水分亏缺对滴灌柑橘光合和产量及水分利用效率的影响
2018-03-01张效星崔宁博胡笑涛龚道枝
张效星,樊 毅,贾 悦,崔宁博,4※,赵 璐,胡笑涛,龚道枝
0 引 言
柑橘(Citrus reticulata Blanco)是中国最主要的水果之一。截止2016年,中国柑橘种植面积228.8万hm2、总产量达到2 944.04万t,均排名世界第一;四川省是中国柑橘生产大省,柑橘栽培面积25.32万hm2,年产量达292.9万 t。但近年来频发的季节干旱与粗放式水肥管理导致其品质较低、产量不稳定。滴灌是一种有效的节水灌溉方式,灌溉水的有效利用率可达 90%,已有大量研究证明滴灌是果树较适宜的灌溉方式,具有显著的节水增产、提质增效的良好效果[1-2]。
亏缺灌溉是基于根冠通信理论[3]、生长冗余理论[4]、生长补偿效应[5]、气孔调节理论[6]和作物有限水量最优分配理论[7]发展起来的生理节水技术。研究表明,亏水会导致作物叶片气孔关闭、光合速率(Pn)下降,引起作物体内的激素、可溶性物质含量的变化,进而使其生长受到抑制[3];当亏水度加剧时,作物叶水势与叶面积均呈减小趋势,并抑制新生梢生长,且亏水时间越长影响越显著[8]。Thomas等[9]表明,亏水会促进苹果树根部脱落酸(absicisic acid,ABA)合成,Panigrahi等[10]发现,轻度亏水处理下柑橘叶片Pn较对照仅降低0.03 μmol/(m2·s),叶片瞬时水分利用效率(instaneous water use efficiency of leaf,WUEi)提高 16%,Pn的提高和蒸腾速率的降低是 WUEi提高的主要原因。Chang等[11]报道,亏水处理2周后,盆栽金钱橘光合指标均显著降低,主要原因是水分亏缺造成叶片气孔开度降低。Gasque等[12]在Navelina橘的亏水试验中发现,轻度亏水处理在节水12%~27%的同时并没有降低产量,并指出当茎水势(Ψst)低于-2.0 MPa时亏水将对产量、品质带来不利影响。Roccuzzo等[13]发现在全生育期进行亏水处理可使柑橘WUEi提高13%~15%。Tejero等[14]发现节水超过1 000 m3/hm2(约3 m3/棵)时,亏水处理会降低柑橘产量、影响果实品质。
作物受到水分胁迫时叶片光合速率减小,通常认为光合能力降低的原因包括气孔限制与非气孔限制[15]。光合气孔分析理论认为气孔限制的一个重要判据是胞间CO2浓度(Ci)的降低方向,并非 Ci降低幅度[16]。徐俊增等[17]在水稻试验中发现亏水后叶片气孔限制值与非气孔限制值呈相反的变化规律。杨泽粟等[18]将春小麦光午休现象的主要原因归因于气孔限制。然而,影响作物光合的主要因素是气孔限制还是非气孔限制不仅与作物基因型[19]有关,还与土壤含水率、所在生育期[20]、水分胁迫持续时长[21]等密切相关,不同品种的同一作物对水分胁迫的响应不同,气孔与非气孔限制之间的亏水度转变阈值也不同。目前国内关于柑橘光合的气孔与非气孔限制的研究少见报道。
本文目的是研究滴灌水分亏缺对柑橘叶片光合特性、产量及水分利用效率的调控效应,揭示滴灌水分亏缺对柑橘叶片光合特性的影响机制,为深入理解滴灌水分调控下柑橘叶片气孔响应规律与水分高效利用机制提供理论基础。
1 材料与方法
1.1 研究区概况
试验于2016年4月—2017年1月在四川省成都市蒲江县长林农场(海拔544 m,30.32°N、103.43°E)进行,试验地土壤类型为黄壤土,平均容重为1.45 g/cm3,田间持水量为26.13%,土壤全氮质量分数为0.69 g/kg,有机质质量分数为9.32 g/kg,有效磷质量分数为4.6 mg/kg,速效钾质量分数为 130 mg/kg。试验区位于成都平原腹地,属亚热带湿润季风气候区,年平均气温16.3 ℃,平均相对湿度87.84%,年平均风速1.67 m/s,年平均降水量1 196.2 mm。
1.2 试验设计及过程
试验以7 a生“不知火”柑橘为试材,株高约2.8~3.0 m,径粗约10 cm,株行距为2.5 m×3.0 m,主要生育期划分为抽梢开花期(4月上旬—下旬,Ⅰ期)、幼果期(5月上旬—6月下旬,Ⅱ期)、果实膨大期(7月上旬—11月上旬,Ⅲ期)和果实成熟期(11月中旬—次年1月下旬,Ⅳ期)。
