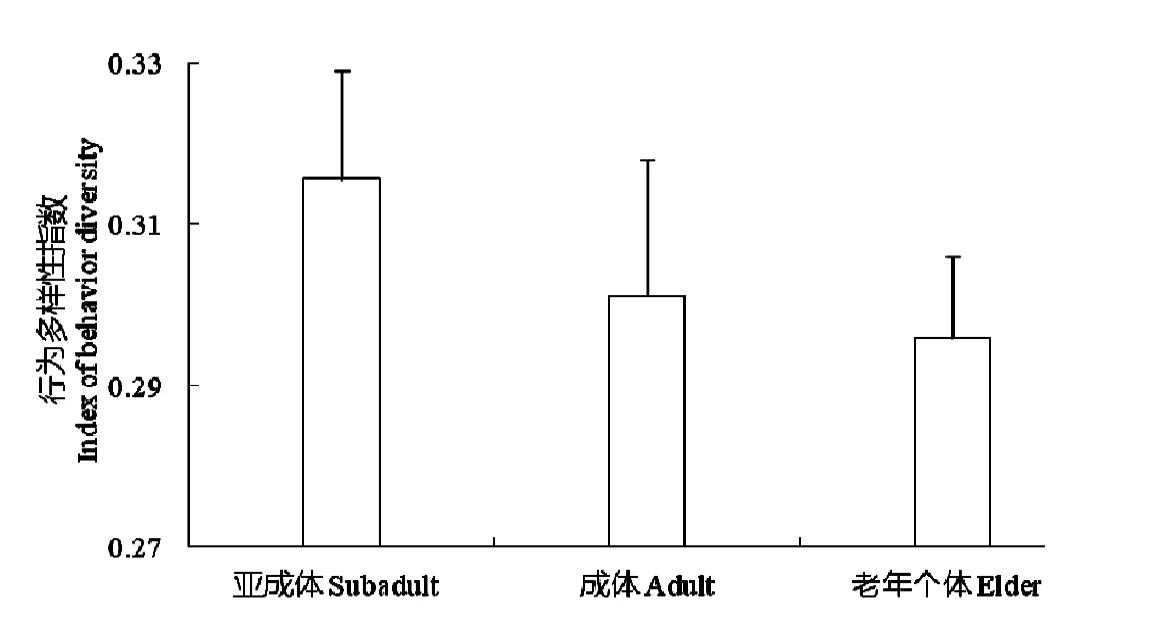
图1 林麝行为多样性的年龄组间变化Fig.1 Behavioral diversity of captive forest musk deer among age groups
3.4 雌麝繁殖状况与行为多样性的关系
2016年繁殖成功雌麝的行为多样性(r=0.28±0.053,n=14)略低于繁殖失败雌麝(r=0.33±0.035,n=13),差异不显著(t=0.785,F=3.89,P=0.44)。
繁殖龄雌麝的行为多样性(r=0.30±0.031,n=27)小于不参与繁殖的亚成体雌麝(r=0.32±0.030,n=31),差异不显著(t=0.496,F=0.252,P=0.622)。
3.5 雄麝泌香情况与行为多样性的关系
结合当年泌香数据发现,雄麝的泌香量与行为多样性相关不显著(ANOVA,df=2,F=0.544,P=0.587)。高泌香量雄麝的行为多样性(r=0.28±0.046,n=9)最低,较高泌香量雄麝的行为多样性最大(r=0.32±0.015,n=14),差异不显著(t=0.446,F=0.019,P=0.660)。
4 讨论
圈养对动物来说,就是一种圈禁状态,这种状态持续的时间影响动物的行为多样性发育[20],年龄可很好地代表其被实际圈养的时间。本研究表明,亚成体行为多样性最高,之后随年龄的增加,行为多样性呈下降趋势,老年个体的行为多样性最低,这与Stolba等[19]对圈养猪(Susscrofadomestica)和孟秀祥等[6]对圈养马麝的研究结果一致。这是因为动物所处的发育阶段及幼年生活史等个体生理特征会对动物的行为多样性产生效应[21]。随着年龄的增长,圈养动物对于贫乏环境的耐受程度渐趋极限,刻板行为发育较多,行为多样性降低,而且随着个体的衰老,机体功能衰退,活跃度也下降,这也是造成老年个体行为多样性减少的重要原因。
由于性别不同,动物内分泌等生理特征也有所不同,动物在圈养环境下行为模式的改变也存在性间变化,故不同性别的圈养动物的行为多样性格局也有差异[22]。对于有蹄类动物来说,不同性别个体因体型差异而产生了不同的能量需求,导致此类动物在行为和时间分配上的差异[23]。本研究显示,在夏季的林麝行为多样性格局中,雄麝行为多样性指数要小于雌麝的行为多样性指数,除了生理特征差异,也可能与此次观察时间正值雄麝泌香期末期和换毛期有关。雄麝泌香期间饮食量相对减少[24],同时麝香分泌消耗较多能量,刚刚结束泌香的雄麝展现出更多的静卧、反刍和安静站立[25]等旨在恢复体能的行为,较早结束泌香的雄麝则开始进行换毛。另一方面,此次观察取样的雌麝绝大多数为亚成体或繁殖失败的成体,体能较充沛,其行为展现较为多样。
