海南岛异尾轮属物种多样性研究
2018-01-24林秋奇吴秋婷韩博平
林秋奇 吴秋婷 韩博平
(暨南大学水生生物研究所, 广州 510632)
轮虫是淡水后生动物中种类多样性比较高的一个类群, 广泛分布于湖泊、水库和河流以及水稻田和池塘等浅水水体或临时性水体中。轮虫通常是淡水水体, 尤其是富营养化水体中重要的初级消费者。水体中轮虫的种类组成不仅与水体的营养状态有关, 而且还取决于其所处的动物地理分区。但是, 对于轮虫而言, 不同的属由于起源于地不同,它们的扩散与地理分布模式可以完全不同[1,2]。如:种类多样性比较高的臂尾轮属起源于冈瓦纳古陆,并通过非洲和印度次大陆向欧亚大陆和北美洲扩散。因此, 臂尾轮属在澳大利亚和南美洲分布着较多的特有种, 在欧洲的特有种则比较少[1]。异尾轮属起源于劳亚古陆, 是轮虫种类多样性比较高的另一个属, 特有种基本上分布在北半球的古北界和新北界, 而南半球没有特有种分布, 呈现由北向南扩散的模式[2]。在目前已确定的67种异尾轮虫中, 9种新北界特有种, 5种古北界特有种, 4种全北界特有种[2]。
海南岛是我国最大的热带岛屿, 原是中国大陆向南延伸的一部分, 但自更新世中期开始, 由于频繁的火山运动与雷州半岛断陷, 之后与大陆多次分分合合, 直到第四纪冰期结束, 才形成一个独立的岛屿[3]。在热带地区, 臂尾轮属、腔轮属和异尾轮属是轮虫种类多样性比较高的三个属。黄丰明等[4]和林秋奇等[5]分别研究了海南岛腔轮属和臂尾轮属的种类组成与多样性, 发现海南岛虽然与华南大陆存在物种交流, 但由于存在气候差异, 海南岛腔轮虫和臂尾轮虫多样性均远高于我国大陆各地区, 而且这种差异性随着纬度的增加而增加。海南岛腔轮虫和臂尾轮虫种类主要由广布种、热带种、泛热带种类和东洋界特有种组成[4,5]。不同于臂尾轮属, 异尾轮属起源于劳亚古陆, 并由北向南扩散, 这种情形是否意味着海南岛异尾轮虫多样性低于我国大陆其他地区?针对上述问题, 我们采集了海南岛上各种类型水体的异尾轮虫, 分析其种类组成和地理分布特征, 并沿着纬度梯度与我国大陆几个异尾轮虫种类资料相对比较详细的地区(广东、华中、华东、华北和东北)进行比较。
1 材料与方法
于2010年11月用35 μm孔径的浮游生物网或手抄网对海南岛的文昌、琼海、万宁、陵水、保亭、三亚、五指山、琼中、屯昌、澄迈和海口11个县市的水库、池塘、河流、溪流、水稻田以及水坑等水体采集了43个轮虫定性样品; 2011年5月对上述县市各类型水体采集了85个样点的轮虫定性样品; 2011年8月在海口、定安、万宁、保亭、三亚和儋州采集了6个轮虫定性样品。采样点的分布详见黄丰明等[4]。现场将轮虫样品用4%的福尔马林固定。在显微镜下参照Koste[6]和Shiel等[7]进行种类鉴定; 同时将各种类制成永久性装片, 在显微描图仪下按一定比例进行绘图。对需要根据咀嚼器进行鉴定的种类, 参考De Smet[8]的方法用浓度为8%—10%的次氯酸钠溶液消解轮虫个体以提取咀嚼器, 用蒸馏水将咀嚼器清洗干净后风干喷金。