大鳍异鰔年龄与生长
2018-01-24黄自豪危起伟王志坚
黄 静 黄自豪 危起伟 王志坚
(1. 西南大学生命科学学院, 淡水鱼类资源与生殖发育教育部重点实验室, 水产科学重庆市重点实验室, 重庆 400715;2. 中国水产科学研究院长江水产研究所 农业部淡水生物多样性保护重点实验室, 武汉 430223)
大鳍异鲦Creteuchiloglanis macropterus隶属于硬骨鱼纲Osteichthyes、鲇形目Siluriformes、鲦科Sisoridae、异鲦属Creteuchiloglanis, 分布于云南的怒江和伊洛瓦底江水系[1], 俗名石扁头、石拓子、老虎鱼等。目前, 对大鳍异鲦的研究主要集中于分类方面, Ng[2]提出, 分布于怒江和伊洛瓦底江的扁头鲦是误定种, 并将其定为新种——大鳍鲦, 孔德平[3]和姚景龙等[4]对其物种有效性做了探讨。2011年, Zhou等[1]提出将大鳍鲦划归异鲦属, 更名为大鳍异鲦。
大鳍异鲦为冷水激流性鱼类, 主要生活在河流上游人为活动较少的区域。随着人口的增加, 人类活动面积扩大, 其生活环境逐渐被破坏。大鳍异鲦肉质鲜嫩, 市场需求量大, 价格昂贵, 在高利益的驱使下, 捕捞强度很大。生存环境的破坏和高强度的捕捞, 使得大鳍异鲦野生资源在急速下降, 对其开展保护工作已迫在眉睫。本文以胸鳍棘作为大鳍异鲦年龄鉴定材料, 通过组织学切片观察其年龄组成。根据胸鳍棘切片半径与全长的关系, 得到退算全长, 通过研究其生长特征, 为今后的人工驯化和繁殖提供理论基础。
1 材料与方法
1.1 样本采集
2013年11月至2014年9月, 在怒江、槟榔江和龙川江共采集大鳍异鲦230尾(图1)。新鲜状态下测量全长和体长(精确到1 mm), 电子天平称重(精确到0.1 g), 常规解剖, 剪取左侧胸鳍编号保存于5%福尔马林溶液中。随机选取30尾标本, 取颅脑后正常形态的脊椎骨第2—5枚, 清洗、编号。
1.2 胸鳍棘年龄鉴定
取胸鳍棘于5%硝酸中脱钙1至数天, 以镊子使其弯曲变软为限, 用剪刀除去关节突起, 剪取基部适合的一段(1 cm左右), 流水冲洗24h。置于75%酒精中30min, 常规组织脱水、包埋、切片, 染色, 切片厚度为10—15 μm, Nikon80i显微镜观察拍照。
在未知体重、全长的情况下, 对胸鳍棘切片进行年轮观察判断。用胸鳍棘切片做年龄鉴定和生长退算的材料, 胸鳍棘半径的测量方法是以胸鳍棘中心为起点, 沿最长径突方向测量胸鳍棘半径, 至每一轮明带外缘处(图2)。
1.3 脊椎骨年龄鉴定
取脊椎骨放入4%NaOH溶液中煮沸3—5min,自来水中用细刷清洗, 在解剖镜下, 用解剖针剔除附着的肌肉, 自然干燥, 乙醚浸泡1—2d, 干燥。将椎体放入装有二甲苯的小培养皿中, 在体视镜下观察拍照。年轮的确认以反射光下暗带向明带转变处为标准, 在未知体重、全长的情况下, 对脊椎骨进行年轮观察判断。
1.4 年龄组的划分
年龄组划分为: 0龄组, 脊椎骨或胸鳍棘尚无年轮形成; Ⅰ龄组, 刚形成第1个年轮和轮纹外已有第2年生长环片, 但尚未形成第2个年轮的个体, 即Ⅰ和Ⅰ+; Ⅱ龄组, 刚形成第2个年轮和轮纹外已有第3年生长环片, 但尚未形成第3个年轮的个体, 即Ⅱ和Ⅱ+, 以此类推进行划分。
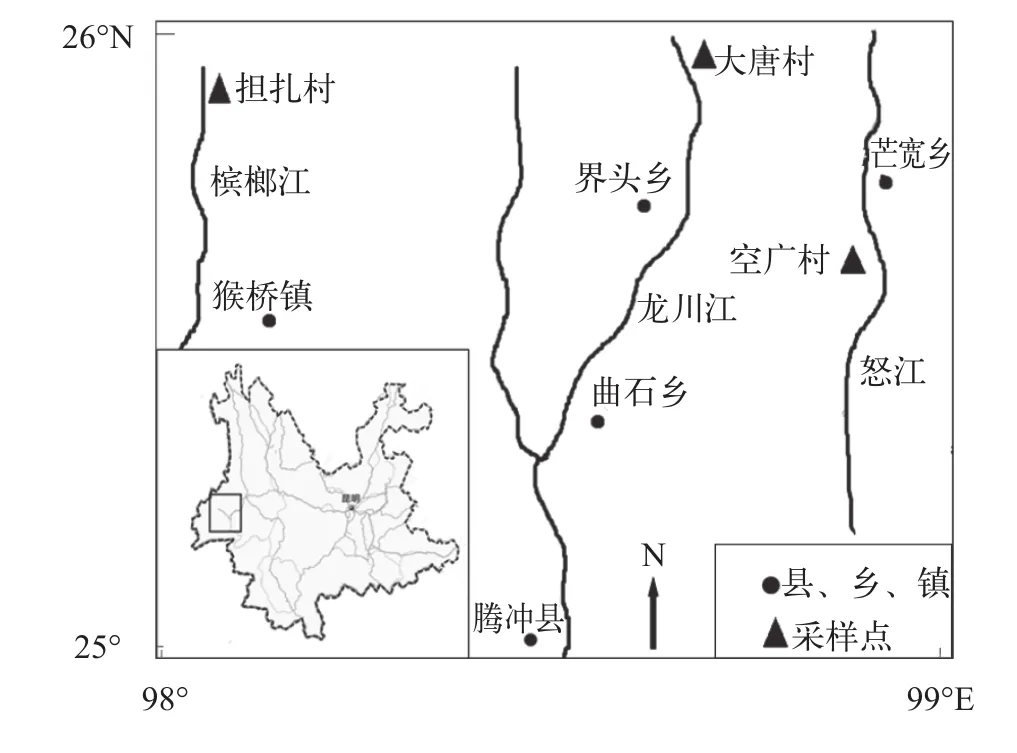
