缺氧条件下巨噬细胞移动抑制因子对肉鸡心肌脂肪酸代谢的影响
2018-01-26李丽芳王彦美裴芳樱李嘉威孙耀贵段智变李宏全王文魁
任 昊,李丽芳,王彦美,黄 楠,裴芳樱,李嘉威,孙耀贵,段智变,李宏全,王文魁*
(1.山西农业大学动物科技学院,太谷 030801; 2.山西农业大学信息学院环境科学与食品工程系,太谷 030801;3.滨州学院生物工程学院,滨州 256600)
肉鸡肺动脉高压综合征(pulmonary hypertension syndrome, PHS),又称腹水综合征(ascites syndrome, AS),作为严重影响肉鸡养殖业的重要代谢性疾病,一直以来备受养鸡从业者和科学研究者的高度重视。AS属代谢综合征,其发生发展涉及多因素、多层次、多环节,以往的研究主要集中于缺氧引起肺动脉高压这一中心环节[1-4],部分解释了本病的发生发展过程,尚不能完全揭示其发病机制。右心肥大是该综合征的显著特征[5-6],阐明右心肥大的发生发展,从另一视角审视肉鸡腹水综合征,无疑有利于全面揭示本病的发病机制。
心力衰竭及心脏肥大过程中腺苷酸活化蛋白激酶(adenosine monophosphate activated kinase, AMPK)激活,一方面,AMPK依赖性磷酸化6-磷酸果糖激酶被激活,促进糖酵解,生成更多ATP,满足心肌能量代谢需求[7-8];另一方面,AMPK通过激活脂肪酸转位酶(fatty acid translocase, FAT/CD36)进而促进心肌摄取游离脂肪酸(free fatty acid, FFA),并通过磷酸化乙酰辅酶A抑制其向丙二酰辅酶A(malonyl-CoA)转化,从而增强心肌对脂肪酸的利用[9-10]。笔者前期研究发现,AS患病肉鸡心脏巨噬细胞移动抑制因子(macrophage migration inhibitory, MIF)表达量显著高于健康对照鸡。哺乳动物研究资料提示,MIF在心肌缺血、缺氧等应激反应过程中通过激活AMPK参与心肌能量代谢调节,对心脏生理活动和病理变化具有重要作用[11-12]。为了确认MIF是否通过激活AMPK参与鸡心肌细胞脂肪酸代谢调节,我们开展了此项研究。
1 材料与方法
1.1 试验动物
9~12日龄AA肉鸡鸡胚,购自山西康牧种鸡场。
1.2 主要试剂
1.2.1 细胞培养用试剂 DMEM高糖培养基(SH30022.01,HyClone)、新生牛血清(16010-159,Gibco)、P/S双抗青链霉素(PAA)、0.25%胰蛋白酶(25200-056,Sigma)、胶原酶Ⅱ(C8150,Solarbio)。氯化钴(C8661,Sigma)、ISO-1(Ab142140,Abcam)。
1.2.2 免疫细胞化学试验用试剂 4%多聚甲醛溶液(Solarbio)、Triton X-100(Solarbio)、α-Sarcomeric actin(BM0001,Boster)、SABC-POD(小鼠IgG)试剂盒(SA1021,Boster)、羊抗鼠IgG-HRP(Boster)、苏木素伊红(HE)染色试剂盒(Solarbio)、DAB显色试剂盒(AR1022,Boster)
1.2.3 蛋白质免疫印迹用试剂 除兔抗鸡MIF多抗为实验室自制外,其他均为商品化多克隆抗体:HIF-1α(D262108,BBI)、AMPK(bs-1115R,Bioss)、p-AMPK(2531,Cell Signaling)、FAT/CD36(D161529,BBI)、ACC(D155300,BBI)、p-ACC(D155180,BBI)、CPT-1A(bs-2047R,Bioss)。羊抗兔IgG-HRP二抗(Bioss),RIPA裂解液(Beyotime),4×蛋白质上样缓冲液(Solarbio)。
1.3 方法