将喷金的咀嚼器置于Hitachi TM-3000台式扫描电子显微镜下观察拍照。海南岛1995年异尾轮虫资料来自Koste等[9]和诸葛燕[10], 广东省异尾轮虫资料来自林秋奇等[11]和张才学等[12], 湖北及其周边地区异尾轮虫资料来自诸葛燕[10], 上海及其周边地区的异尾轮虫资料来自王家楫[13]和金丽文[14], 北京及其周边地区异尾轮虫资料来自王家楫[13]和诸葛燕[10], 吉林及其周边地区异尾轮虫资料来自诸葛燕[10]。基于异尾轮虫的种类定性数据, 计算海南岛与其他5个地区间的Bray-Curtis距离。统计分析在R3.2.5中用vegan程序包进行。
2 结果
2.1 种类组成
本次调查在海南岛检出异尾轮虫17种(表 1)。其中, 中国新记录种2种: 无颌异尾轮虫(Trichocerca agnatha)和巴西异尾轮虫(T. braziliensis); 东洋界新记录种4种: 无颌异尾轮虫、臂棘异尾轮虫(T. pygocera)、钝齿异尾轮虫(T. obtusidens)和红异尾轮虫(T. rosea)。暗小异尾轮虫(T. pusilla)、对棘异尾轮虫(T. similis)、纤巧异尾轮虫(T. tenuior)和巴西异尾轮虫是常见种类; 无颌异尾轮虫、长刺异尾轮虫(T. longiseta)、瓷甲异尾轮虫(T. porcellus)和鼠异尾轮虫(T. rattus)则比较罕见。
2.2 新记录种描述
巴西异尾轮虫Trichocerca braziliensis (Murray, 1913)(图1)体矮胖, 背部具2个高而明显的龙骨状隆脊, 长度为体长的1/4—1/3; 身体最宽处位于隆脊末端处; 左趾长度长于体长, 基部呈“S”状弯曲; 右趾长度为左趾长度的1/3; 咀嚼器杖型, 左右不对称。砧基末端具托, 呈倒“T”形; 右砧枝具明显的双分叉小翼板, 左砧枝具一个刺状延伸。左槌柄末端呈倒“7”形, 右槌柄末端略弯呈“J”形。体长:120—150 μm, 体寛60—90 μm, 右趾长20—45 μm,左趾长145—150 μm, 咀嚼器长55—60 μm。
巴西异尾轮虫和鼠异尾轮虫体型相似, 容易混淆(图1a和图2a)。它们最主要的区别是被甲背面龙骨状隆脊数量不同, 分别为 2 片和 1 片。咀嚼器的形状也有所不同, 如左槌柄(图1b和图2b)。
分布: 暖水性种类。在海口、文昌、琼海、五指山和琼中的鱼塘及水稻田检出该种。水温27—34℃, pH6.7—7.1。
无颌异尾轮虫Trichocerca agnatha Wulfert,1939(图3)体较细长, 背部略凸; 腹部凸; 头部甲鞘上有纵长的褶痕, 将头前半部隔成12个差不多大小的褶片, 无棘刺; 头冠具一对盘顶触手和一根感觉触毛; 背触手略偏向体右侧。左侧触手位于体左侧中部; 右触手位于腹部末端; 左趾直, 为体长的1/2; 右趾短于左趾长度的1/5。咀嚼器杖型, 左右不对称, 左槌柄长且粗壮, 末端弯曲呈“J”形; 右槌柄较左槌柄短且细, 末端弯曲呈“J”形(图3b); 左砧枝具长而尖的翼板; 砧基末端扇状(图3c)。体长137 μm,体寛50 μm, 左趾长75 μm; 咀嚼器长37 μm。
