长湖浮游动物群落结构及其与环境因子的关系*
2018-01-22罗静波杨德国何勇凤
郭 坤 彭 婷 罗静波 杨德国 何勇凤 柴 毅,①
(1. 长江大学动物科学学院 荆州 434025; 2. 中国水产科学研究院长江水产研究所农业部淡水生物多样性保护重点实验室武汉 430223)
长湖是湖北省第三大天然淡水湖泊, 地处荆州、荆门、潜江三市交界处, 是长江中游地区生态平衡的重要湿地生态系统(何勇凤等, 2016), 农业部于 2010年正式下文批准在长湖建立鲌类国家级水产种质资源保护区。长湖周围为居民生活区, 包括水产养殖、农业施肥和工业生产在内的人为因素已经极大的影响了长湖的水质与生态平衡。2006—2011年的水质监测数据显示, 长湖综合平均水质为Ⅴ类, 整体处于轻度富营养状态(刘建峰等, 2014)。
浮游动物是湖泊生态系统中重要的基础环节,对湖泊生态系统结构和功能的稳定起重要的调控作用: 一方面通过捕食作用控制浮游植物数量, 另一方面其作为鱼类等高营养级生物的饵料, 其丰度和生物量直接影响鱼虾蟹等的资源量(姜会超等, 2015)。然而, 有关长湖浮游动物的研究鲜有报道, 现有研究多集中于该区域浮游植物的群落结构变化及其与环境因子间的关系、水质变化、高等水生植物的分布等(吴翠等, 2008; 余明勇等, 2013; 柴毅等, 2014; 郝孟曦等, 2015)。本研究对长湖浮游动物(不包括原生动物)(陈非洲, 2016)的群落结构进行分析, 探讨其时空分布与环境因子间的关系, 以期对长湖水生生物资源的可持续发展与利用提供重要的参考。
1 材料与方法
1.1 采样点设置
参考《湖泊富营养化调查规范》(金相灿等, 1990)和《全国淡水生物物种资源调查技术规定》(试行)(国家环境保护总局自然生态司, 2010)进行采样点的设置。长湖自西向东分为庙湖区、海子湖区、马洪台区、圆心湖区四个湖区, 其中, “引江济汉”工程已将庙湖区拦截成静水, 因此舍弃该区域; 海子湖区为一狭形湖湾, 周围共有5个进出水口; 马洪台区、圆心湖区均为开阔水域, 且都有不同程度的围网养殖, 马洪台区同时作为航道。共在长湖设置 19个采样点, 依次为海子湖区(H)9个(H1—H9), 马洪台区(M)4个M1—M4, 圆心湖区(Y)6个 Y1—Y6, 详见图 1。于2012年 4月(春季)、2012年 7月(夏季)、2012年 10月(秋季)、2013年1月(冬季)进行采样。
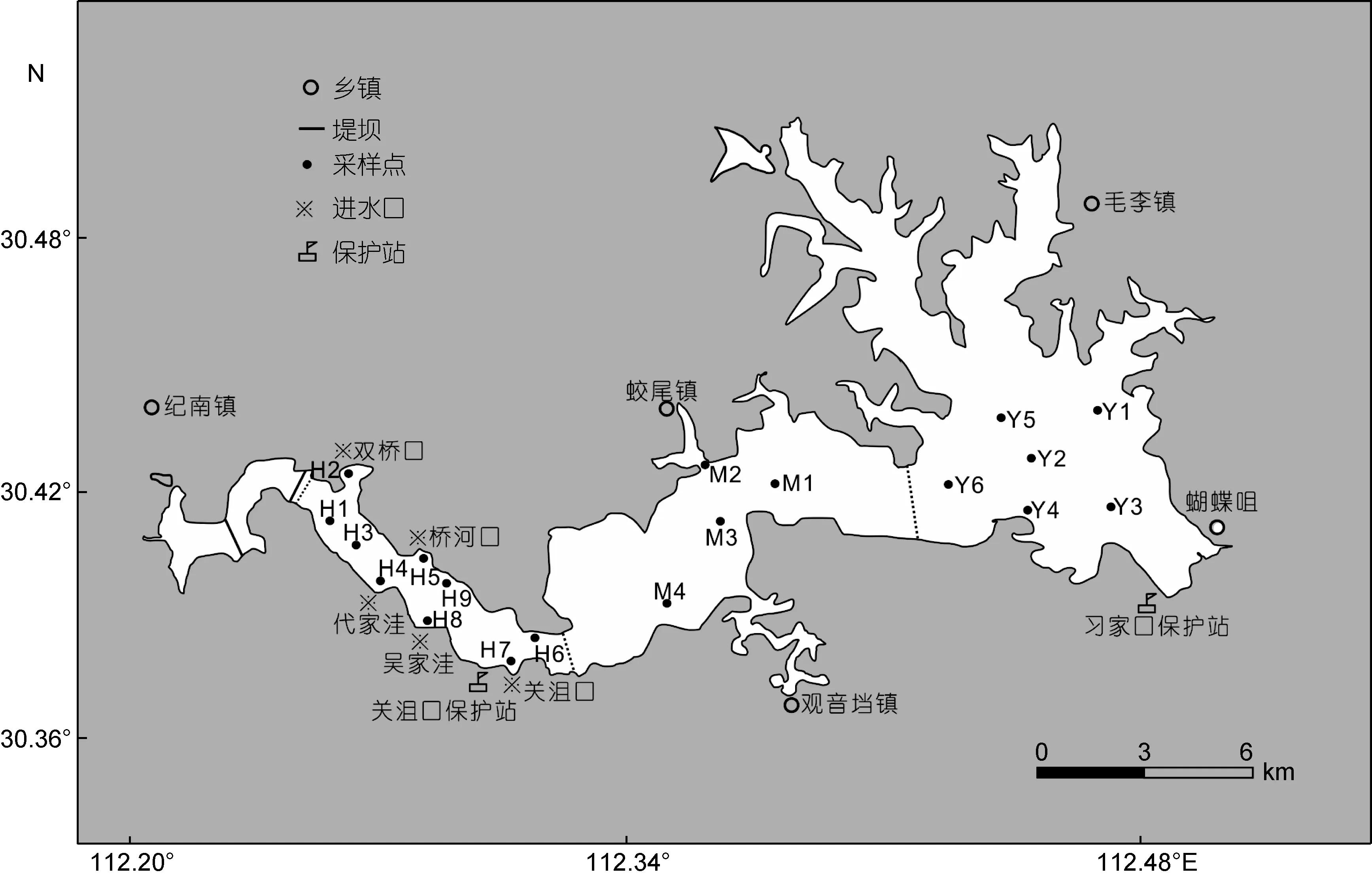
图1 长湖采样点示意图Fig.1 The sampling sites in Changhu Lake
1.2 浮游动物样本采集