图1 大鳍异鲦采样点分布图Fig. 1 Geographical location of sampling sites for C. macropterus
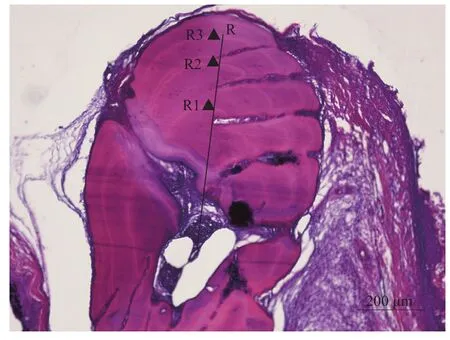
图2 大鳍异鲦胸鳍棘切片半径测量Fig. 2 The measurement of pectoral fin radius of C. macropterus
1.5 数据分析
丰满度采用K=W/L3·100。
相对生长率g=(l2-l1)/l1(t2-t1)。
生长比速Cv=(lgl2-lgl1)/0.4343·(t2-t1)。
生长常数Cvt=Cv(t2+t1)/2。
生长指标Clt=Cvl1。
对胸鳍棘半径与全长进行回归拟合, 选取相关系数最大的方程式。
全长与体重的关系用幂函数W=aLb来描述。
采用von Bertalanffy生长方程来描述大鳍异鲦的生长特性, 公式为:Lt:t时的全长,l1和l2为对应相邻年龄t1、t2的全长;Wt:t时的体重;L∞: 渐进全长;W∞: 渐进体重;t0: 假设理论生长起点年龄;k: 生长系数。
数据的分析与处理采用Excel 2007、Origin 9.1和SPSS 17.0软件。
2 结果
2.1 年龄鉴定
年轮特征胸鳍棘切片为不规则椭圆形, 年轮存在单环型和窄带型2种: 单环型, 年轮由一条环纹和宽带组成, 在HE染色下, 环纹着色浅, 透光度强, 易分辨; 窄带型, 年轮由一条窄带和一条宽带构成, 窄带由环纹及夹在中间的部分构成, 中间部分着色与宽带相同。随着年龄的增加, 胸鳍棘上的年轮排列逐渐变密(图3)。脊椎骨为双凹形, 反射光下, 椎体上存在窄带(暗带)和宽带(亮带), 二者相互交替排列, 与椎体成同心圆, 一个窄带和一个宽带构成一个年轮(图4)。
判别结果胸鳍棘切片230份, 年轮清晰可辨的有180份, 辨别率为78.26%。制得脊椎骨30份,年轮可辨的材料22份, 辨别率73.33%, 其中有19份与胸鳍棘切片相吻合, 吻合率为86.36%。
2.2 渔获物全长、体重和年龄组成
对180尾大鳍异鲦进行统计(图5—7), 雌性108尾, 雄性72尾, 雌雄性比为1.5鲶1。群体全长范围为60—182 mm, 主要分布在110 mm以下, 占67.78%,其次为110—160 mm, 占29.44%, 160 mm以上的极少; 体重范围为2.5—66.0 g, 主要分布在20 g以下,占86.11%, 其次为20—35 mm, 占9.45%, 35 g以上占4.44%; 年龄范围为1—9龄, 主要分布在1—3龄, 占65.00%, 其次为4—6龄, 占26.67%, 7—9龄仅占8.33%。
雌性群体全长范围为60—182 mm, 主要分布在120 mm以下, 占73.15%, 120 mm以上占26.85%;体重范围为2.5—66.0 g, 主要分布在20 g以下, 占82.41%, 20 g以上占17.59%; 年龄范围为1—9龄, 主要分布在1—4龄, 占73.15%, 4—9龄占26.85%。雄性群体全长范围为62—153 mm, 主要分布在110 mm以下, 占75.00%, 110以上占25.00%; 体重范围为2.5—34.9 g, 主要分布在20 g以下, 占91.67%; 年龄范围为1—7龄, 主要分布在1—3龄, 占75.00%。