按照土壤含水率的下限来控制灌水量可以达到较为准确的精度,但在指导实际的山丘区果树种植灌溉时,农户缺乏相应的仪器设备,而试验地土壤质地比较均一、土壤空间变异性较低,考虑到实际应用,本文通过控制灌水量来进行调亏灌溉。试验分别在Ⅲ期、Ⅳ期设置轻度亏水、中度亏水、偏重度亏水和重度亏水处理,灌水量分别为对照(CK)的85%(LD)、70%(MD1)、55%(MD2)和40%(SD),设置1个对照处理(CK),灌水周期约为7~10 d,对照处理灌水量为100%,相当于90%的田间持水量。4个生育期各处理灌水量见表1。试验用肥均按照安杰农业有限公司的水溶肥配方施加,每次施肥均采用总干管进口处的水肥一体化设备操作。抽梢开花期N、P、K肥各施42.75 kg/hm2,幼果期分别施N、P、K肥19.50、55.50、19.50 kg/hm2,果实膨大期分别施N、P、K肥250.95、259.35、395.55 kg/hm2,果实成熟期分别施N、P、K肥84.00、203.70、235.50 kg/hm2。
试验采用低压滴灌水肥一体化系统,滴灌采用以色列进口Dripnet PC压力补偿式滴灌管,沿地面布设,滴灌管环形布置,每条滴灌管等间距内嵌20个迷宫式压力补偿滴头,流量为1.6 L/h,试验每2棵树1个处理小区,每个处理2条毛管,支管进口设置阀门,且毛管流量为32.0 L/h;干管垂直支管布置,并在试验区进口设置精量水表。进行亏水处理试验组的试验时进行时间控制,当灌水时间乘以灌水流量等于灌水量时,关闭该处理的阀门,直至所有处理灌水结束。试验各小区间用埋深1 m的硬塑料薄膜相隔,防止处理间侧向水分运移的影响,以3.6 m高透光薄膜电动防雨棚作为避雨设施,试验区地下水埋深12 m,地下水对试验无影响。其他田间管理措施均相同。
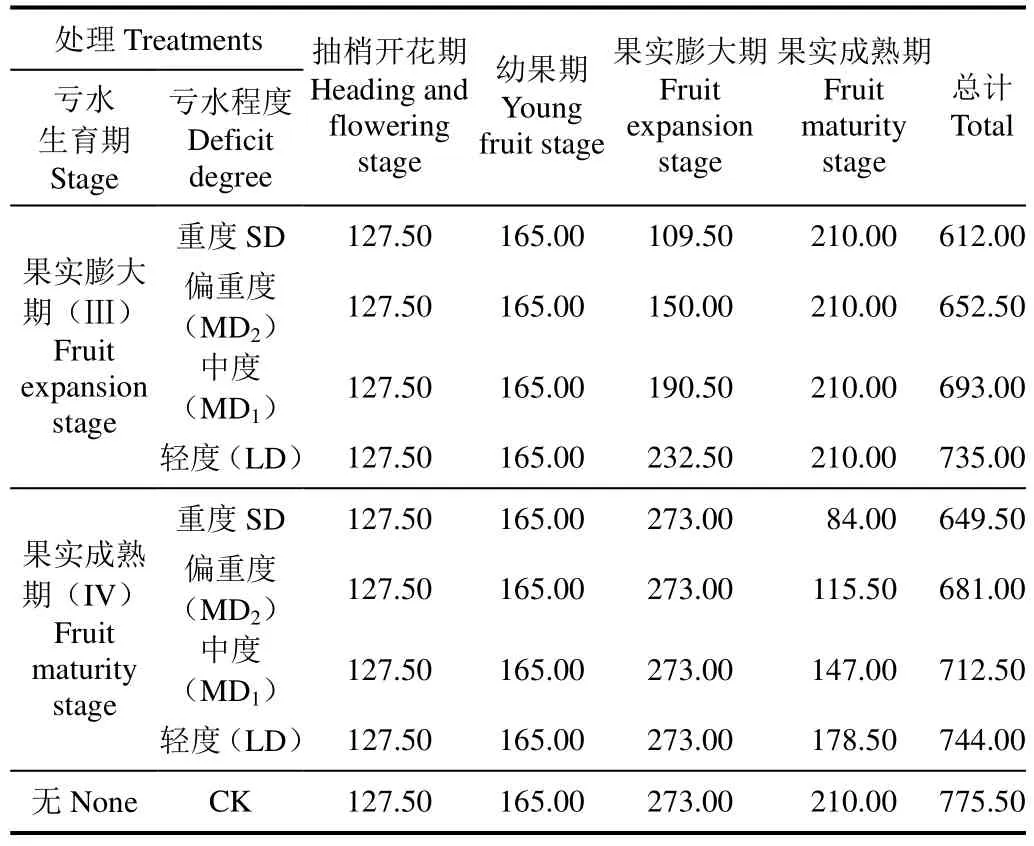
表1 滴灌试验各处理灌水量Table 1 Irrigation amount for treatments during drip experiment m3·hm-2
1.3 指标测定
分别在Ⅲ、Ⅳ期亏水第6、7天(8月12日、11月15日,天气晴朗)08:00—18:00使用全自动便携式光合仪(LCPro-SD,英国ADC)测定叶片光合日变化、水汽压差(Δe)、光合有效辐射(photosynthetically active radiation,PAR)。通过试验基地小型自动气象站BO8-HOBO测定大气温度(Ta)和相对湿度(relative humidity,RH)。
叶片光合特性的测定方法:各处理选择具有代表性的4片叶子,测定其光合特性,包含胞间CO2浓度(Ci)、蒸腾速率(Tr)、气孔导度(gs)及净光合速率(Pn)。由于gs降低,进入气孔的CO2减少,不能满足光合作用的要求,用气孔限制值(Ls)来表示光合作用的气孔限制(Ls=1-Ci/Ca,Ca表示大气CO2浓度);由于叶片温度增高,叶绿体活性与Rubisco活性降低、RuBP羧化酶再生能力降低,导致叶片光合作用能力降低,非气孔限制值用Ci/gs来计算[17]。
耗水量采用水量平衡法计算:
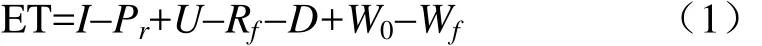
式中ET为果树耗水量,mm;I为灌水量,mm;Pr为有效降雨量,mm;U为地下水补给量,mm;Rf为地表径流量,mm;D为深层渗漏量,mm;W0和Wf分别为时段初和时段末的土壤储水量,mm;因为试验在避雨棚内进行,因此无降雨和径流量产生,忽略地下水补给和深层渗漏,故 Pr=0,Rf=0,U=0,D=0。因此,式(1)可以简化为:

叶片瞬时水分利用效率计算公式为:

式中WUEi为瞬时水分利用效率,μmol/mmol;Pn为净光合速率,μmol/(m2·s);Tr为蒸腾速率,mmol/(m2·s)。
虽然本研究的结论有前人的研究作为依据,但仍然存在一些不足。首先,本研究为回顾性研究,样本的同质性以及对照的选择存在一定的局限性,一定程度上会影响结果的信度及效度;其次,本研究样本量有限,一定程度上会影响结果的准确性;最后,本研究采用的术后椎体旋转评价方式并未得到广泛应用,也会使得结果有一定偏差。