1.3.1 鸡胚心肌细胞分离培养 根据P. Simpson等的方法适当改进[13]。取孵化9~12日龄肉鸡蛋,用75%酒精消毒蛋壳后转入超净工作台,进行无菌操作:取出鸡胚,暴露胸腔,摘取心放入预冷的无钙、镁PBS缓冲液,吹打清洗,剔除心冠脂肪血管等组织; PBS缓冲液(无钙、镁)漂洗3次后剪成直径约1 mm大小的组织块,再用PBS缓冲液(无钙、镁)漂洗3次。加入0.08%胰蛋白酶溶液,37 ℃恒温振荡消化,大约5 min后弃上清液,再加0.125%胰蛋白酶与0.1% Ⅱ型胶原酶(等量混合),37 ℃静置消化8 min,加10%新生牛血清DMEM高糖液体培养液终止消化,如此重复消化数次,直至组织块呈絮状半透明样为止。经200目筛网过滤,去除细胞团块,常温1 000 r·min-1离心8 min,弃上清,用完全培养基重悬,移入30 mm细胞培养皿,常规(37 ℃、5% CO2)培养40 min除成纤维细胞,再以1×106·mL-1接种于6孔培养板常规培养,并分别于18、42 h更换培养液。
1.3.2 心肌细胞形态机能观察 细胞培养至24、48 h时,倒置显微镜下观察心肌细胞生长情况及自主搏动频率、强度,进行拍照、录像。
1.3.3 免疫细胞化学鉴定 采用免疫细胞化学染色鉴定心肌细胞。将培养48 h的细胞爬片用预冷PBS清洗3次,4%多聚甲醛室温固定细胞15 min,再用0.5% Triton X-100细胞通透剂处理30 min;接着按即用型SABC-POD试剂盒说明进行操作:10% H2O2阻断内源性过氧化物酶,5% BSA封闭,一抗(1∶300)4 ℃湿盒孵育过夜,二抗室温孵育1 h,DAB显色(黄)。稀释液PBS作阴性对照。
1.3.4 心肌细胞化学性缺氧模型的建立 24 h培养良好的心肌细胞,换无血清DMEM高糖培养基饥饿培养8~12 h,细胞呈现同步化搏动时开始建模。试验组给予不同浓度(400、500、600和700 μmol·L-1)氯化钴,正常对照组加入无血清培养基,37 ℃、5% CO2培养24 h,摸索化学性缺氧最佳条件。
1.3.5 MIF阻断试验 在缺氧的同时,试验组给予不同浓度(10、50和100 μmol·L-1)MIF阻断剂(ISO-1),正常对照组加入无血清培养基,37 ℃、5% CO2培养24 h,观察ISO-1对MIF的阻断效果。

2 结 果
2.1 原代鸡胚心肌细胞形态学及机能活动观察
刚分离消化的心肌细胞呈类圆形,悬浮于培养液中;培养24 h后细胞大分部贴壁,呈梭形、不规则形,立体感明显且具较强折光性(图1A),细胞搏动明显,但节律不一致,30~60次·min-1;48 h后心肌细胞伸出伪足相互接触交联成网状(图1B),收缩明显而有力,出现同步搏动,节律80~130次·min-1。
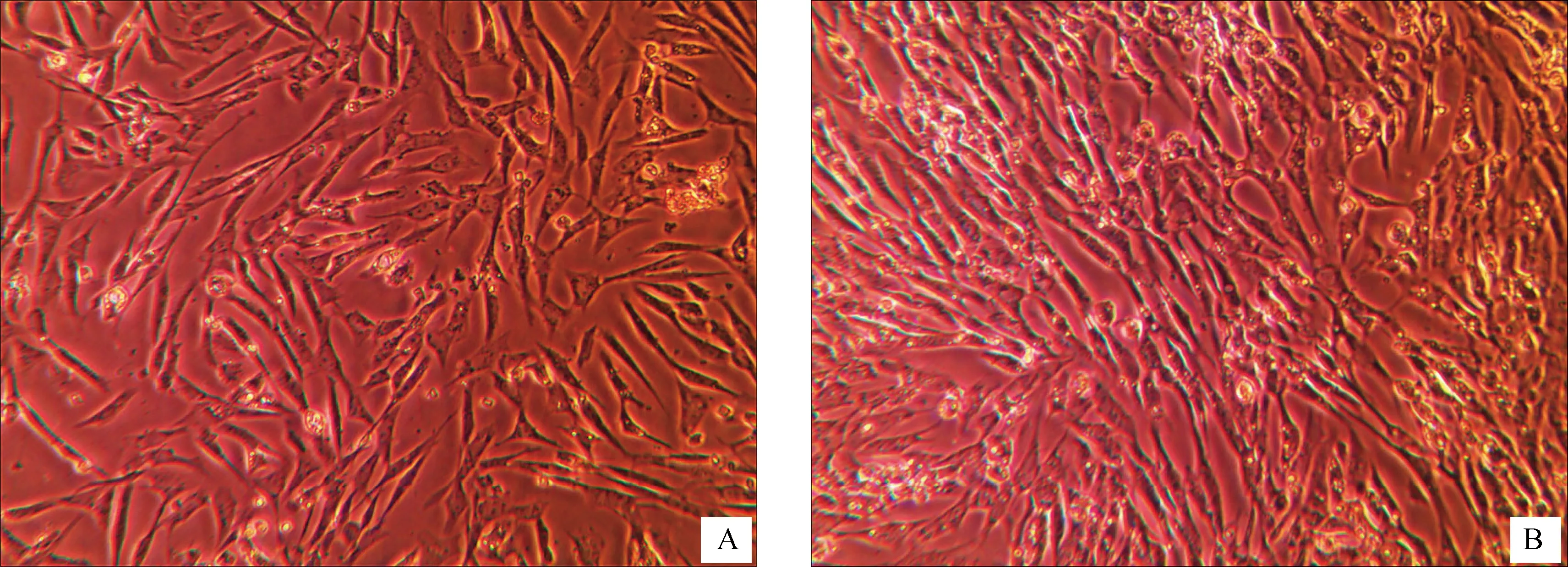
A.培养24 h;B.培养48 hA. Cardiomyocytes cultured for 24 hours;B. Cardiomyocytes cultured for 48 hours图1 光镜下原代鸡胚心肌细胞(200×)Fig.1 Primary chicken embryonic cardiomyocytes in vivo (200×)
2.2 免疫细胞化学鉴定
免疫细胞化学染色结果显示,阳性组心肌细胞核蓝染,细胞质充满细小均匀的棕黄色丝状结构,心肌细胞占90%以上(图2A);阴性对照组以PBS替代一抗,细胞质内未见棕黄色肌丝结构(图2B)。
2.3 心肌细胞缺氧模型的构建
Western blot检测结果显示(图3、表1),与正常组相比,600 μmol·L-1氯化钴处理组HIF-1ɑ和MIF蛋白表达量极显著升高(P<0.01)。
2.4 ISO-1阻断MIF试验
Western blot检测结果显示(图4、表2),与正常组相比,氯化钴处理组MIF及p-AMPK蛋白表达量均极显著升高(P<0.01);与氯化钴处理组相比,ISO-100 μmol·L-1阻断组MIF及p-AMPK蛋白表达量均极显著降低(P<0.01)。

A. 阳性染色组;B. 阴性对照组A. Positive staining group incubated with α-sarcomeric actin;B. Negative control with PBS图2 原代鸡胚心肌免疫细胞化学鉴定(400×)Fig.2 The chicken embryonic cardiomyocytes incubated with α-sarcomeric actin (400×)