2.3 胸鳍棘半径与全长的关系及生长退算
分别将雌雄样本的全长和胸鳍棘半径回归, 选择相关系数最大的拟合方程作为退算方程(图8):
雌性群体:L=2.0193(R2=0.8861,n=108)
雄性群体:L=2.0603(R2=0.8897,n=72)
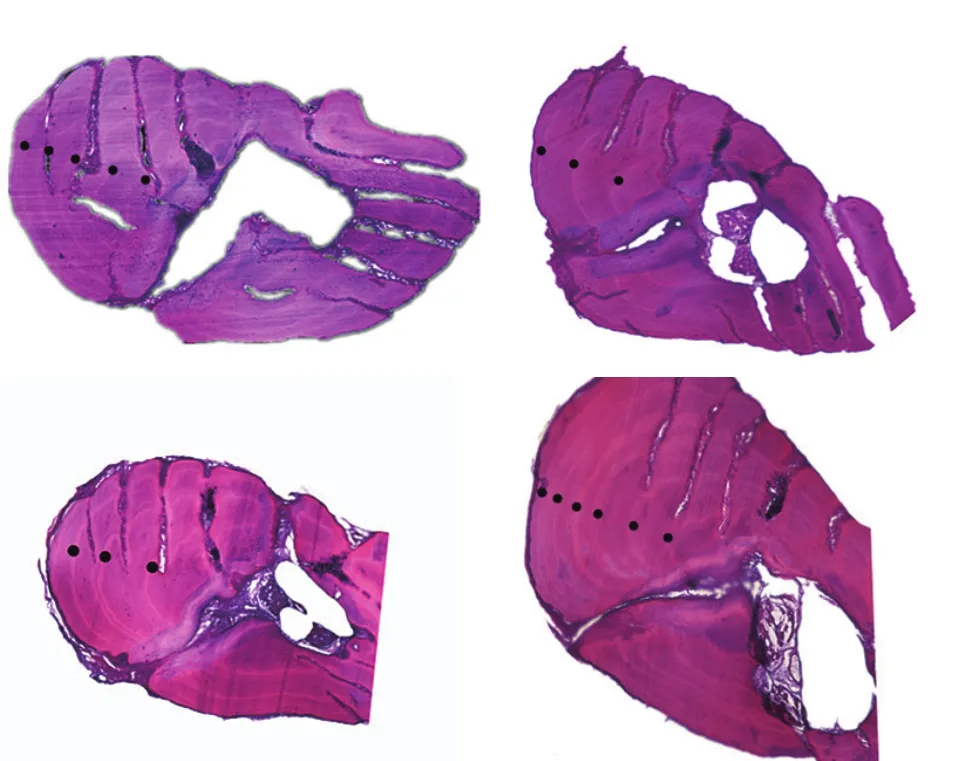
图3 大鳍异鲦胸鳍棘年轮特征Fig. 3 Annuli characters of pectoral fin for C. macropterus
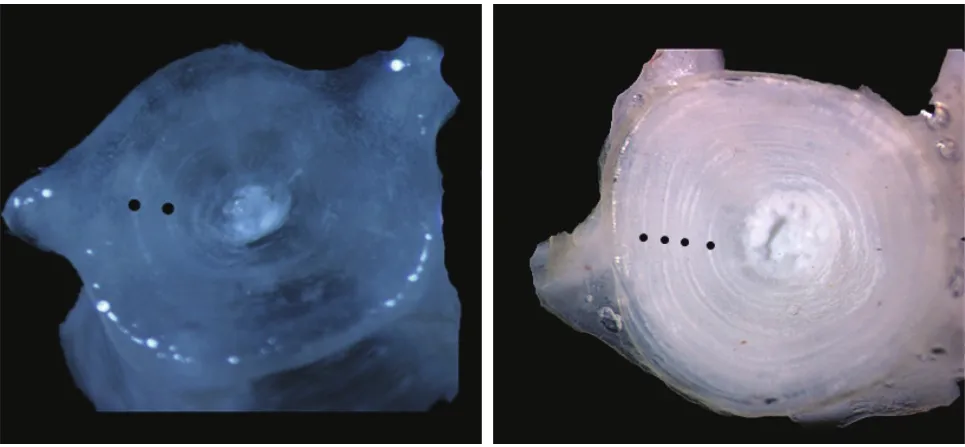
图4 大鳍异鲦脊椎骨年轮特征Fig. 4 Annuli character of vertebrae for C. macropterus
根据退算方程, 分别退算出大鳍异鲦雌雄各龄的全长(表 1、表 2)。通过χ2检验, 退算值与实测值间无显著差异: 雌:χ2=0.997,df=8,P=0.998; 雄:χ2=0.329,df=6,P=0.999。
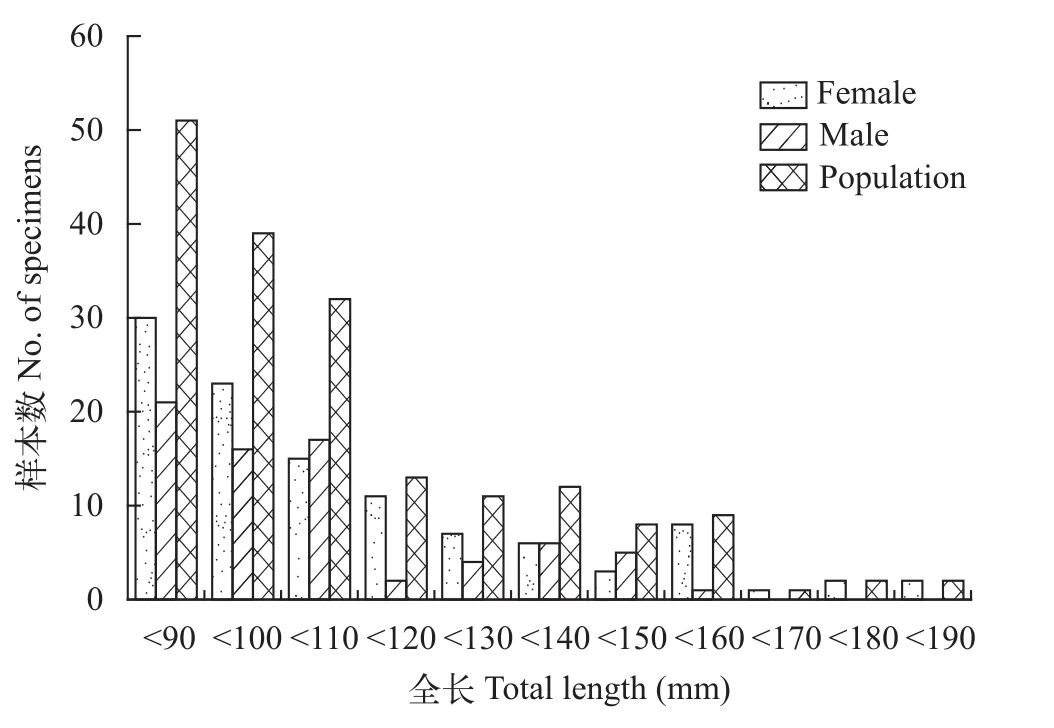
图5 大鳍异鲦渔获物全长分布Fig. 5 The distribution of total length of C. macropterus
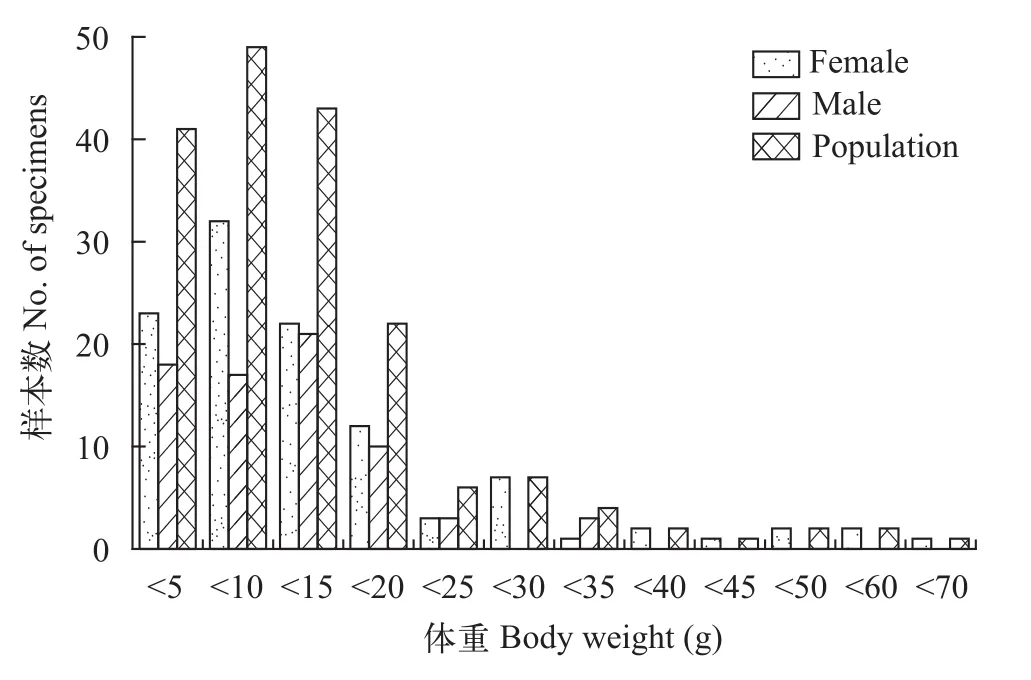
图6 大鳍异鲦渔获物体重分布Fig. 6 The distribution of Body length of C. macropterus
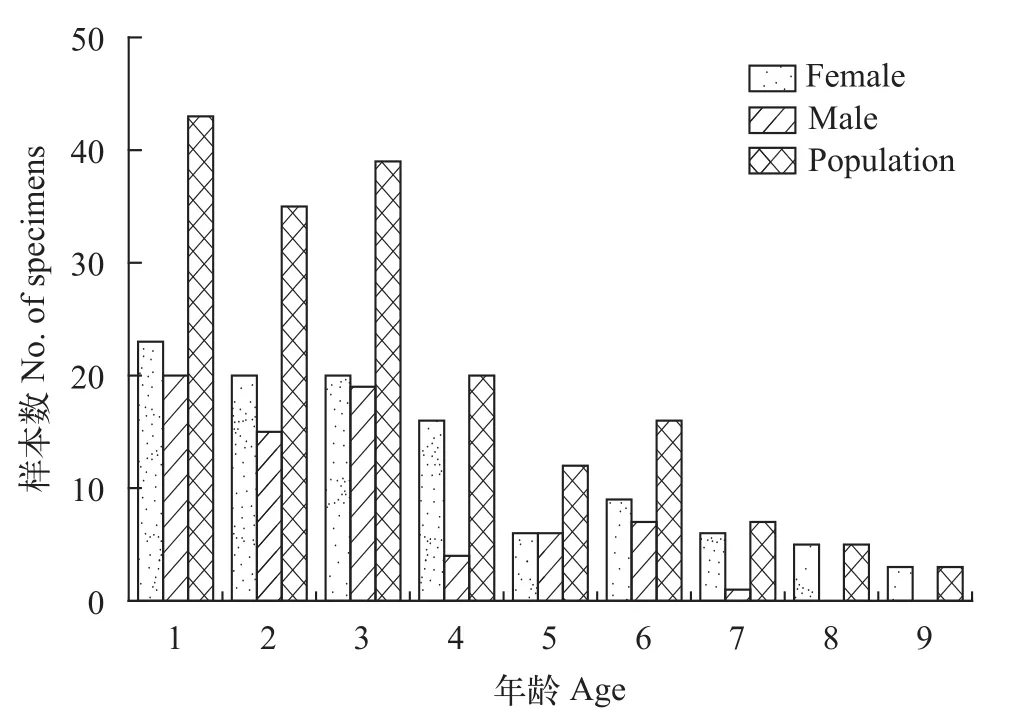
图7 大鳍异鲦渔获物年龄分布Fig. 7 The distribution of age of C. macropterus
将雌雄各龄退算全长进行t检验, 结果显示, 除2龄外, 其他各龄均无显著差异, 说明大鳍异鲦雌雄个体生长不存在显著差异, 因此可将其统一进行分析(表 3)。
把雌雄群体统一, 对全长和胸鳍棘半径做回归,选择相关系数最大的拟合方程做为退算方程(图9):
群体:L=2.0787(R2=0.8866,n=180)
根据退算方程, 退算出大鳍异鲦各龄全长(表 4),退算值与实测值χ2检验结果无显著差异:χ2=0.776,df=8,P=0.999。
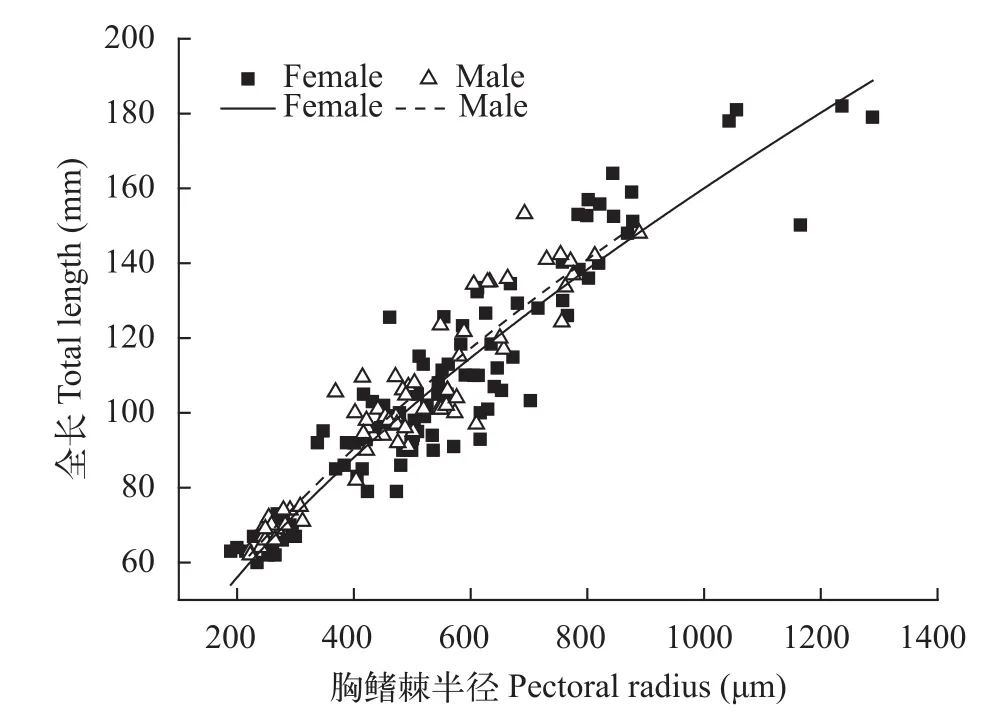
图8 大鳍异鲦雌雄全长与胸鳍棘半径关系Fig. 8 The relationship between total length and pectoral fin radius of female and male for C. macropterus
2.4 全长与体重关系
协方差分析表明大鳍异鲦雌雄实测全长与体重之间无明显差异(ANCOVA,P=0.554>0.05), 因此做统一分析。拟合得体重与全长的关系式为:W=1E-4L2.4807(R2=0.8189,n=180)(图10)。大鳍异鲦的b值为2.4807, 小于3, 属异速生长。
2.5 生长模型
根据Ford方程和von Bertalanffy方程, 将各龄退算全长带入, 求得各参数值:L∞=377.015 mm;k=0.0491;t0=-4.0195;W∞=246.134 g。由此得出大鳍异鲦的生长方程和生长曲线(图11):