羧化速率计算公式为:

式中 CE 为羧化速率,mol/(m2·s);Ci为胞间CO2浓度,10-6。
灌溉水利用效率(WUEI)为产量与灌水量的比值,kg/m3;作物水分利用效率(WUEET)为产量与耗水量的比值,kg/m3。
各处理平均值之间的差异显著性采用SPSS19.0软件进行单因素方差(analysis of variance,ANOVA)分析,文中图表在Microsoft Excel2007绘制。
2 结果与分析
2.1 环境因子日变化
2个生育期柑橘主要环境因子日变化如图1所示。由图1可知,Ⅲ、Ⅳ期日Ta呈单峰状,峰值出现在12:00,日均Ta分别为32、16 ℃;Ⅲ、Ⅳ期日均Δe分别为4.30×102Pa、3.21×102Pa;PAR呈单峰型日变化趋势,Ⅲ、Ⅳ期日均值分别为 751.2、247.12 μmol/(m2·s);RH 均呈下凹状变化趋势,Ⅲ、Ⅳ期日均值分别为80.12%、88.33%。
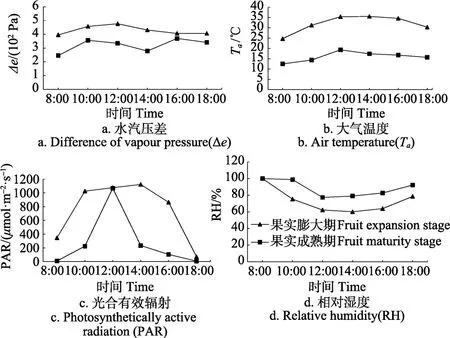
图1 主要环境因子日变化Fig.1 Diurnal variation of main environmental factors
2.2 滴灌水分亏缺光合特性日变化
图2 a和图2b为果实膨大期和果实成熟期不同水分亏缺处理柑橘叶片净光合速率(Pn)日变化。Pn日变化呈双峰状,12:00出现“午休”现象,不同处理Pn随亏水度的加剧而降低。Ⅲ、Ⅳ期CK的日均Pn分别达3.94、2.85 μmol/(m2·s),Ⅲ-MD1、Ⅲ-LD、Ⅳ-MD1和Ⅳ-LD 处理日均Pn与CK无显著差异(P>0.05),但Ⅲ-MD2、Ⅲ-SD、Ⅳ-MD2、Ⅳ-SD处理日均Pn下降18.6%~31.4%(P<0.05),Ⅲ期较Ⅳ期 Pn显著提高 26.28%~43.50%(P<0.05),变化趋势与水分胁迫下梭梭[15]、怪柳[22]的研究结果一致。
图2c和图2d为果实膨大期和果实成熟期滴灌不同水分亏缺处理柑橘叶片蒸腾速率(Tr)日变化。Tr日变化呈单峰状,Ⅲ、Ⅳ期 CK日均 Tr分别为 1.75、2.42 mmol/(m2·s)。Ⅲ-MD1、Ⅲ-LD、Ⅳ-MD2、Ⅳ-MD1、Ⅳ-LD处理 Tr与CK无显著差异(P>0.05),Ⅲ-SD、Ⅳ-SD处理Tr比CK处理分别降低20.9%、23.7%(P<0.05)。
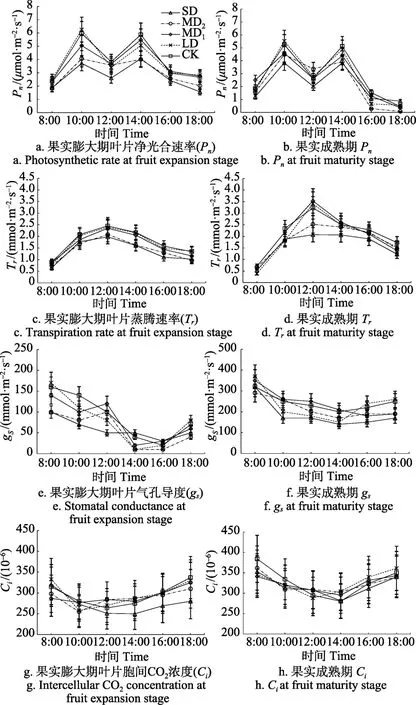
图2 不同处理柑橘叶片光合参数日变化Fig.2 Diurnal variation of citrus leaf photosynthetic parameters for different treatments
图2 e和图2f为果实膨大期和果实成熟期滴灌不同水分亏缺处理柑橘叶片气孔导度(gs)日变化。gs日变化呈下凹状,不同亏水处理gs在10:00—12:00、14:00—16:00变化趋势存在明显差异,与10:00相比,12:00时Ⅲ-SD、Ⅲ-LD、CK处理gs减小27.27%~28.57%,而Ⅲ-MD1、Ⅲ-MD2处理gs分别增加25.00%、20.00%(P<0.05);与14:00相比,16:00时除Ⅳ-MD1处理 gs减小14.29%外,其余处理均增大。Ⅲ、Ⅳ期 gs最大值分别为 150、350 mmol/(m2·s),主要是由于Ⅲ期气温较高,叶片通过减小gs避免过多的水分流失。
图2g和图2h为果实膨大期和果实成熟期滴灌不同水分亏缺处理柑橘叶片胞间 CO2浓度(Ci)日变化。Ci日变化呈“V”型,日变化范围在250×10-6~340×10-6,当日 Ca变化范围为 350×10-6~480×10-6,Ca始终高于 Ci,Ci的日变化规律与武阳等[19]试验结果一致。然而,各亏水处理间Ci变化趋势存在明显差异:与10:00相比,12:00时Ⅲ-MD2、Ⅲ-MD1、Ⅲ-LD处理Ci增大2.53%~11.37%,Ⅲ-SD、CK分别下降 8.06%、5.71%,除Ⅲ-MD2外其余处理差异均达显著水平(P<0.05),Ⅳ-LD、CK分别下降10.18%、11.77%,Ⅳ-SD、Ⅳ-MD2、Ⅳ-MD1无显著差异(P>0.05)。