0、400、500、600和700 μmol·L-1氯化钴处理鸡胚心肌细胞24 h HIF-1α及MIF的免疫印迹The chicken embryonic cardiomyocytes were treated with 0, 400, 500, 600 or 700 μmol·L-1 CoCl2 for 24 h, and the expression levels of HIF-1α and MIF were determined by Western blot图3 氯化钴处理的鸡胚心肌细胞HIF-1α、MIF表达变化Fig.3 Effects of cobalt chloride on the expression of HIF-1α, MIF in the chicken embryonic cardiomyocytes
表1经氯化钴处理鸡胚心肌细胞HIF-1α、MIF表达量
Table1ExpressionofHIF-1αandMIFinthecardiomyocytestreatedwithCoCl2

目的蛋白质Targetprotein对照组Controlgroup氯化钴组CoCl2group400μmol·L-1500μmol·L-1600μmol·L-1700μmol·L-1HIF-1α11.167±0.007*1.537±0.010**1.721±0.021**1.243±0.019*MIF11.235±0.0152.085±0.075**4.492±0.071**2.931±0.004**
与对照组相比,*.P<0.05;**.P<0.01
Compared with the control group,*.P<0.05;**.P<0.01

0、10、50和100 μmol·L-1 ISO-1处理鸡胚心肌细胞24 h,MIF及p-AMPK免疫印迹The chicken embryonic cardiomyocytes were treated with 0, 10, 50 and 100 μmol·L-1 ISO-1 for 24 h, and the expression levels of MIF and p-AMPK were determined by Western blot图4 ISO-1处理鸡胚心肌细胞24 h后其MIF、p-AMPK表达变化Fig.4 Effects of ISO-1 on the expression of MIF and p-AMPK in the chicken embryonic cardiomyocytes
表2ISO-1处理鸡胚心肌细胞MIF、p-AMPK表达量
Table2ExpressionofMIFandp-AMPKinthecardiomyocytestreatedwithISO-1

目的蛋白质Targetprotein对照组Controlgroup氯化钴组CoCl2groupCoCl2+ISO组CoCl2+ISOgroupCoCl2+10μmol·L-1ISOCoCl2+50μmol·L-1ISOCoCl2+100μmol·L-1ISOMIF13.835±0.105**3.232±0.089#2.985±0.053##2.880±0.080##p-AMPK11.668±0.087**1.414±0.0041.208±0.055#1.083±0.083##
与对照组相比,*.P<0.05;**.P<0.01。与氯化钴组相比,#.P<0.05;##.P<0.01
Compared with the control group,*.P<0.05;**.P<0.01. Compared with the CoCl2group,#.P<0.05;##.P<0.01
2.5 缺氧状态下鸡胚心肌细胞脂肪酸代谢途径关键蛋白质的变化