表 1 大鳍异鰔雌鱼实测全长与退算全长Tab. 1 Measured and back-calculated total length of female C. macropterus
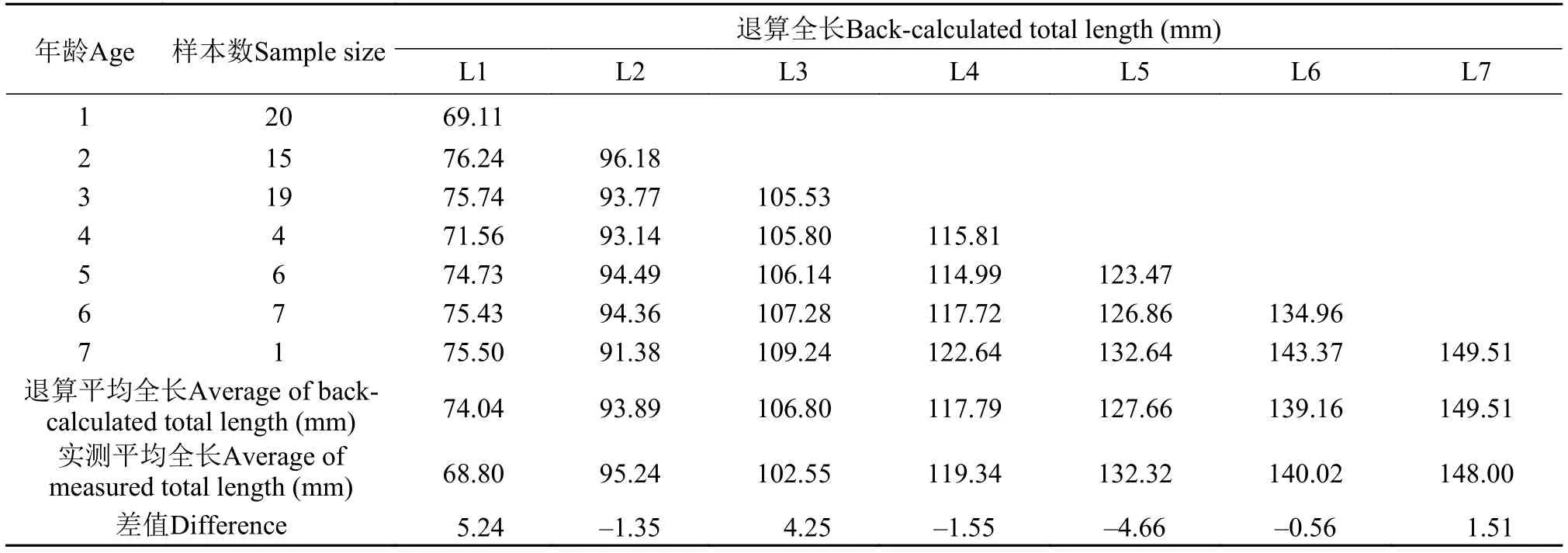
表 2 大鳍异鰔雄鱼实测全长与退算全长Tab. 2 Measured and back-calculated total length of male C. macropterus
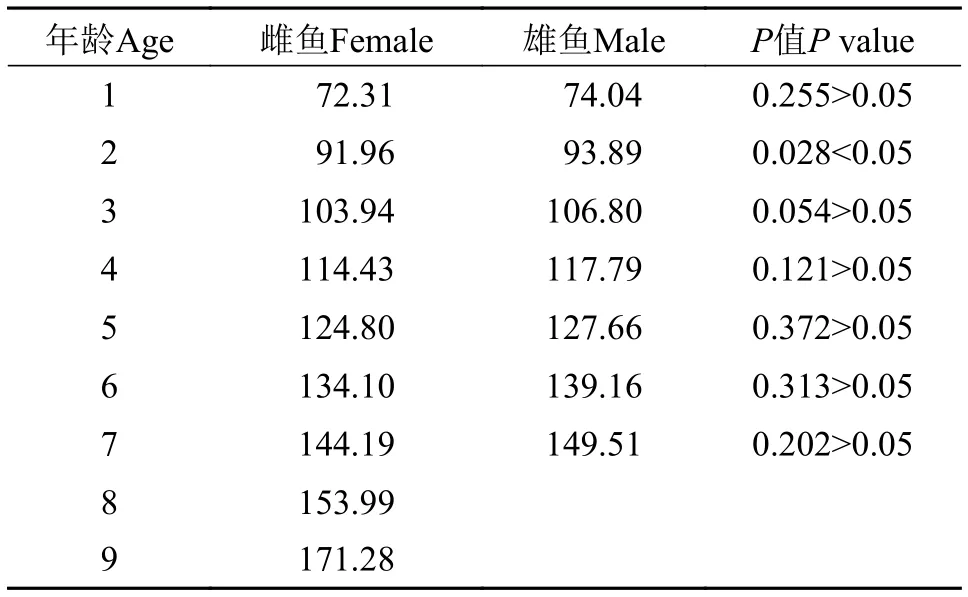
表 3 大鳍异鰔雌雄各龄退算全长比较Tab. 3 Comparison of back-calculated total length between female and male C. macropterus
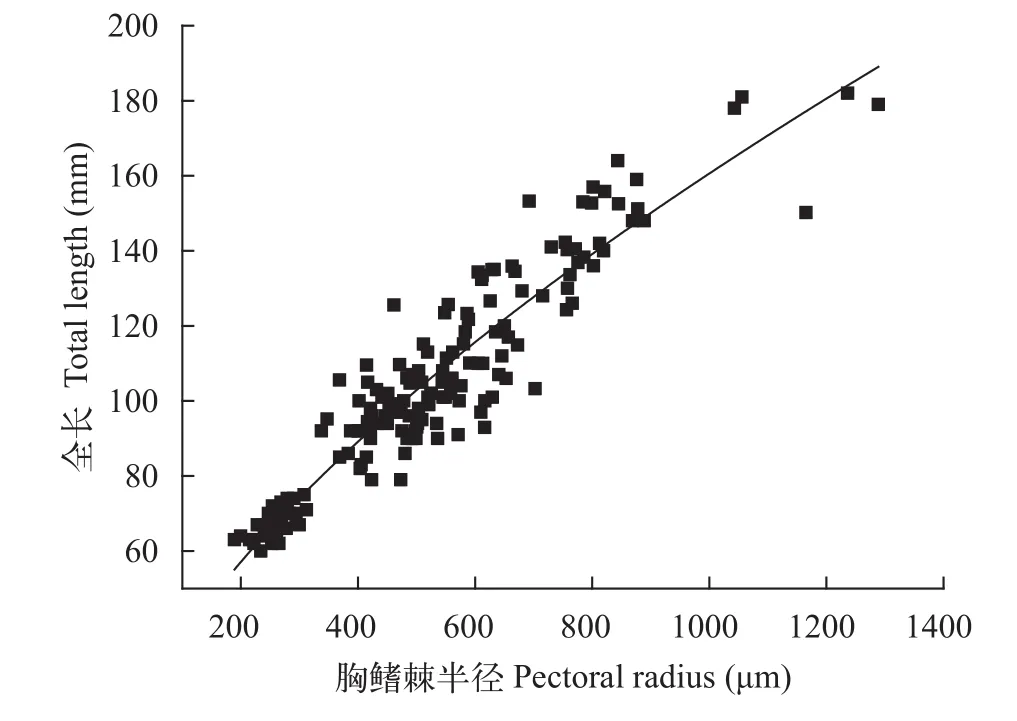
图9 大鳍异鲦全长与胸鳍棘半径关系Fig. 9 The relationship between total length and pectoral fin radius of C. macropterus
全长生长方程:Lt=377.015·[1-e-0.0491(t+4.0195)]
体重生长方程:Wt=246.134·[1-e-0.0491(t+4.0195)]2.4807
2.6 生长速度和生长加速度
将大鳍异鲦生长方程一阶、二阶求导, 分别得到全长和体重的生长速度和生长加速度:

从图12可以看出, 大鳍异鲦的全长生长不具生长拐点, 随着年龄增长, 全长生长速度逐渐减小, 并趋向于0, 全长生长加速度逐渐增大, 亦无限接近于0, 全长生长速度和生长加速度随着年龄增长变化趋势逐步减缓。
大鳍异鲦的体重生长速度先上升再下降(图13), 具有明显的生长拐点, 生长拐点处的体重生长加速度为零(d2W/dt2=0), 生长速度最大。大鳍异鲦的生长拐点年龄为14.48龄, 拐点处的全长和体重分别为225.00 mm和68.40 g, 14.48龄以后, 大鳍异鲦体重生长速度逐步下降, 生长加速度小于0。约30龄时, 体重生长加速度降至最低点, 之后逐步上升, 并趋近于0, 体重生长速度下降趋势减缓。
2.7 生长指数
根据生长指数经验公式, 结合退算全长, 求得大鳍异鲦的生长指数。从表 5可以看出: 大鳍异鲦的生长比速、生长指标、相对生长率随着年龄增加而逐渐减小; 生长常数可划分为2个阶段, 1至4龄属于未性成熟阶段, 生长常数均值为0.332, 5龄达到性成熟, 5至8龄为性成熟阶段, 生长常数均值为0.430。
2.8 肥满度
如表 6所示, 大鳍异鲦的肥满度随着年龄的增大逐步下降, 5龄以后基本稳定。5龄为大鳍异鲦的初次性成熟年龄, 说明大鳍异鲦性成熟后其肥满度基本固定。
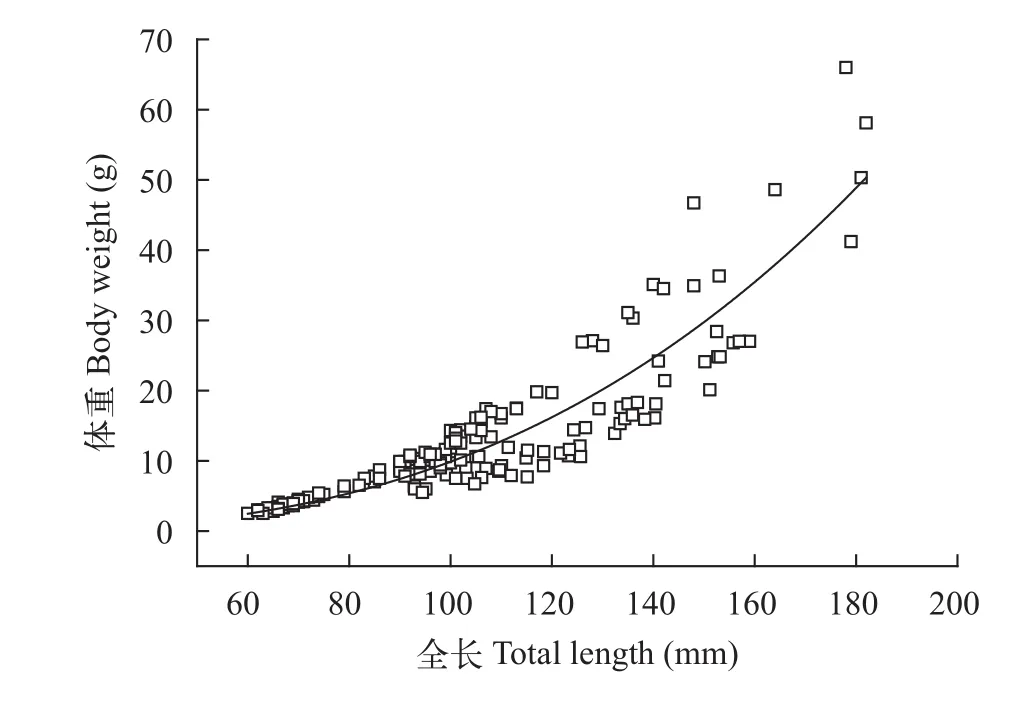
图10 大鳍异鲦全长与体重的关系Fig. 10 The relationship between total length and body weight of C. macropterus

图11 大鳍异鲦生长曲线Fig. 11 Growth curves of C. macropterus
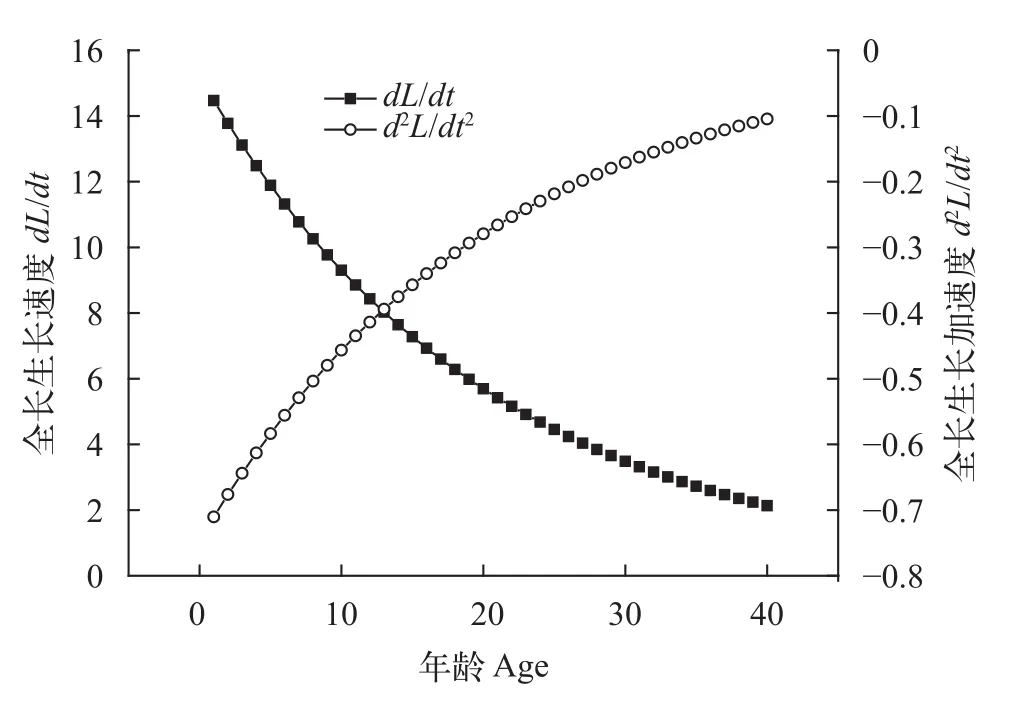