表2 滴灌水分亏缺处理对柑橘果实膨大期和成熟期光合特性的影响Table 2 Effect of water deficit on citrus’s photosynthesis during fruit expansion stage and fruit maturity stage under drip irrigation
2.3 滴灌水分亏缺气孔限制值与净光合速率、蒸腾速率和瞬时水分利用效率的关系
图 3为不同生育期滴灌水分亏缺处理柑橘叶片气孔限制值(Ls)和非气孔限制值(Ci/gs)日变化。由图3a、3c可知,Ⅲ期各亏水处理后Ls呈08:00—10:00逐渐增大、10:00—12:00缓慢减小、14:00—18:00持续增大的变化趋势,Ci/gs呈08:00—12:00保持较低水平、12:00后迅速升高的变化趋势,二者基本呈相反的变化规律,与CK相比,各亏水处理后 Ls和 Ci/gs随亏水度的加剧分别增大1.24%~20.11%、2.17%~27.47%。结合图2分析可得,08:00—10:00间,各亏水处理的光合指标均表现为 Pn增大,gs减小,Ci降低,Ls增大,Ci/gs保持较低水平,说明气孔限制是该时段影响柑橘叶片Pn的主要因素;10:00—12:00 Pn出现“光午休”现象,各亏水处理的光合指标变化存在明显差异,其中Ⅲ-MD2、Ⅲ-MD1、Ⅲ-LD处理的 gs、Pn、Ls均降低,Ci增大,说明气孔限制是“光午休”现象的主要影响因素,Ⅲ-SD处理的 Ci、Ls、Pn均减小,gs、Ci/gs均增大,说明非气孔限制是Ⅲ-SD处理出现“光午休”现象的主要影响因素;与12:00相比,14:00时Ⅲ-MD2、Ⅲ-MD1、Ⅲ-LD处理Ci/gs为12:00的2.13~3.94倍(P<0.05),非气孔因素占主导地位;同理可知,14:00以后非气孔限制是各亏水处理下影响Pn的主要因素。综上所述,Ⅲ期各亏水处理不同时段影响Pn的主要因素是:08:00—10:00各处理Pn的主要影响因素均为气孔限制,10:00—12:00Ⅲ-LD、Ⅲ-MD1、Ⅲ-MD2处理 Pn减小的主要原因是气孔限制,Ⅲ-SD处理Pn降低的主要原因是非气孔限制,12:00—18:00各处理 Pn的主要影响因素为非气孔限制。

图3 不同处理柑橘叶片气孔限制值与非气孔限制值日变化Fig.3 Diurnal variation of stomatal and non-stomatal limits of citrus leaf for different treatments
图3 b、d显示,Ⅳ期亏水处理后Ls日变化呈08:00—14:00保持较低水平、14:00后逐渐升高、16:00达到最大的变化趋势,Ci/gs大致呈“M”型。与CK相比,Ⅳ-SD处理日均Ls、Ci/gs分别提高13.07%、33.99%(P<0.05),Ⅳ-MD1、Ⅳ-LD处理无显著变化(P>0.05)。Ⅳ期各亏水处理影响Pn的主要因素为:08:00—10:00各亏水处理均为气孔限制,10:00—12:00间Ⅳ-LD、Ⅳ-MD1处理为气孔限制,Ⅳ-MD2、Ⅳ-SD处理为非气孔限制,12:00—14:00各亏水处理均为气孔限制,14:00—18:00均为非气孔限制。
图 4为不同生育期滴灌水分亏缺处理下柑橘 Ls与Pn、Tr、WUEi的日变化。由图可知,Ls与Pn并无直接的线性关系,但Tr随Ls的增加而降低,说明气孔限制对于 Tr的影响更加显著。从 Ls对 WUEi的影响来看,Ⅲ期 WUEi随 Ls的增加而增大,当Ⅲ期 Ls>0.4、Ⅳ期Ls>0.22时,Pn较低,导致WUEi大幅降低,Ⅲ期Ls=0.4大于Ⅳ期Ls=0.22的原因可能是随着叶龄的增大,叶片酶活性下降。
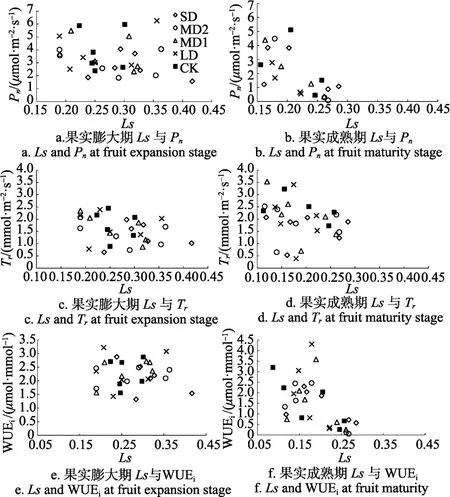
图4 柑橘Ls与Pn, Tr, WUEi的关系Fig.4 Relationship between Ls and Pn, Tr, WUEi of citrus
2.4 滴灌水分亏缺产量及水分利用效率