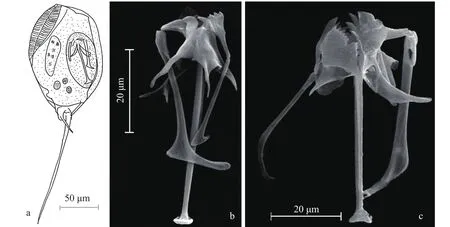
图1 巴西异尾轮虫Fig. 1 Trichocerca braziliensis
分布: 主要分布在古北界、新热带界和澳新界。为东洋界首次记录。在定安县和海口市的池塘和水稻田中检出该种。水温31—33℃; pH7.2—8.3。

图2 鼠异尾轮虫Fig. 2 Trichocerca rattus
2.3 地理区系与多样性特征
本次调查结合Koste等[9]和诸葛燕[10]的研究结果, 目前在海南岛共记录到异尾轮虫24种。种类组成包括广布性种类17种、冷水性种类3种[圆筒异尾轮虫(T. cylindrica)、鼠异尾轮虫和钝齿异尾轮虫]、暖水性种类2种[巴西异尾轮虫和卡顿异尾轮虫(T. chattoni)]、古北界特有种1种(臂棘异尾轮虫)和旧世界特有种1种(无颌异尾轮虫)。没有东洋界特有种发现。异尾轮虫在海南岛的种类多样性高于我国其他地区, 并且其种类多样性在我国呈现由低纬度向高纬度逐渐下降的趋势(图4)。此外,海南岛异尾轮虫种类组成与同处东洋界的广东、湖北(华中)和上海(华东)的Bray-Curtis相异性远小于位于古北界的北京(华北)和吉林(东北)(图5)。广东省与海南岛相隔一琼州海峡, 异尾轮虫种类组成Jaccard相似性指数为0.55; 高纬度的吉林省与海南岛相距3000 km以上, Jaccard相似性指数只有0.30。

图3 无颌异尾轮虫Fig. 3 Trichocerca agnatha
3 讨论
本调查与Koste等[9]和诸葛燕[10]对海南岛的调查记录相比, 增加了5个新记录种, 但有9种未在本次调查中发现(表 1)。Koste等[9]和诸葛燕[10]在1995年的调查在海口、琼海和三亚采集了40个样点; 本次采样点有134个, 遍布海南岛东部和中部地区, 以及西部的儋州; 生境包括水库、池塘、河流、小溪、水稻田和水坑等各种类型水体。2次调查结果基本上反映了海南岛异尾轮虫的种类多样性。我国动物地理分区属于世界动物地理分区的古北界和东洋界。Zhuge等[15]报道了中国共有异尾轮虫34种。本次调查增加了中国新记录种2种, 而金丽文[14]对上海的调查增加了中国新记录种1种,因此, 到目前为至, 中国异尾轮虫共有37种, 约占全球已确定异尾轮虫种类的55%。海南岛属于东洋界, 现有异尾轮虫24种, 约占我国异尾轮虫种类的65%。异尾轮虫起源于劳亚古陆, 仅在古北界和新北界有特有种。理论上讲, 温带地区异尾轮虫种类多样性高于热带地区。但是, 海南岛异尾轮虫多样性不仅远高于我国处于古北界的各地区, 也高于同处东洋界的广东[11]、湖南[10]和上海[14], 呈现高种类多样性的特征。