图12 大鳍异鲦全长生长速度和生长加速度Fig. 12 Growth rate and acceleration of total length of C.macropterus
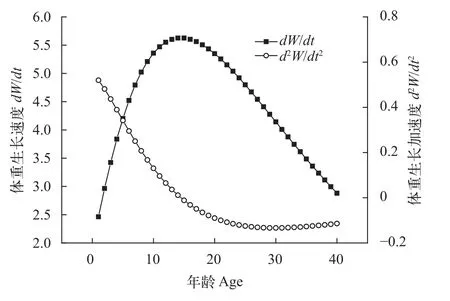
图13 大鳍异鲦体重生长速度和生长加速度Fig. 13 Growth rate and acceleration of body weight of C.macropterus
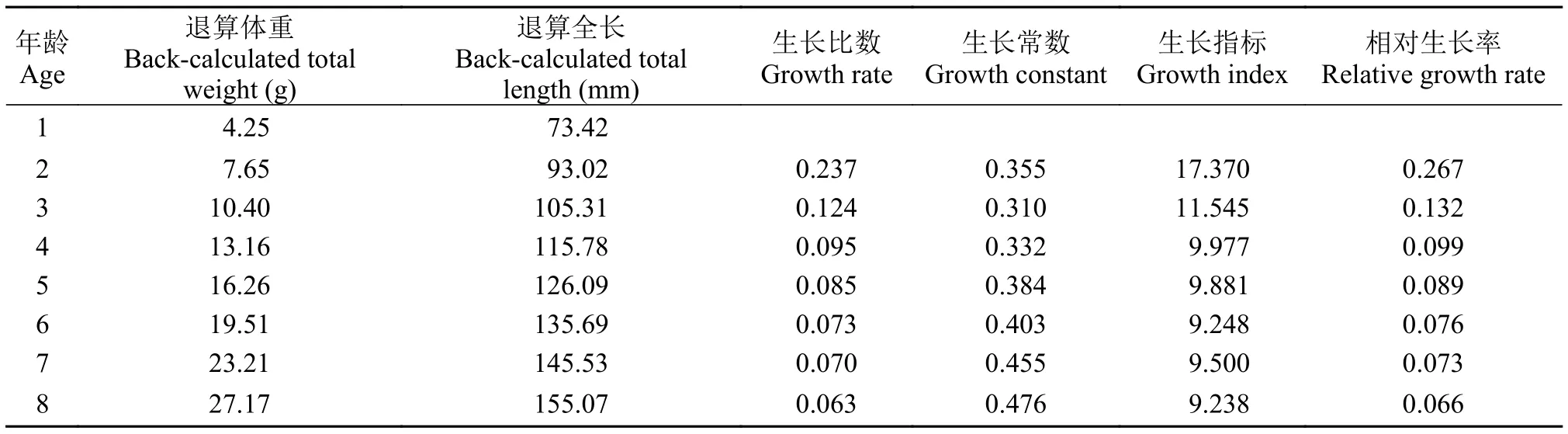
表 5 大鳍异鰔生长指数Tab. 5 Growth parameters of C. macropterus
3 讨论
3.1 年龄鉴定材料
鲇形目鱼类无鳞, 常用的年龄鉴定材料为脊椎骨、耳石、胸鳍棘等。谢小军[5]以脊椎骨为材料研究了南方鲇(Silurus meridionalis)的年龄和生长, 提出脊椎骨是鲇形目鱼类年龄和生长研究的好材料。以耳石作为年龄鉴定材料的有中华鲱鲇(Clupisoma sinensis)[6]、越鳠(Mystus pluriradiatus)[7]和切尾拟鲿(Pseudobagrus truncatus)[8]等。陈焜慈等[9]以鳃盖骨作为斑鳠(Mystus guttatus)的年龄材料。
Robert等[10]以耳石和胸鳍棘作为材料, 对斑点叉尾鲙(Ictalurus punctatus)年龄进行鉴定, 胸鳍棘与耳石鉴定结果基本一致, 认为二者均为理想的年龄材料。Penha等[11]以胸鳍棘为材料鉴定铲吻油鲇(Sorubim lima)年龄, 判定其为铲吻油鲇可靠的年龄材料。铲鲙(Pylodictis olivaris)[12]和扁吻半丘油鲇(Hemisorubim platyrhynchos)[13]也均以胸鳍棘为年龄鉴定材料。
王德寿等[14]以胸鳍棘作为大鳍鳠(Mystus macropterus)年龄材料, 通过简易石蜡切片观察其年龄结构, 并认为该方法适用于小型逘属鱼类及长吻逘(Leiocassis longirostris)等的幼鱼的年龄鉴定。在盔鲇(Cranoglanis sinensis)[15]、长吻逘[16]和南方鲇[17]的年龄研究中, 胸鳍棘与脊椎骨均具有很高的一致性。因此, 胸鳍棘应为鲇形目鱼类可靠的年龄鉴定材料。
大鳍异鲦胸鳍棘易采集, 实验操作简单, 可批量操作。横切面上年轮结构清晰, 副轮少, 易区别,是可靠的年龄鉴定材料。脊椎骨年轮清楚, 但材料处理和鉴定较繁琐。因此, 胸鳍棘是大鳍异鲦最佳的年龄鉴定材料。
3.2 生长特征
大鳍异鲦雌雄生长无显著差异, 这与大鳍鳠[18]、斑点叉尾鲙和铲鲙[19]相似, 与瓦氏黄颡鱼(Pelteobagrus vachelli)[20]不同。大鳍异鲦体重生长速度呈先上升后下降的特点, 全长生长速度则逐渐减小;体重生长加速度先下降, 由正转为负, 30龄后逐步回升, 趋近0, 全长生长加速度逐步增大, 并趋向0,生长速度和生长加速度随着年龄的增大, 变化趋势减缓。