表3为不同生育期滴灌水分亏缺处理对柑橘耗水量、产量及水分利用效率的影响。由表3可知,CK的耗水量达852.60 mm,灌水量为880.95 mm,产量达38 539.20 kg/hm2,与 CK相比,Ⅲ期各亏水处理的耗水量降低 10.7%~36.3%,灌水量降低33.5%~8.4%,其中Ⅲ-SD、Ⅲ-MD2处理的产量分别降低38.1%、11.8%(P<0.05),而Ⅲ-LD、Ⅲ-MD1处理的产量无显著差异(P>0.05),同时Ⅲ-LD、Ⅲ-MD1处理的WUEET分别提高13%、26%,WUEI分别提高 11%、22%(P<0.05);Ⅳ期各亏水处理的耗水量降低12.2%~6.7%,灌水量降低17.2%~4.3%,其中Ⅳ-SD、Ⅳ-MD2、Ⅳ-MD1处理产量分别降低 42.7%、32.2%和23.0%(P<0.05),而Ⅳ-LD处理产量无显著差异(P>0.05),同时Ⅳ-LD处理的WUEET、WUEI分别提高9.5%、6.87%(P<0.05)。因此,在柑橘果实膨大期和果实成熟期适度的水分亏缺(Ⅲ-LD、Ⅲ-MD1、Ⅳ-LD)可在保证柑橘产量不显著变化的条件下,节约灌溉用水6.87%~22%的同时,有效提高WUEET9.5%~26%。
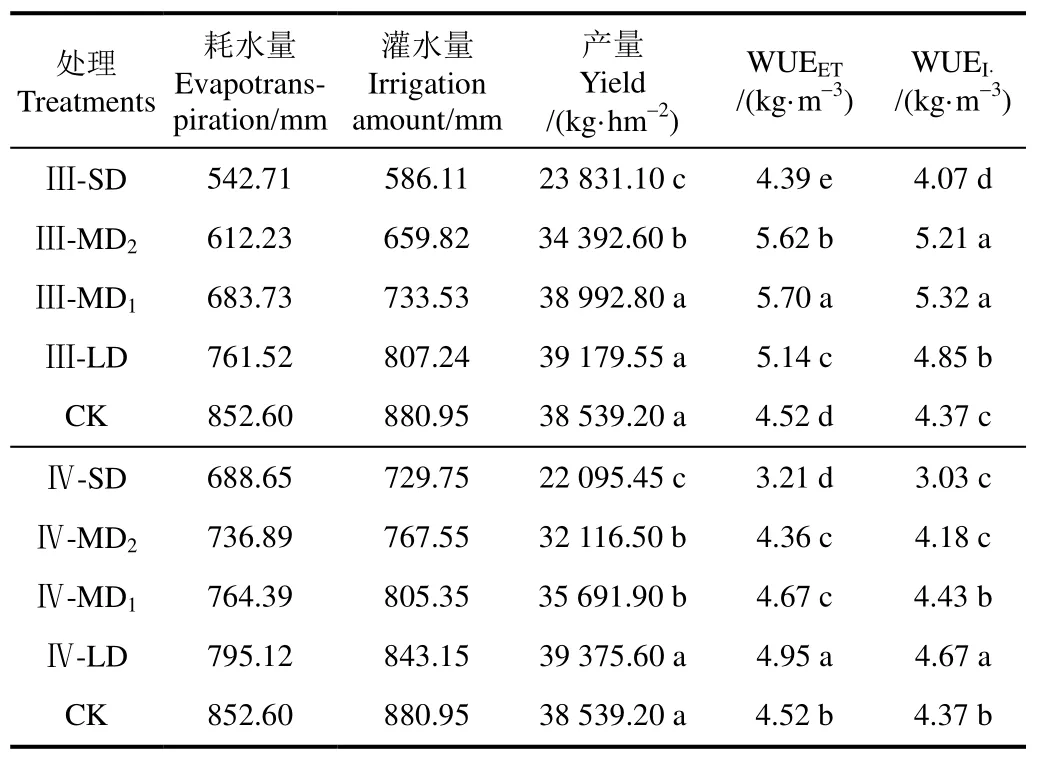
表3 水分亏缺对柑橘产量、耗水量及水分利用效率的影响Table 3 Effect of water deficit on fruit yield, evapotranspiration and water use efficiency of citrus
3 讨 论
本研究发现滴灌条件下在柑橘Ⅲ期和Ⅳ期进行轻度亏水处理均有利于提高叶片WUEi。与CK相比,Ⅳ-LD处理WUEi显著提高(P<0.05)。研究表明,作物遭受重度亏水后,叶片出现叶绿体膨胀、排列紊乱,基质片层模糊,基粒间连接松驰,类囊体片层肿胀或解体,光合器官的超微结构遭到破坏[23],导致从质体醌的电子转移池到 PSI终端受体的电子转移受损、气孔关闭、最大羧化速率减慢、RuBP酶的再生能力减弱,电子传递速率的减小也使卡尔文循环中ATP或NADPH的供应量减少,进而又影响 RuBP酶的再生能力[19],因此重度水分亏缺会显著降低叶片 Pn,甚至破坏光合系统,不利于有机物的形成。然而,Cui等[24]在冬小麦拔节期和分蘖期进行轻度亏水处理后(叶水势为(-1.40 MPa,-1.20 MPa),土壤含水率为田间持水量的60%~65%)发现,叶片光系统II的最大光化学效率、实际光化学效率、最大羧化速率、光合电子传递速率均增加,非光化学猝灭降低,Pn均表现为增大。Zhang等[25-27]发现,胡瓜轻度亏水初期光合关键酶 rbcl和 rbcs的相关 mRNA表达呈上升趋势,且Rubisco酶活性提高,其Pn的提高较CK差异不显著。WUEi的提高得益于Pn降幅小于Tr降幅,当土壤含水率越低,棵间蒸发强度就越低,土壤毛管传导度相应减小,从而降低了根系吸水速率,引起叶片含水率降低、保卫细胞因失水而收缩、叶片气孔开度减小以及气孔水分扩散阻力增加,最终导致叶面蒸腾强度降低[28]。叶片蒸腾强度的显著降低致使WUEi得到提高。Xia等[29]也得到了相似结论,研究发现怪柳在亏水处理后(相对土壤含水量为60%~75%)Pn、WUEi比对照(相对土壤含水量为85%~93%)分别提高 49%、41.94%。Roccuzzo等[13]也发现柑橘各生育期适度亏水使WUEi提高13%~15%。
本研究发现滴灌水分亏缺后柑橘叶片Pn的主要影响因素由气孔限制向非气孔限制的转变与亏水度和生育期均密切相关。Ⅲ期亏水度由MD2加剧为SD、Ⅳ期亏水度由MD1加剧为MD2时,影响柑橘光合作用的主要因素由气孔因素转为非气孔因素。研究表明,亏水初期气孔阻力增大,叶片水分散失减小,但 CO2供应的减少又使细胞内 O2成为电子传递的受体使活性氧自由基的产生加剧,因此,光合的气孔限制阻碍了水分胁迫的加剧,但同时也诱发了由自由基引发的非气孔限制[23]。显然,Ⅲ期气孔到非气孔的土壤含水率变化阈值低于Ⅳ期,说明Ⅳ期更容易由气孔限制转化为非气孔限制,可能由于Ⅲ期果实细胞分裂加速,细胞膨大,果实体积的增加对水分需求迫切,柑橘叶片在Ⅲ期对水分胁迫环境做出相应的适应。