异尾轮虫地方特有种集中分布在北半球的古北界和新北界, 而热带地区没有特有种分布, 地理分布模式完全不同于臂尾轮虫和腔轮虫[2]。海南岛现有异尾轮虫记录24种, 其中种类组成包括广布型种类17种、冷水性种类3种、暖水性种类2种、古北界特有种1种和旧世界特有种1种。海南岛异尾轮虫种类分布模式与臂尾轮虫[5]和腔轮虫[4]也有所不同。臂尾轮虫和腔轮虫在海南岛上均有东洋界特有种的分布, 而且热带和泛热带种类所占的比例均超过50%; 异尾轮虫在海南岛上没有东洋界特有种的分布, 广布性种类占据了71%, 而暖水性种类只占8%。巴西异尾轮虫与鼠异尾轮虫、圆筒异尾轮虫与卡顿异尾轮虫被甲轮廓相似, 易混淆, 而且巴西异尾轮虫和卡顿异尾轮虫是暖水性种类, 而鼠异尾轮虫和圆筒异尾轮虫是冷水性种类, 一般不出现在热带和亚热带地区[2]。但是, 根据我们的调查结果以及其他文献资料, 上述冷水性种类, 也常常出现在东洋界的热带亚热带地区的轮虫记录中[10,11,16],这些冷水性种类可能正在向热带地区扩散, 并具有成为广布性种类的潜在可能性。另外, 被认为是古北界特有种的臂棘异尾轮虫和旧世界特有种的无颌异尾轮虫在海南岛的发现也表明异尾轮虫存在扩大分布区域的扩散行为。近几年来, 这种不同生物地理分区间的扩散行为也出现在其他的科和属中。如: 分布于美洲地区的美洲龟甲轮虫(Keratella americana)近几年陆续在武汉的东湖和广东、广西与云南的一些水库中出现[17,18]; 分布于美洲地区的哈瓦那臂尾轮虫(Brachionus havanaensis)在上海的一些水体中出现[14]。
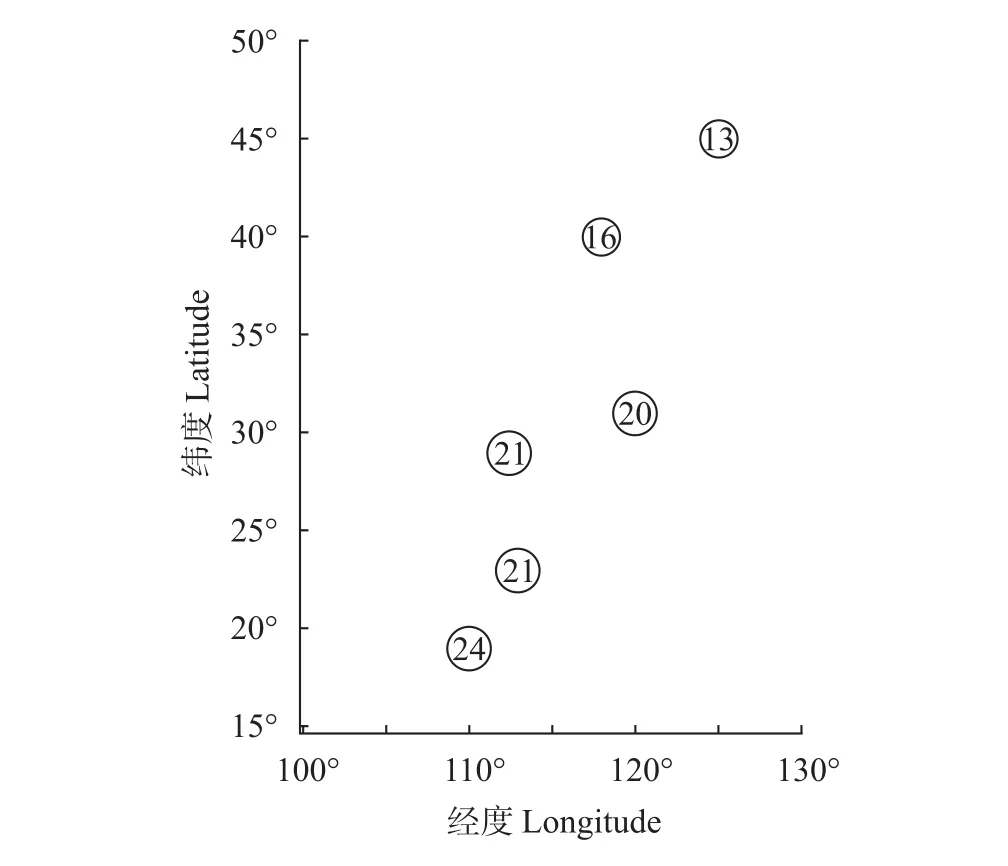
图4 异尾轮虫种类多样性沿着纬度和经度的梯度变化(圆圈中数字代表种类数量)Fig. 4 Species richness pattern of Trichocerca along the latitudinal and longitude gradients (The number in each circle indicates species richness)
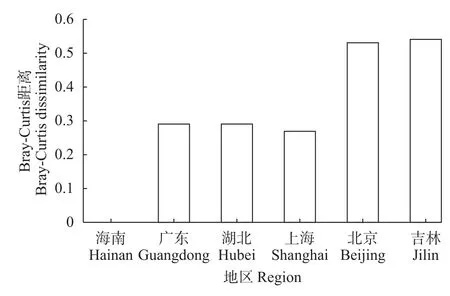
图5 海南岛与我国大陆其他五个地区异尾轮虫种类组成的Bray-Curtis距离Fig. 5 Bray-Curtis dissimilarity of Trichocerca between Hainan Island and the other five regions
海南岛原是中国大陆向南延伸的一部分, 自更新世中期, 由于频繁的火山运动与雷州半岛断陷,之后与大陆多次分分合合, 直到第四纪冰期结束,才形成一个独立的岛屿[3]。理论上讲, 地史上与大陆这种连、断、再连、再断的变化有助于海南岛与华南大陆之间的物种交流, 降低两地之间的种类组成差异性。古北界和旧世界特有种类在海南岛的出现说明海南岛与欧亚大陆存在物种的交流。虽然海南岛与我国大陆存在物种的交流, 但是, 根据林秋奇等对臂尾轮虫的研究结果看, 由于气候的差异, 臂尾轮虫种类组成仍然与我国大陆存在一定的差异, 而且差异度随着纬度的增加而增加[5]。