拖动 13#浮游生物网在水面以下做“∞”状运动数次, 将浓缩的水样收集于50mL标本瓶中, 使用4%的甲醛溶液固定后获得定性样本。使用5L采水器分别于上中下水层采集两次, 混合均匀后使用 25#浮游生物网浓缩至 100mL, 用 4%的甲醛固定后获得枝角类和桡足类的定量样本。轮虫、无节幼体的定量直接使用浮游植物的定量样品。
参考《中国淡水轮虫志》(王家楫, 1961)、《中国动物志淡水枝角类》(蒋燮治等, 1979)、《中国动物志桡足类》(中国科学院动物研究所甲壳动物研究组,1979)在实验室中对样本进行种类的鉴定与计数, 生物量为固定样品后称得的湿重(mg/L)。
1.3 水体理化因子测定
采集样本的同时对水质理化指标进行检测, 使用黑白盘测定透明度(SD), 使用多参数水质分析仪(HQ40D, Hach, Loveland, USA)测定溶氧(DO)、pH、水温(T)。采集水样在实验室测定总磷(TP)、总氮(TN)、化学耗氧量(COD)、叶绿素(chl a)和总悬浮物(TSS),所有指标的测定参照《地表水和污水监测技术规范》(国家环境保护总局, 2003)中的方法进行。
1.4 数据处理方法
浮游动物群落多样性指数的测定: 以浮游动物每升出现的个体数(ind./L)来测定其丰度, 使用优势度指数 Y确定优势种的种类。使用 Shannon-wiener指数(H′)、Margalef指数(dMa)、Pielou均匀度指数(e)测定物种的多样性, 并依此评价长湖水质现状。各计算公式为:
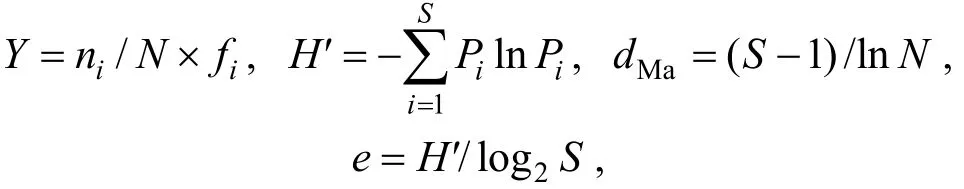
式中, Y是优势度, ni是第i种的丰度, N是该区域出现的所有种类的总丰度, fi是第 i种出现的频率,Y>0.02为优势种; Pi是ni与N的比值, S是该站动物的种类数。
使用肥度指数 E对长湖的富营养状态进行评价(杨亮杰等, 2014), E = K( x + 1 )/[(A + Y ) (y + 1 )], 其中K表示轮虫种类数, A表示桡足类种类数, Y表示枝角类种类数, x表示富-中营养型指示种, y表示贫-中营养型指示种。
季节更替率R: R = ( a + b - 2 c) /(a + b - c )× 1 00%,其中a与b分别为相邻季节的种数, c为相邻季节共享的种数。
浮游动物丰度、水质指标的区域性及季节性差异的检验使用单因素方差分析, 两两之间的比较使用多重比较完成。使用相关性分析、多元逐步回归分析和冗余分析(Redundancy Analysis, RDA)探索影响浮游动物群落结构的环境因子。