HIF-1α、MIF、FAT/CD36和CPT-1A均以β-actin为内参,p-AMPK以AMPK和β-actin为内参,p-ACC以总ACC和β-actin为内参。Western blot检测结果显示(图5、表3),与对照组相比,缺氧组鸡胚心肌细胞HIF-1α、MIF、FAT/CD36、CPT-1A及磷酸化p-AMPK和p-ACC表达量极显著增多(P<0.01)。
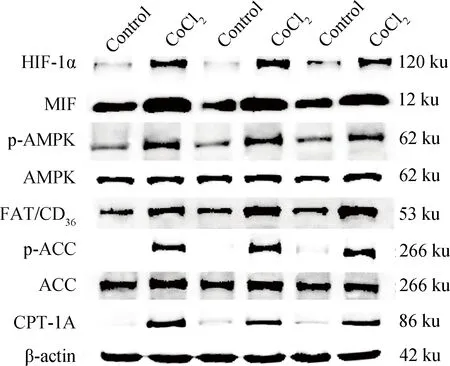
图5 缺氧状态下鸡胚心肌细胞HIF-1α、MIF、p-AMPK、FAT/CD36、p-ACC、CPT-1A表达的免疫印迹分析Fig.5 The Western blot analysis of HIF-1α、MIF、p-AMPK、FAT/CD36、p-ACC and CPT-1A in chicken embryonic cardiomyocytes in hypoxia
表3缺氧状态下鸡胚心肌细胞脂肪酸代谢途径关键蛋白质的变化
Table3Theexpressionlevelsofthekeyproteinsoffattyacidmetabolicpathwayinchickenembryoniccardiomyocytesinhypoxia

目的蛋白质Targetprotein对照组Controlgroup氯化钴组CoCl2groupHIF-1α0.993±0.0341.176±0.021**MIF1.000±0.0471.731±0.167**p-AMPK1.053±0.0491.753±0.104**FAT/CD361.057±0.0531.905±0.052**p-ACC1.140±0.1443.901±0.163**CPT-1A1.074±0.1222.844±0.253**
与对照组相比,*.P<0.05;**.P<0.01
Compared with the control group,*.P<0.05;**.P<0.01
2.6 阻断MIF后各目的蛋白质在鸡胚心肌中的表达变化
Western blot检测结果显示(图6、表4),与缺氧组相比,ISO-1阻断组鸡胚心肌细胞MIF、FAT/CD36、CPT-1A及磷酸化p-AMPK和p-ACC表达量极显著降低(P<0.01)。

图6 ISO-1处理后缺氧鸡胚心肌细胞MIF、p-AMPK、FAT/CD36、p-ACC、CPT-1A表达的免疫印迹分析Fig.6 The Western blot analysis of MIF、p-AMPK、FAT/CD36、p-ACC and CPT-1A in chicken embryonic cardiomyocytes treated with ISO-1 in hypoxia
3 讨 论
肉鸡腹水综合征这一代谢性疾病严重影响着养鸡业的经济效益和鸡肉品质,目前认为缺氧是导致肉鸡发生AS的主要原因[14-15]。肉鸡生长快,心肺功能相对较弱,这是造成机体相对缺氧或绝对缺氧的先天因素;饲养管理不当、环境气候变化、营养失调、罹患其他疾病等造成机体处于应激状态,加剧缺氧,引起肺动脉重构,肺循环阻力增大,使处于“能量饥饿”的心脏,特别是右心,代偿性肥大[16-17]。
为了探讨缺氧对心肌能量代谢的影响,我们首先建立了肉鸡鸡胚心肌细胞缺氧模型。获得高纯度、高活力的鸡胚心肌细胞是本研究的基础。结果
表4ISO-1处理后缺氧鸡胚心肌细胞脂肪酸代谢途径关键蛋白质的变化
Table4TheexpressionlevelsofthekeyproteinsoffattyacidmetabolicpathwayinchickenembryoniccardiomyocytestreatedwithISO-1inhypoxia

目的蛋白质Targetprotein对照组Controlgroup氯化钴组CoCl2group抑制剂组ISOgroupMIF1.005±0.0053.995±0.222**2.847±0.295##p-AMPK0.995±0.0052.361±0.124**1.446±0.034##FAT/CD360.830±0.1701.676±0.040**1.039±0.050##p-ACC0.885±0.1152.359±0.038**1.087±0.189##CPT-1A0.975±0.0253.760±0.090**2.605±0.045##