本研究发现滴灌条件下在柑橘果实膨大期和果实成熟期进行轻度水分亏缺,对柑橘产量不会产生不利影响,并显著提高WUEET和WUEI。诸多研究得到了相似的结论,如Panigrahi等[10]发现,(相比对照)75%灌水量、75%施肥量的灌溉模式能使柑橘获得最大产量(16.39 t/ha)、最大水分利用效率(3.9 kg/m3)和最大肥料利用效率(87.3 kg/kg);Gasque等[12]在滴灌条件下对Navelina橘7月中旬—9月上旬进行亏水处理,经过连续5a的重复试验发现,在亏水期结束恢复全额灌溉时,果实出现明显加速生长的现象,最终柑橘增产4.5%~68%,节水12%~27%,并指出当Ψst>-2.0 MPa时亏水将对产量、品质带来不利影响;武阳等[18]发现,在果实缓慢膨大期中度水分亏缺后,香梨产量提高14.0%~18.0%,节水11.3%~13.2%;Tejero等[14]发现节水超过1 000 m3/hm2(约3 m3/棵)时,亏水处理会降低柑橘产量、影响果实品质。
4 结 论
1)在柑橘果实膨大期(Ⅲ)、果实成熟期(Ⅳ)亏水处理后,气孔导度(gs)、光合速率(Pn)、蒸腾速率(Tr)均随亏水度的加剧而降低,轻度亏水处理可以提高叶片瞬时水分利用效率(WUEi)。与 CK 相比,Ⅳ-LD 处理WUEi提高 36.61%(P<0.05)。
2)影响光合速率的主要因素由气孔限制与非气孔限制之间的转化与亏水度和生育期均密切相关,气孔限制值与非气孔限制值有互补的变化规律。处理间差异分析显示,随亏水度的加剧,影响光合作用的主要因素由气孔限制逐步转向非气孔限制,叶片气孔限制值与非气孔限制值分别增大1.24%~20.11%、2.17%~33.99%。Ⅲ期亏水度由MD2加剧为SD、Ⅳ期亏水度由MD1加剧为MD2时,影响柑橘光合作用的主要因素由气孔因素转为非气孔因素。
3)在柑橘Ⅲ期、Ⅳ期亏水处理后,各处理的耗水量随亏水度的加剧而减小,轻度亏水处理后产量较CK差异不显著。与CK相比,Ⅲ-LD和Ⅳ-LD处理的产量差异未达显著水平(P>0.05),WUEET显著提高 13%和 9.5%,WUEI分别显著提高 11%和 6.87%(P<0.05)。因此,柑橘Ⅲ、Ⅳ期适度的亏水处理可保证柑橘产量,节约灌溉用水,有效提高WUEET和WUEI,是柑橘适宜的滴灌水分亏缺模式。
[1] 韩启彪,冯绍元,曹林来,等. 滴灌技术与装备进一步发展的思考[J]. 排灌机械工程学报,2015,33(11):1001-1005.Han Qibiao, Feng Shaoyuan, Cao Linlai, et al. Thinking about further development of drip irrigation technology and equipment[J]. Journal of Drainage and Irrigation Machinery Engineering. 2015, 33(11): 1001-1005. (in Chinese with English abstract)
[2] 谭娟,郭晋川,吴建强,等. 不同灌溉方式下甘蔗光合特性[J]. 农业工程学报,2016,32(11):150-158.Tan Juan, Guo Jinchuan, Wu Jianqiang, et al. Photosynthetic characteristics of sugarcane underdifferent irrigation modes[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2016, 32(11): 150-158. (in Chinese with English abstract)
[3] Davies W J, Zhang J. Root signals and the regulation of growth and development of plants in drying soil [J]. Annual Review of Plant Physiology and Plant Molecular Biology,2003, 42(1): 55-76.