对于异尾轮虫而言, 由于起源于劳亚古陆, 特有种集中分布在古北界和新北界, 所以分布在吉林或北京的冷水性种类[鼠异尾轮虫、圆筒异尾轮虫、马氏异尾轮虫(T. macera)、罗氏异尾轮虫(T. rousseleti)和钝齿异尾轮虫]、古北界特有种类[双褶异尾轮虫(T. vargai)和臂棘异尾轮虫]和全北界特有种类[奇异异尾轮虫(T. taurocephala)和钩状异尾轮虫(T.uncinata)]所占的比例相对比较高。相反, 海南岛异尾轮虫主要由广布性种类组成, 兼少数冷水性种类和暖水性种类。因此, 海南岛与处于古北界的吉林和北京异尾轮虫种类组成差异性比较大(Jaccard指数为0.30左右), 而与同处东洋界的广东、湖北和上海相对比较小(Jaccard指数为0.55左右)。总体上,海南岛异尾轮虫呈现高的种类多样性以及无特有种分布的特征。
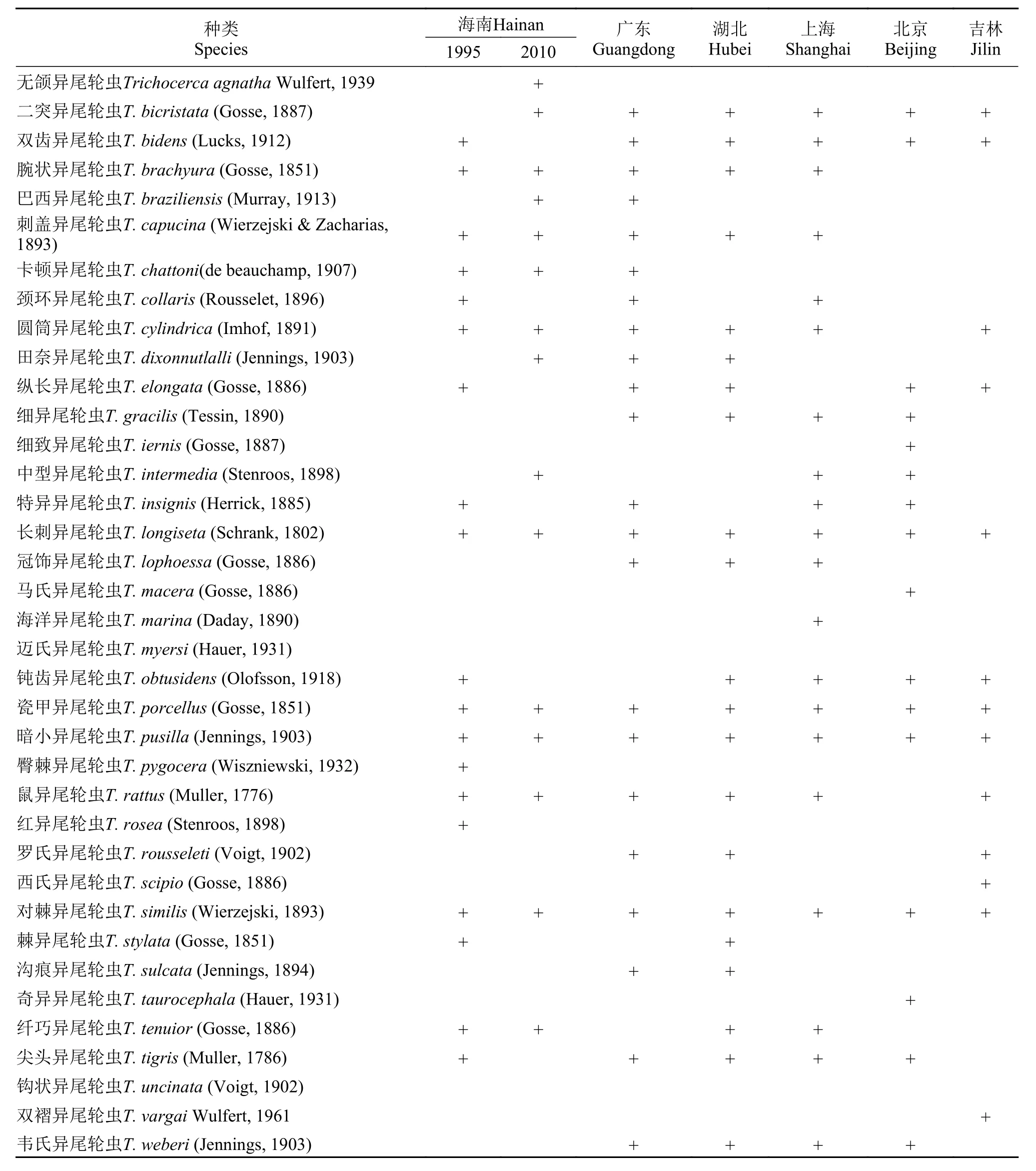
表 1 异尾轮虫种类组成Tab. 1 List of Rotifera of the genus Trichocerca from China
[1]Segers H. Annotated checklist of the rotifers (Phylum Rotifera), with notes on nomenclature, taxonomy and distribution [J].Zootaxa, 2007, 1564: 1—104
[2]Segers H. A biogeographical analysis of rotifers of the genusTrichocercaLamarck, 1801 (Trichocercidae,Monogononta, Rotifera), with notes on taxonomy [J].Hy-drobiologia, 2003, 500(1-3): 103—114
[3]Yan J A. Paleontology and ecologic environmental evolution of the Quaternary in Hainan Island [J].Journal of Palaeogeography, 2006, 8(1): 103—115 [颜家安. 海南岛第四纪古生物及生态环境演变. 古地理学报, 2006, 8(1):103—115]
[4]Huang F M, Wu Q T, Han B P,et al. Species diversity and distribution of Lecanidae in Hainan Island, China [J].Biodiversity Science, 2017, 25(4): 430—436 [黄丰明, 吴秋婷, 韩博平, 等. 海南岛腔轮科种类组成与区系分布.生物多样性, 2017, 25(4): 430—436]