与对照组相比,*.P<0.05;**.P<0.01。与氯化钴组相比,#.P<0.05;##.P<0.01
Compared with the control group,*.P<0.05;**.P<0.01. Compared with the CoCl2group,#.P<0.05;##.P<0.01
显示(图1、2),免疫细胞化学鉴定及心肌特有功能(自律性搏动)均确认,采用含10%新生牛血清的高糖DMEM培养液培养鸡胚心肌细胞获得成功,其搏动频率在80~130次·min-1。
氯化钴作为一种化学缺氧模拟剂,其二价钴通过竞争二价铁抑制脯氨酸羟化酶的活性,提高HIF-1α的稳定性,促进HIF-1的转录活性,使常氧条件下低氧反应元件调控的基因得以转录表达[18-20]。笔者的研究结果显示(图3),400~700 μmol·L-1CoCl2作用24 h时HIF-1α表达显著增加,且二者在400~600 μmol·L-1CoCl2间呈正相关,CoCl2浓度再增大(700 μmol·L-1)时,HIF-1α表达量反下降。提示600 μmol·L-1CoCl2处理心肌细胞24 h,可获得良好的体外培养细胞缺氧模型。
S. M. Welford等研究发现,HIF-1α能与MIF启动子区域特异性低氧反应原件(HRE)结合,进而调控MIF的转录表达[21]。图3表明,CoCl2浓度在400~700 μmol·L-1范围内,鸡胚心肌细胞HIF-1α和MIF的表达具有相同变化趋势,在400~600 μmol·L-1间二者表达量逐渐增大,增至700 μmol·L-1时二者表达量均下降。这种高度拟合的变化趋势充分佐证,鸡MIF基因同样具有低氧反应元件,其转录表达受HIF-1α调控。
AMPK作为细胞的“能量感受器”,在细胞能量代谢中扮演着“总开关”的作用[22-24]。心脏缺血再灌注损伤时引起心肌自分泌MIF,通过激活AMPK,改善其能量代谢[25-26]。我们前期研究发现[27],AS患病肉鸡心脏组织MIF、p-AMPK表达量均显著高于正常肉鸡,猜测缺氧时MIF、p-AMPK与腹水的发生有关。试验结果显示(图4),缺氧时心肌MIF表达增多、AMPK活化增强;ISO处理缺氧心肌细胞后,MIF表达减少、AMPK活化降低。这种现象提示:缺氧时鸡心肌细胞自分泌MIF增多,后者通过激活AMPK调节心肌能量代谢。心肌主要依赖脂肪酸供能(占心肌总耗能的60%~90%)[28],我们利用Western bolt检测了缺氧鸡胚心肌细胞脂肪酸代谢途径关键蛋白的变化(图5、6),缺氧时FAT/CD36、磷酸化乙酰辅酶A羧化酶(acetyl CoA carboxylase, p-ACC)和肉毒碱棕榈酰转移酶1(carnitine palmitoyltransferase 1, CPT-1A)的蛋白质表达增加;ISO阻断MIF后这三种蛋白表达均显著降低。由此推断,活化的AMPK一方面激活FAT/CD36,促进脂肪酸跨细胞膜向细胞内转运;另一方面磷酸化ACC使其失活,从而阻止丙二酰辅酶A形成,解除丙二酰辅酶A对CPT-1A的抑制,加速脂酰辅酶A(fatty acyl-CoA)进入线粒体氧化供能。
综上,缺氧可以刺激心肌自分泌MIF,MIF经跨膜信号转导引起AMPK磷酸化,活化的AMPK促进脂肪酸代谢,加强氧化供能,为缺氧提供代偿性保护。我们的研究只静态地观测了一个时间点,对心肌缺氧后能量代谢进行了初步框架性研究,其动态过程还有待今后深入探究。
4 结 论
采用体外心肌细胞缺氧模型及MIF阻断法证明,缺氧可以刺激心肌自分泌MIF,MIF经跨膜信号转导引起AMPK磷酸化,活化的AMPK促进脂肪酸代谢,为缺氧心肌提供代偿性能量供应。