鲇形目鱼类比较k为生长系数, 规定了生长曲线接近渐进值的速率,k越大意味着曲线接近渐进值越快, 生长速度越快[21]。k值为0.05—0.10的鱼类属生长缓慢种, 0.10—0.20属均生长种, 0.20—0.50属快速生长种[18]。如表 7所示, 大鳍异鲦、中华鲱鲇[6]、斑鳠[9]属生长缓慢种, 福建纹胸鲦(Glyptothorax fukiensis)[22]和雄性南方鲇[23]为快速生长种,雌性南方鲇为均生长种。大鳍异鲦k值为0.0491, 与其他鲇形目鱼类相比生长速度较为缓慢。
大鳍异鲦的生长拐点年龄为14.48龄, 较5龄的性成熟年龄大, 这与其他4种鲇形目鱼类类似。鱼类在性成熟以后, 其摄食所获得能量需优先供应于繁殖需求, 其次才供应于生长, 二者之间存在替代效应, 谢小军等[23]认为南方鲇在性成熟后保持快速生长的原因是其每年仅产一次卵, 繁殖投入较低,故生长投入较大, 而且, 南方鲇为肉食性鱼类, 捕食能力强, 能量摄入高。大鳍异鲦亦为一次产卵型的肉食性鱼类, 因此, 在性成熟后的几年内仍可保持快速的生长。
高原冷水鱼类比较青藏高原鱼类主要由3大类群组成: 裂腹鱼类、条鳅类、鲦科鱼类[24]。如表 8所示, 除雄性叶尔羌高原鳅(Triplophysa yarkandensis)[25]和雌性黑斑原鲦(Glyptosternum maculatum)[26,27]为均生长种外, 大鳍异鲦、拉萨裂腹鱼(Schizothorax waltoni)[28]、异齿裂腹鱼(Schizothorax oconnori)[29]、雌性叶尔羌高原鳅[25]和雄性黑斑原鲦[26,27]均为生长缓慢种。大鳍异鲦k值为0.0491,小于其他4种高原冷水鱼类, 生长速度最慢。
大鳍异鲦的生长拐点年龄为14.48龄, 较5龄的性成熟年龄大, 与叶尔羌高原鳅和黑斑原鲦类似,在性成熟后的几年内仍可保持快速的生长。拉萨裂腹鱼拐点年龄与性成熟年龄基本一致, 性成熟后生长速度减缓, 异齿裂腹鱼拐点年龄早于性成熟年龄, 即在性成熟前就达到了最大生长速度。
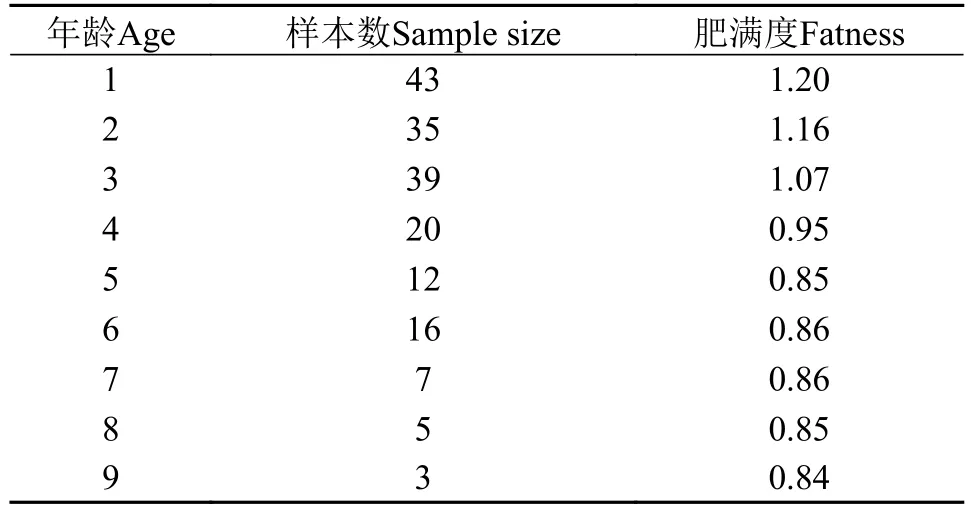
表 6 大鳍异鰔不同年龄的肥满度Tab. 6 Fullness of C. macropterus at different ages

表 7 五种鲇形目鱼类生长参数比较Tab. 7 Comparison of growth parameters of five Siluriformes species
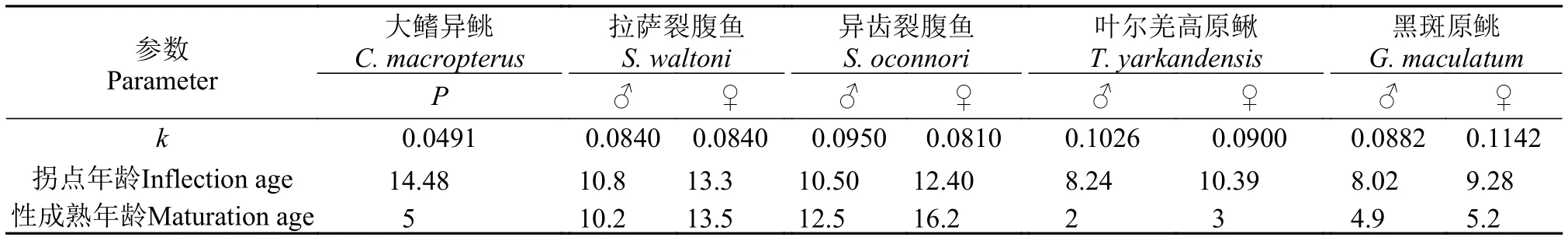
表 8 高原冷水鱼类生长参数比较Tab. 8 Comparison of growth parameters of cold-water fishes in plateau
3.3 资源保护
Musick[30]指出k值小于0.1即为长寿命鱼类, 长寿命的鱼类具有快速死亡、消亡的缺陷, 且资源快速锐减后的恢复需要数十年的时间。大鳍异鲦k值为0.0491, 生长较其他鲇形目鱼类和高原冷水鱼类都缓慢, 属于长寿命鱼类。近几年, 大鳍异鲦渔获物快速低龄化, 以3龄以下为主, 大量未达到性成熟的个体被捕捞, 这所造成的资源枯竭将极难恢复。因此, 合理捕捞, 资源复壮, 保护大鳍异鲦种质资源将是必要的。大鳍异鲦性成熟年龄为5龄, 且肥满度在5龄以后稳定, 故建议适度捕捞5龄以上个体,保证每年均有繁殖活动, 使得物种可以延续。