以上所有计算均在 R_V3.24中完成, 主要使用vegan包中的相关函数进行多样性指数的计算及RDA排序分析和制图(Borcard et al, 2014), 优势度指数、肥度指数和季节更替率使用自编程函数实现。参照《统计建模与R软件》(薛毅等, 2007)进行单因素方差分析、多重比较、相关性分析及多元逐步回归等计算, 分布图的制作在Excel2016中完成。
2 结果与分析
2.1 理化因子时空变化
水质指标的时空分布箱线图见图 2, 所有水质因子季节性差异显著, pH、T、COD四个季节均无显著区域性差异, 其他因子仅在个别季节表现出区域性差异。
2.2 浮游动物群落结构与多样性分析
2.2.1 浮游动物群落结构特征 此次调查共鉴定出浮游动物67种, 其中轮虫42种, 枝角类16种, 桡足类9种。各季节浮游动物种类数依次为春季(41种)>夏季(37种)>秋季(32种)>冬季(29种)。各季节多数区域轮虫在种类数、丰度及生物量上均占据较大优势,种类的更替率均大于 58.14%(春夏间), 最高达82.05% (冬春间)。

图2 长湖水质因子的季节性和区域性变化箱型图Fig.2 Spatio-temporal distribution of the main environmental factors in different parts of the Changhu Lake注: 每组依次为海子湖区(H)、马洪台区(M)、圆心湖区(Y); 箱型图从下到上的横线依次为下边缘、四分之一分位数、中位数、四分之三分位数、上边缘, 图外的点为异常点; a、b、c表示区域间的差异, A、B、C、D表示季节间的差异。
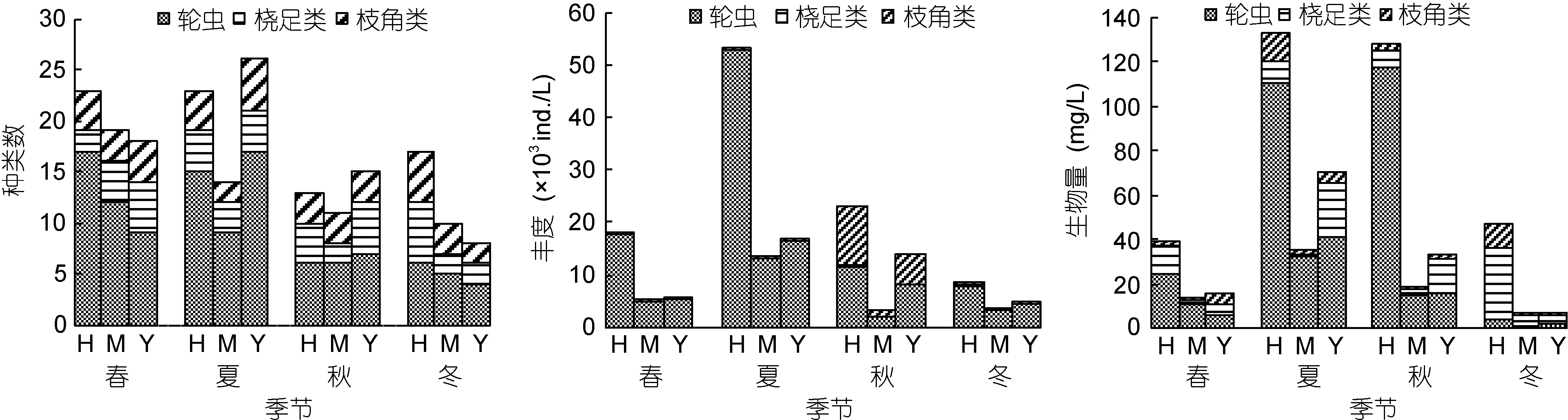
图3 浮游动物种类数、丰度及生物量的区域性及季节性分布Fig.3 Spatio-temporal distributions of zooplankton density and biomass注: H: 海子湖区; M: 马洪台区; Y: 圆心湖区
浮游动物丰度和生物量的区域性及季节性分布见图 3。浮游动物丰度的季节性差异显著(ANOVA,F=18.03, P=0.000), 且夏季((4412±2127)ind./L)>秋季((2111±1626)ind./L)> 春 季 ((1454±1016)ind./L)> 冬 季((831±606)ind./L), 仅夏季浮游动物丰度区域间差异显著(ANOVA, F=5.49, P=0.015), 且海子湖区丰度大于马洪台区、圆心湖区。各季节各区域丰度均以轮虫占优, 春、夏、冬季各区域轮虫的丰度占总丰度91%以上, 秋季占比也超过50%。夏季, 海子湖区的轮虫丰度达 5.31×104ind./L, 超过其他季节浮游动物的总丰度; 此时总丰度达 8.29×104ind./L, 为全年最大值,是其他季节总丰度的 2—5倍。枝角类丰度在秋季达到最大值, 总丰度为 1.83×104ind./L, 其中海子湖区枝角类的总丰度为1.15×104ind./L, 占比63.1%, 其他季节各区域枝角类丰度均小于370ind./L。桡足类的丰度除在冬季海子湖区达到 398ind./L, 其他季节各区域的丰度均小于99ind./L。