[5]Lin Q Q, Wu Q T, Han B P. Species diversity ofBrachionusin Hainan Island, China [J].Chinese Journal of Zoology, 2017, 52(5): 783—791 [林秋奇, 吴秋婷, 韩博平.海南岛臂尾轮属物种多样性研究. 动物学杂志, 2017,52(5): 783—791]
[6]Koste, W. Rotatoria. Die Rädertiere Mitteleuropas [M].Borntraeger, Berlin. 1978, 131—142
[7]Shiel R J, Kost W. Rotifera from Australian inland waters VIII. Trichocerciade (Monogononta) [J].Transactions of the Royal Society of South Australia, 1992,116(1): 1—27
[8]De Smet W H. Preparation of rotifer trophi for light and scanning electron microscopy [J].Hydrobiologia, 1998,387/388: 117—121
[9]Koste W, Zhuge Y. A preliminary report on the occurrence of rotifer in Hainan [J].Quekett Journal of Microscopy, 1996, 37: 666—883
[10]Zhuge Y. Studies on taxonomy and distribution of Rotifera in typical zones of China [D].Thesis for Doctor of Science. Institute of Hydrobiology, the Chinese Academy of Sciences, Wuhan, 1997 [诸葛燕. 中国典型地带轮虫的研究. 博士学位论文, 中国科学院水生生物研究所,武汉, 1997]
[11]Lin Q Q, Zhao S Y, Han B P. Rotifer distribution in tropical reservoirs, Guangdong Province, China [J].Acta Ecologica Sinica, 2005, 25(5): 1123—1131 [林秋奇, 赵帅营,韩博平. 广东省水库轮虫分布特征. 生态学报, 2005,25(5): 1123—1131]
[12]Zhang C X, Zhan D L, Chen C L,et al. Analysis of plankton and pollution state in Xiaodongjiang River in Maoming Area [J].Journal of Guangdong Ocean University,2010, 30(4): 22—28 [张才学, 詹冬玲, 陈春亮, 等. 小东江茂名段浮游生物及污染状况分析. 广东海洋大学学报, 2010, 30(4): 22—28]