[4] Reynolds M P, Acevedo E, Sayre KD, Fischer RA. Yield potential in modem wheat varieties:its association with a less competitive ideotype[J]. Field Crops Research, 1994, 37(3):149-160.
[5] 郝树荣,郭相平,张展羽. 作物干旱胁迫及复水的补偿效应研究进展[J]. 水利水电科技进展,2009,29(1):81-84.Hao Shurong, Guo Xiangping, Zhang Zhanyu, et al. Research progress on compensatory effects of crops in drought stress and rehydration[J]. Advances in Science and Technology of Water Resources, 2009, 29(1): 81-84. (in Chinese with English abstract)
[6] Chaves M M, Pereira J S, Maroco J, et al. How plants cope with water stress in the field? Photosynthesis and Growth[J].Photosynthesis and growth. Annual Botany, 2002, 89(7):907-916.
[7] 康绍忠,杜太生,孙景生,等. 基于生命需水信息的作物高效节水调控理论与技术[J]. 水利学报,2007,38(6):661-667.Kang Shaozhong, Du Taisheng, Sun Jingsheng, et al. Theory and technology of improving irrigation water use efficiency based on crop growing water demand information[J]. Journal of Hydraulic Engineering, 2007, 38(6): 661-667. (in Chinese with English abstract)
[8] Hsiao T C. Plant responses to water stress[J]. Ann,Rev Plant Physoil, 1973, 24: 519-570.
[9] Thomas F R, Ronald L P, James A F. Drought response of young apple tree on three root stockexchange,chlorophy ll fluore scence,water relations and leaf abscisic acid[J] . Amer SocHort Sci, 1997, 122(6): 841-848.
[10] Panigrahi P, Srivastava A K. Effective management of irrigation water in citrus orchards under a water scarce hot sub—humid region[J]. Scientia Horticulturae, 2016, 210: 6-13.
[11] Chang Y C, Chang Y S, Lin L H. Response of shoot growth,photosynthetic capacity, flowering, and fruiting of potted‘Nagami’ kumquat to different regulated deficit irrigation[J].Horticulture, Environment and Biotechnology, 2015, 56(4):444-454.