[1] SHEN Y R, SHI S R, TONG H B, et al. Metabolomics analysis reveals that bile acids and phospholipids contribute to variable responses to low-temperature-induced ascites syndrome[J].MolBiosyst, 2014, 10(6): 1557-1567.
[2] ZHANG J J, FENG X J, ZHAO L H, et al. Expression of hypoxia-inducible factor 1α mRNA in hearts and lungs of broiler chickens with ascites syndrome induced by excess salt in drinking water[J].PoultSci, 2013, 92(8): 2044-2052.
[3] YANG F, CAO H B, XIAO Q Y, et al. Transcriptome analysis and gene identification in the pulmonary artery of broilers with ascites syndrome[J].PLoSOne, 2016, 11(6): e0156045.
[4] LI H Y, WANG Y M, CHEN L L, et al. The role of MIF, cyclinD1 and ERK in the development of pulmonary hypertension in broilers[J].AvianPathol, 2017, 46(2): 202-208.
[5] TAN X, HU S H, WANG X L. Possible role of nitric oxide in the pathogenesis of pulmonary hypertension in broilers: a synopsis[J].AvianPathol, 2007, 36(4): 261-267.
[6] LI K, ZHAO L H, GENG G R, et al. Increased calcium deposits and decreased Ca2+-ATPase in erythrocytes of ascitic broiler chickens[J].ResVetSci, 2011, 90(3): 468-473.
[7] WALLHAUS T R, TAYLOR M, DEGRADO T R, et al. Myocardial free fatty acid and glucose use after carvedilol treatment in patients with congestive heart failure[J].Circulation, 2001, 103(20): 2441-2446.
[8] RAMANNA H, ELVAN A, WITTKAMPF F H, et al. Increased dispersion and shortened refractoriness caused by verapamil in chronic atrial fibrillation[J].JAmCollCardiol, 2001, 37(5): 1403-1407.
[9] NEUBAUER S. The failing heart—an engine out of fuel[J].NEnglJMed, 2007, 356(11): 1140-1151.
[10] STANLEY W C, RECCHIA F A, LOPASCHUK G D. Myocardial substrate metabolism in the normal and failing heart[J].PhysiolRev, 2005, 85(3): 1093-1129.