对雌性马麝的研究发现,在非交配季节,性活跃雌麝警觉性较高,用于站立凝视和环境探究等行为的时间显著多于性不活跃雌麝,运动时间显著地少于性不活跃雌麝,造成其行为多样性小于性不活跃雌麝[8]。而那些性活跃雌麝多为当年繁殖成功者,在非繁殖季节正处于哺乳期[15,26]。本研究也发现,2016年繁殖成功雌麝的行为多样性低于繁殖失败雌麝,这与雌麝的繁殖和哺乳等母性行为有关,表明生产繁殖会对林麝的行为多样性产生影响。同时,育龄雌麝的行为多样性低于亚成体雌麝,印证了生产繁殖对于雌麝的行为多样性展现有一定的抑制作用。
对驯养麝类而言,其麝香产量主要受分布区域、个体来源[27]、年龄、体质及饲养管理模式等影响[28,29],也受泌香盛期的长短[30]和整个泌香期雄性激素的影响[31,32]。本研究通过对比发现,泌香雄麝的行为多样性高于未泌香的亚成体,但差异没有达到显著水平。结合当年泌香数据发现,雄麝的行为多样性不能显著反映该个体泌香量的多少,原因除了影响雄麝泌香的因素较多外,还可能与泌香数据有限和饲养管理方式等有关,需要进一步的研究。
综合以上分析可以发现,林麝的行为多样性受到性别、年龄以及繁殖等多种因素影响,加强行为多样性研究有助于改善动物福利,并且为麝类动物的迁地保育工作提供理论依据。
[1]蒋志刚,马克平,韩兴国.保护生物学[M].杭州:浙江科技出版社,1997:43-70.
[2]蒋志刚.动物行为原理与物种保护方法[M].北京:科学出版社,2004:254-279.
[3]Wemelsfelder F,Haskell M,Mendl M T,et al.Diversity of behavior during novel object tests is reduced in pigs housed in substrate-impoverished conditions[J].Animal.Behaviour,2000,60:385-394.
[4]郭经.人工麝香研究进展[J].中国医学科学院学报,2014,36(6):577-580.
[5]孟秀祥,冯金朝,周宜君,等.麝类行为谱的初步建构及行为型的描述性定义[J].四川动物,2007,26(1):46-50.
[6]孟秀祥,杨奇森,冯祚建,等.兴隆山圈养马麝的行为多样性[J].动物学报,2006,52(6):1026-1033.
[7]孟秀祥,杨奇森,冯祚建,等.圈养雌性马麝性活跃度与行为的关系[J].经济动物学报,2007,11(1):30-33.
[8]Meng X X,Perkins G,Yang Q S,et al.Relationship Between Behavioral Frequency and Reproductive Potential of Female Alpine Musk Deer in Captivity[J].Agricultural Sciences in China,2008,7(12):1497-1502.