[12] Gasque María, Martí Pau, Granero Beatriz, et al. Effects of long-term summer def i cit irrigation on ‘Navelina’ citrus trees[J]. Agricultural Water Management, 2016, 169: 140-147.
[13] Roccuzzo Giancarlo, Francisco J Villalobos, et al. Effects of water def i cits on whole tree water use eff i ciency of orange[J].Agricultural Water Management, 2014, 140: 61-68.
[14] Tejero I Garcı´a, Vicente R Romero, Jime´nez-Bocanegra J A,et al. Response of citrus trees to def i cit irrigation during different phenological periods in relation to yield, fruit quality, and water prod uctivity[J]. Agricultural Water Management. 2010, 97: 689-699.
[15] 江天然,张立新,毕玉蓉,等. 水分胁迫对梭梭叶片气体交换特征的影响[J]. 兰州大学学报,2001,37(6):57-62.Jiang Tianran, Zhang Lixin, Bi Yurong, et al. Effects of water stress on gas exchange characteristics of Haloxylon Ammodendron leafs[J]. Journal of Lanzhou University, 2001,37(6): 57-62. (in Chinese with English abstract)
[16] 许大全. 光合作用气孔限制分析中的一些问题[J]. 植物生理学通讯,1997,33(4):241-244.Xu Daquan. Some problems in stomatal limitation analysis of photosynthesis[J]. Plant Physiology Communications, 1997,33(4): 241-244. (in Chinese with English abstract)
[17] 徐俊增,彭世彰,魏征,等. 节水灌溉水稻叶片胞间CO2浓度及气孔与非气孔限制[J].农业工程学报. 2010,26(7):76-80.Xu Junzeng, Peng Shizhang, Wei Zheng, et al. Intercellular CO2concentration and stomatal or non—stomatal limitation of rice under water saving irrigation[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2010, 26(7): 76-80. (in Chinese with English abstract)
[18] 杨泽粟,张强,郝小翠. 自然条件下半干旱雨养春小麦生育后期旗叶光合的气孔和非气孔限制[J]. 中国生态农业学报,2015(2):174-182.Yang Zesu, Zhang Qiang, Hao Xiaocui. Stomatal or non-stomatal limitation of photosynthesis of spring wheat flag leaf at late growth stages under natural conditions in semiarid rainfed regions[J]. Chinese Journal of Eco—Agriculture, 2015(2): 174-182. (in Chinese with English abstract)
[19] 武阳,王伟,雷廷武,等. 调亏灌溉对滴灌成龄香梨果树生长及果实产量的影响[J]. 农业工程学报,2012,28(11):118-124.Wu Yang, Wang Wei, Lei Tingwu, et al. Impact of regulated deficit irrigation on growth and fruit yield of mature fragrant pear trees under trickle irrigation[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2012, 28(11): 118-124. (in Chinese with English abstract)
[20] Campos H, Trejo C, Peña—Valdivia CB, et al, Stomatal and non-stomatal limitations of bell pepper (Capsicum annuum L.)plants under water stress and re-watering: Delayed restoration of photosynthesis during recovery[J]. Environmental & Experimental Botany, 2014, 98 (1): 56-64.
[21] 杨楠. 滴灌设备技术及其应用现状[J]. 民营科技,2016(2):18-18.
[22] Crafts-Brandner SJ. Inhibition and acclimation photosynthesis to heat stress in closely correlated with activation of rubulose—1,5 bisphosphate earboxylase/oxygensase[J]. Plant Physiology, 1999, 120(1): 173-181.
[23] Liu F, Andersen MN, Jacobsen SE, et al. Stomatal control and water use eff i ciency of soybean (Glycine max L. Merr.)during progressive soil drying[J]. Environmental &Experimental Botany, 2005, 54 (1): 33-40.
[24] Cui Yakun, Tian Zhongwei, Zhang Xu, et al. Effect of water def i cit during vegetative growth periods on post-anthesis photosynthetic capacity and grain yield in winter wheat(Triticum aestivum L.) [J]. Acta Physiol Plant, 2015, 37(10):1-10.
[25] Zhang Lidong, Zhang Liuxia, Sun Jianlei, et al. Rubisco gene expression and photosynthetic characteristics of cucumber seedlings in response to water def i cit[J]. Scientia Horticulturae. 2013, 161: 81-87.
[26] de Souza TC, Magalha˜es PC, de Castro EM, et al. ABA application to maize hybrids contrasting for drought tolerance:changes in water parameters and in antioxidant enzyme activity[J]. Plant Growth Regulation, 2014, 73(3): 205-217.
[27] 裴冬,孙振山,陈四龙,等. 水分调亏对冬小麦生理生态的影响[J]. 农业工程学报,2006,22(8):68-72.Pei Dong, Sun Zhenshan, Chen Silong, et al. Effects of water stress on physiological and agronomic features of winter wheat[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2006, 22(8): 68-72. (in Chinese with English abstract)
[28] Doorenbos J, Pruitil W. Crop Water Requirement[M]. 2nded.Rome: FAO, 1977.
[29] Xia J, Zhang S, Guo J, et al. Critical effects of gas exchange parameters in Tamarix chinensis Lour on soil water and its relevant environmental factors on a shell ridge island in China’s Yellow River Delta[J]. Ecological Engineering, 2015,76: 36-46.