各季节总生物量依次为夏季(238.2mg/L)>秋季(180.7mg/L)>春季(69.09mg/L)>冬季(60.64mg/L), 生物量的季节性差异不显著(ANOVA, F=0.887,P=0.452)。各区域除冬季以桡足类的生物量占优(60%左右)外, 其他季节轮虫生物量仍占较大的比例, 春、夏、秋季海子湖区轮虫生物量占比超过62.05%, 马洪台区超过80.19%, 圆心湖区超过36.56%。
2.2.2 浮游动物优势种 使用优势度指数 Y筛选出优势种8种, 均是轮虫, 枝角类和桡足类不形成优势种, 优势种种类及在各季节出现的频率、优势度指数见表1, 浮游动物优势种在各样点的丰度分布见图4。
各季节的优势种都较为单一, 优势度与丰度相对集中。萼花臂尾轮虫、针簇多肢轮虫是四个季节均出现的优势种, 其中, 针簇多肢轮虫四季的优势度和出现频率均是各个季节的最大值。
2.2.3 浮游动物多样性指数分布及水质评价 长湖四季浮游动物多样性指数的样点分布及对应的水质评价见图5。ANOVA分析显示这三种指数均无显著季节性差异, Shannon指数、Margalef指数仅在冬季区域性差异显著, 且均表现为海子湖区大于马洪台区、圆心湖区。肥度指数年季节性和区域性差异均不显著。
2.3 影响长湖浮游动物群落结构的关键环境因子分析
2.3.1 浮游动物与环境因子相关性分析 浮游动物与环境因子的相关性结果分析见表2。表中数据显示TSS、pH、T、TN与上述三个指标呈显著正相关, DO与之呈显著负相关。
应用多元逐步回归分析筛选出对长湖浮游动物丰度、种类数、和生物量变化相对重要的水质因子,所有数据均进行 lnX转化, 各回归方程及对应的 F值、P值见表3, F值与P值显示各回归方程回归极显著, 表明结果可靠。回归分析的结果显示: T在三个回归方程中都入选, TN在浮游动物丰度与浮游动物种类数的回归方程中入选。
2.3.2 浮游动物优势种与环境因子RDA分析 为进一步分析影响长湖浮游动物群落结构变化的重要因子, 首先对经 ln(x+1)转化后的四季优势种数据做除趋势对应分析(DCA), 结果显示各 DCA分析中四个轴的最大梯度均小于3(春夏秋冬依次为1.31、0.67、0.92、0.63), 因此再选取线性模型中的 RDA分析对同时经过 ln(x+1)转化后的优势种与环境因子数据进行探索, 获得优势种-环境因子相关双序图, 各季节RDA相关双序图见图6。对RDA分析结果进行置换检验, 设置置换次数为 1000, 即显著水平达 0.001。各季节RDA分析前两轴的特征根、解释率、累计解释率与置换检验结果数据见表4。