[1]Zhou W, Li X, Thomson A W. A new genus of glyptosternine catfish (Siluriformes: Sisoridae) with description of two new species from Yunnan, China [J].Copeia,2011(2): 226—241
[2]Ng H H. Two new glyptosternine catfishes (Teleostei:Sisoridae) from Vietnam and China [J].Zootaxa, 2004,428(1): 1—12
[3]Kong D P. Phylogeny and biogeography of Glyptosternoid fishes (Siluriformes: Sisoridae) [D]. Thesis for Doctor of Science. Kunming Institute of Zoology, Chinese Academy of Sciences, Yunnan. 2006 [孔德平. 襇鲦鱼类(Glyptosternoid fishes)的系统进化和动物地理学研究.博士学位论文, 中国科学院昆明动物研究所, 云南.2006]
[4]Yao J L, Yan Y Z, Gao Y,et al. Study on morphological variations ofPareuchiloglanis kamengensisand species validity ofP. macropterusin southwestern of China [J].Acta Zootaxonomica Sinica, 2007, 32(4): 814—821 [姚景龙, 严云志, 高勇, 等. 扁头鲦地理种群形态变异的研究兼论大鳍鲦的物种有效性. 动物分类学报, 2007, 32(4):814—821]
[5]Xie X J. A study on the age and growth ofSilurus soldatovi meridionalisin the Jialing River [J].Acta Ecologica Sinica, 1987, 7(4): 359—367 [谢小军. 嘉陵江南方大口鲶的年龄和生长的初步研究. 生态学报, 1987, 7(4):359—367]
[6]Yin S Y, Liu M D, Tian H W,et al. Age discrimination and growth characteristics ofClupisoma sinensisin Lancang River [J].Chinese Journal of Zoology, 2011, 46(2):12—21 [阴双雨, 刘明典, 田辉伍, 等. 中华鲱鲇年龄鉴定及生长特征. 动物学杂志, 2011, 46(2): 12—21]
[7]Tian Z F. Studies on age, growth and population protection ofMystus pluriradiatusin Yuanjiang River [D].Thesis for Master of Science. Huazhong Agricultural University, Wuhan. 2010 [田志福. 元江水系越鳠的年龄、生长与种群保护研究. 硕士学位论文, 华中农业大学,武汉. 2010.]