[13]Wang J J. Fauna of freshwater Rotifera of China [M].Beijing: Science Press. 1961, 208—228 [王家楫. 中国淡水轮虫志. 北京: 科学出版社. 1961, 208—228]
[14]Jin L W. The study of Rotifera in Shanghai and surrounding areas [D]. Thesis for Master of Science. Shanghai Normal University, Shanghai, 2013 [金丽文.上海及周边地区的轮虫研究. 硕士学位论文, 上海师范大学, 上海.2013]
[15]Zhuge Y, Huang X F, Koste W. Rotifera recorded from China, 1893-1997, with remarks on their composition and distribution [J].International Review of Hydrobiology,1998, 83(3): 217—232
[16]Wu Q T, Wei Z H, Zhu J,et al. Seasonal dynamics of rotifer in a seasonal fishing moratorium - Lake Erhai, Yunnan Province, China [J].Journal of Lake Sciences, 2012,24(4): 586—592 [吴秋婷, 卫志宏, 朱江, 等. 一座季节性休渔湖泊—洱海轮虫的季节动态特征. 湖泊科学, 2012,24(4): 586—592]
[17]Yan Q Y, Yu Y H, Feng W S,et al. Genetic diversity of plankton community as depicted by PCR-DGGE fingerprinting and its relation to morphological composition and environmental factors in Lake Donghu [J].Microbial Ecology, 2007, 54(2): 290—297
[18]Yu J Y, Wen Z M, Han B P,et al. Morphological comparison onKeratella americanaCarlin, 1943 andKeratellacommon species in tropical China [J].Acta Hydrobiologica Sinica, 2017, 41(2): 448—453 [喻婧怡, 温展明,韩博平, 等. 美洲龟甲轮虫的分类特征及与我国热带地区常见龟甲轮虫的比较. 水生生物学报, 2017, 41(2):448—453]