[11] MA H, WANG J J, THOMAS D P, et al. Impaired macrophage migration inhibitory factor (MIF)-AMPK activation and ischemic recovery in the senescent heart[J].Circulation, 2010, 122(3): 282-292.
[12] MILLER E J, LI J, LENG L, et al. Macrophage migration inhibitory factor stimulates AMP-activated protein kinase in the ischaemic heart[J].Nature, 2008, 451(7178): 578-582.
[13] SIMPSON P, SAVION S. Differentiation of rat myocytes in single cell cultures with and without proliferating nonmyocardial cells. Cross-striations, ultrastructure, and chronotropic response to isoproterenol[J].CircRes, 1982, 50(1): 101-116.
[14] WIDEMAN R F, RHOADS D D, ERF G F, et al. Pulmonary arterial hypertension (ascites syndrome) in broilers: a review[J].PoultSci, 2013, 92(1): 64-83.
[15] OLKOWSKI A A. Pathophysiology of heart failure in broiler chickens: structural, biochemical, and molecular characteristics[J].PoultSci, 2007, 86(5): 999-1005.
[16] HASSANZADEH M, BUYSE J, TOLOEI T, et al. Ascites syndrome in broiler chickens: a review on the aspect of endogenous and exogenous factors interactions[J].JPoultSci, 2014, 51(3): 229-241.
[17] BAGHBANZADEH A, DECUYPERE E. Ascites syndrome in broilers: physiological and nutritional perspectives[J].AvianPathol, 2008, 37(2): 117-126.
[18] MAXWELL P H, WIESENER M S, CHANG G W, et al. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis[J].Nature, 1999, 399(6733): 271-275.
[19] JAAKKOLA P, MOLE D R, TIAN Y M, et al. Targeting of HIF-α to the von hippel-lindau ubiquitylation complex by O2-regulated prolyl hydroxylation[J].Science, 2001, 292(5516): 468-472.
[20] KALLIO P J, WILSON W J, O′BRIEN S, et al. Regulation of the hypoxia-inducible transcription factor 1α by the ubiquitin-proteasome pathway[J].JBiolChem, 1999, 274(10): 6519-6525.
[21] WELFORD S M, BEDOGNI B, GRADIN K, et al. HIF1α delays premature senescence through the activation of MIF[J].GenesDev, 2006, 20(24): 3366-3371.
[22] RUSSELL III R R, LI J, COVEN D L, et al. AMP-activated protein kinase mediates ischemic glucose uptake and prevents postischemic cardiac dysfunction, apoptosis, and injury[J].JClinInvest, 2004, 114(4): 495-503.
[23] OAKHILL J S, STEEL R, CHEN Z P, et al. AMPK is a direct adenylate charge-regulated protein kinase[J].Science, 2011, 332(6036): 1433-1435.
[24] ADACHI Y, KANBAYASHI Y, HARATA I, et al. Petasin activates AMP-activated protein kinase and modulates glucose metabolism[J].JNatProd, 2014, 77(6): 1262-1269.
[25] WANG J Y, TONG C, YAN X Y, et al. Limiting cardiac ischemic injury by pharmacological augmentation of macrophage migration inhibitory factor-AMP-activated protein kinase signal transduction[J].Circulation, 2013, 128(3): 225-236.
[26] MU J, BROZINICK J T Jr, VALLADARES O, et al. A role for AMP-activated protein kinase in contraction-and hypoxia-regulated glucose transport in skeletal muscle[J].MolCell, 2001, 7(5): 1085-1094.
[27] 韩丽娟. 肉鸡腹水综合征(AS)中MIF与AMPK表达关系的研究[D]. 太谷: 山西农业大学, 2015.
HAN L J. The relationship between MIF and AMPK expression in ascites syndrome (AS)[D]. Taigu: Shanxi Agricultural University, 2015. (in Chinese)
[28] PIRRO M, MAURIGE P, TCHERNOF A, et al. Plasma free fatty acid levels and the risk of ischemic heart disease in men: prospective results from the Québec cardiovascular study[J].Atherosclerosis, 2002, 160(2): 377-384.