[8]Zou Y C, Wang Y M, Yue X J,et al. Histological studies of the digestive system inPseudobagrus truncates[J].Freshwater Fisheries, 2014, 44(2): 25—28 [邹远超, 王永明, 岳兴建, 等. 切尾拟鲿消化系统的组织学研究. 淡水渔业, 2014, 44(2): 25—28]
[9]Chen K C, Wu G M, Li H S,et al. Studies on the age and growth ofMystus guttatus[J].Journal of Fishery Sciences of China, 1999, 6(4): 62—66 [陈焜慈, 邬国民, 李恒颂, 等. 珠江斑鳠年龄和生长的研究. 中国水产科学,1999, 6(4): 62—66]
[10]Robert E C, Quinton E P, Candice M M,et al. Comparison of channel catfish age estimates and resulting population demographics using two common structures [J].North American Journal of Fisheries Management, 2010,30(1): 305—308
[11]Penha J M, Mateus L A, Barbieri G. Age and growth of the duckbill catfish (Sorubim cf. lima) in the Pantanal [J].Brazilian Journal of Biology, 2004, 64(1): 125—134
[12]Kirk L Y, Paul C M. Age and growth of Flathead Catfish in four southwestern rivers [J].California Fish and Game, 1990, 76(4): 224—233
[13]Penha J M, Mateus L A, Barbieri G. Age and growth of the porthole shovelnose catfish (Hemisorubim platyrhynchos) in the Pantanal [J].Brazilian Journal of Biology, 2004, 64(4): 833—840
[14]Wang D S, Luo Q S. A study on the age and growth ofMystus macropterus(Bleeker) [J].Acta Hydrobiologica Sinica, 1993, 17(2): 157—165 [王德寿, 罗泉笙. 嘉陵江大鳍鳠的年龄和生长的研究. 水生生物学报, 1993,17(2): 157—165]
[15]Zhang S G. Age and growth ofCranoglanis sinensis[J].Freshwater Fisheries, 1987, 3: 2—6 [张世光. 西江盔鲇年龄和生长研究. 淡水渔业, 1987, 3: 2—6]
[16]Wu Q J. Population ecology of [Leiocassis longirostris(Günther)](Pisces, Bagridae) with reference to the problem of maximum sustained yield [J].Acta Hydrobiologica Sinica, 1975, 5(3): 387—408 [吴清江. 长吻逘[Leiocassis longirostris(Günther)]的种群生态学及其最大持续渔获量的研究. 水生生物学集刊, 1975, 5(3):387—408]
[17]Wang Z L, Wu G X, Yang D G,et al. Age and growth ofSilurus meridionalisin the middle and upper reaches of the Yangtze River [J].Freshwater Fisheries, 1990, 6:3—9 [王志玲, 吴国犀, 杨德国, 等. 长江中上游大口鲇的年龄和生长. 淡水渔业, 1990, 6: 3—9]
[18]Wu J M, Zhang F T, Liu F,et al. Age and growth ofMystus macropterusin the Chishui River [J].Freshwater Fisheries, 2011, 41(4): 21—25 [吴金明, 张福铁, 刘飞,等. 赤水河大鳍鳠的年龄与生长. 淡水渔业, 2011,41(4): 21—25]
[19]Matthew D M, Michael J M, Michael P H. Age and growth variability between sexes of three catfish species in Lake Wilson, Alabama [J].North American Journal of Fisheries Management, 2009, 29(5): 1283—1286
[20]Duan Z H, Sun J Y. Studies on the age and growth ofPelteobagrus vachelli(Richardson) [J].Acta Hydrobiologica Sinica, 1999, 23(6): 317—623 [段中华, 孙建贻. 瓦氏黄颡鱼年龄与生长的研究. 水生生物学报, 1999, 23(6):317—623]
[21]Yin M C. Ecology of Fishes [M]. Beijing: China Agriculture Press. 1995, 57—58 [殷名称. 鱼类生态学. 北京: 中国农业出版社. 1995, 57—58]
[22]Shen Y J, Pu D Y, Gao Mei,et al. Studies on the relationship between the age and growth ofGlaptothorax fukiensisin the Jialing river [J].Journal of Southwest Agricultural University, 2005, 27(1): 106—110 [申严杰, 蒲德永, 高梅, 等. 福建纹胸鲦年龄与生长的初步研究. 西南农业大学学报(自然科学版), 2005, 27(1): 106—110]
[23]Xie X J, Long T D, Cao Z D. Studies on the composition and growth in the reproductive population ofSilurus meridionalis[J].Journal of Southwest China Normal University(Natural Science), 1994, 19(1): 71—78 [谢小军,龙天澄, 曹振东. 南方鲇的繁殖群体的结构及生长. 西南师范大学学报(自然科学版), 1994, 19(1): 71—78]
[24]Wu Y F, Tan Q J. Characteristics of the fish-fauna of the characteristics of Qinghai-Xizang plateau and its geological distribution and formation [J].Acta Zoologica Sinica, 1991, 37(2): 135—152 [武云飞, 谭齐佳. 青藏高原鱼类区系特征及其形成的地史原因分析. 动物学报, 1991,37(2): 135—152]
[25]Chen S S. Study on population ecology ofTriplophysa yarkandensis(Day) in Tarim river [D]. Thesis for Master of Science. Huazhong Agricultural University, Wuhan.2012 [陈生熬. 塔里木河叶尔羌高原鳅种群生态学研究.硕士学位论文, 华中农业大学, 武汉. 2012]
[26]Li J H. Studies on the biology and population ecology ofGlyptosternum maculatum(Regan) in the Brahmaputra River, China [D]. Thesis for Doctor of Science. Huazhong Agricultural University, Wuhan. 2008 [李红敬. 黑斑原鲦个体生物学及种群生态研究. 博士学位论文, 华中农业大学, 武汉. 2008]
[27]Li H J, Xie C X. Age and growth of the Tibetan catfishGlyptosternum maculatumin the Brahmaputra River,China [J].Zoological Studies, 2008, 47(5): 555—563
[28]Zhou X J. Study on the biology and population dynamics ofSchizothorax waltoni[D]. Thesis for Doctor of Science. Huazhong Agricultural University, Wuhan. 2014[周贤君. 拉萨裂腹鱼个体生物学和种群动态研究. 博士学位论文, 华中农业大学, 武汉. 2014]
[29]Ma B S. Study on the biology and population dynamics ofSchizothorax oconnori[D]. Thesis for Doctor of Science.Huazhong Agricultural University, Wuhan. 2011 [马宝珊. 异齿裂腹鱼个体生物学和种群动态研究. 博士学位论文, 华中农业大学, 武汉. 2011]
[30]Musick J A. Ecology and conservation of long-lived marine animals [C]. Symposium on conservation of longlived marine animals, Monterey. 1997