[9]孟秀祥,杨奇森,冯祚建,等.甘肃兴隆山保护区圈养雄性马麝繁殖性能的行为判别[J].动物学研究,2007,28(2):149-154.
[10]李林海,黄祥云,刘刚,等.我国麝养殖种群现状及其养殖业发展的分析[J].四川动物,2012,31(3):492-496.
[11]吴民耀,王念,惠董娜,等.林麝保护的现状及研究进展[J].重庆理工大学学报:自然科学版,2011,25(1):34-39.
[12]胡忠军,王淯,薛文杰,等.紫柏山自然保护区林麝冬季生境选择[J].河南大学学报:自然科学版,2006,36(1):70-74.
[13]郭建,程晓峰,巨云为,等.冶勒自然保护区林麝对生境选择研究[J].应用与环境生物学报,2001,7(2):183-185.
[14]胡忠军,王淯,薛文杰,等.陕西凤县林麝家庭养殖:历史,现状与繁殖特性[J].四川动物,2007,26(4):948-951.
[15]杜卫国,盛和林.圈养林麝的分娩及一日龄行为[J].浙江师范大学学报:自然科学版,1996,(S1):84-86.
[16]杜卫国,盛和林.林麝哺乳期的时间分配和行为研究[J].兽类学报,1998,18(1):21-26.
[17]徐正强,徐宏发.饲养林麝的种群特征和幼麝的存活研究[J].兽类学报,2003,23(1):17-20.
[18]戚文华,汪小方,杨承忠,等.圈养林麝夏季昼夜活动节律及时间分配[J].四川动物,2013,(1):19-22.
[19]Stolba A,Baker N,Wood-Gush D G M.The characterisation of stereotyped behaviour in stalled sows by informational redundancy[J].Behaviour,1983,87(3):157-182.
[20]Haskell M,Wemelsfelder F,Mendl M T,et al.The effect of substrate-enriched and substrate-impoverished housing environments on the diversity of behaviour in pigs[J].Behaviour,1996,133(9):741-761.
[21]Wemelsfelder F,Haskell M,Mendl M T,et al.Diversity of behaviour during novel object tests is reduced in pigs housed in substrate-impoverished conditions[J].AnimalBehaviour,2000,60(3):385-394.
[22]Schoenecker B,Heller K E,Freimanis T.Development of stereotypies and polydipsia in wild caught bank voles(Clethrionomys glareolus)and their laboratory-bred offspring:Is polydipsia a symptomofdiabetes mellitus[J].Applied animal behaviour science,2000,68(4):349-357.
[23]Ruckstuhl K E.Foraging behaviour and sexual segregation in bighorn sheep[J].Animal Behaviour,1998,56(1):99-106.
[24]盛和林,刘志霄.中国麝科动物[M].上海:科学技术出版社,2007:1-158.
[25]白康,任战军,王永奇,等.林麝泌香期性激素变化及其与泌香量的关系[J].中国兽医学报,2013,33(6):956-962.
[26]张保良.麝的驯养[M].北京:农业出版社,1979:20-43.
[27]李红亮,黎勇,周密,等.驯养马麝麝香产量及其影响因素[J].中国农业科学,2011,45(3):522-528.
[28]Homes V.On the scent:conserving musk deer:the uses of musk and Europe's role in its trade[M].Brussels:Traffic Europe,1999:1-30.
[29]Zhang B L.Musk-deer:their capture,domestication and care according to Chinese experience and methods[J].Unasylva(FAO),1983,35:16-24.
[30]Cheng J G,Luo Y,Qiao M P,et al.The factors affecting musk secretion quantity in Moschus berezovskii[J].Special Wild Economic Animal and Plant Research,2002,(3):15-18.
[31]Bai K,Ren Z J,Wang Y Q,Li F R,et al.An association with gonadal hormone level and musk yield in Moschus berezovskii during musk-secreting period[J].Chinese Journal of Veterinary Science,2013,(6):30.
[32]Jie H,Feng X L,Zhao G J,et al.Research progress on musk secretion mechanism of forest musk deer[J].China journal of Chinese material medical,2014,39(23):4522-4525.