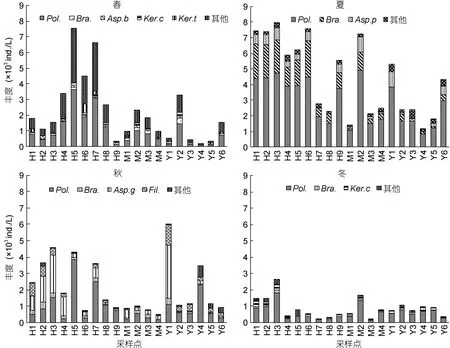
图4 长湖浮游动物优势种丰度样点分布Fig.4 Seasonal distribution of zooplankton density in each sampling sites in Changhu Lake注: 图中种类缩写见表1

表1 长湖浮游动物优势种Tab.1 The dominant species of zooplankton in Changhu Lake

表2 浮游动物丰度、种类、生物量与环境因子相关性分析(显著性水平α=0.05)Tab.2 Correlation analysis between zooplankton abundance, biomass, species number and environmental factors (significance levelα=0.05)
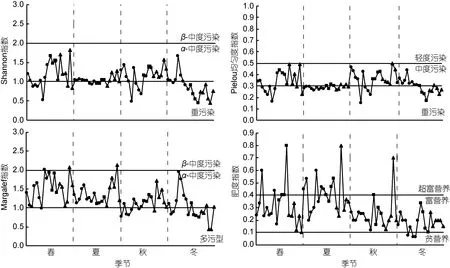
图5 长湖浮游动物Shannon指数、Pielou指数、Margalef指数、肥度指数四季样点分布Fig.5 Distribution of the Shannon index, Pielou index, Margalef index and E index of zooplankton in each sampling sites in Changhu Lake注: ●、■、▲分别代表海子湖区、马洪台区和圆心湖区的样点

表3 浮游动物与环境因子多元逐步回归方程Tab.3 Stepwise multiple regression among zooplankton and environmental factors

表4 各季节RDA分析前两轴特征根、解释率、累计解释率及置换检验结果数据Tab.4 Summary statistics of RDA
表 4的数据显示, 前两轴的累计解释率均高于42.50%, 且置换检验显示前两轴的结果是显著的, 因此认为RDA排序结果较好的解释了浮游动物优势种与环境因子间的关系。RDA相关双序图的解读遵循如下规则: 响应变量(物种)与解释变量(环境因子)箭头之间的夹角余弦值反映它们之间的相关性, 响应变量之间、解释变量之间作同样的解读。基于上述规则对图6进行解读。
3 讨论
3.1 长湖浮游动物群落结构分布特征及水质评价
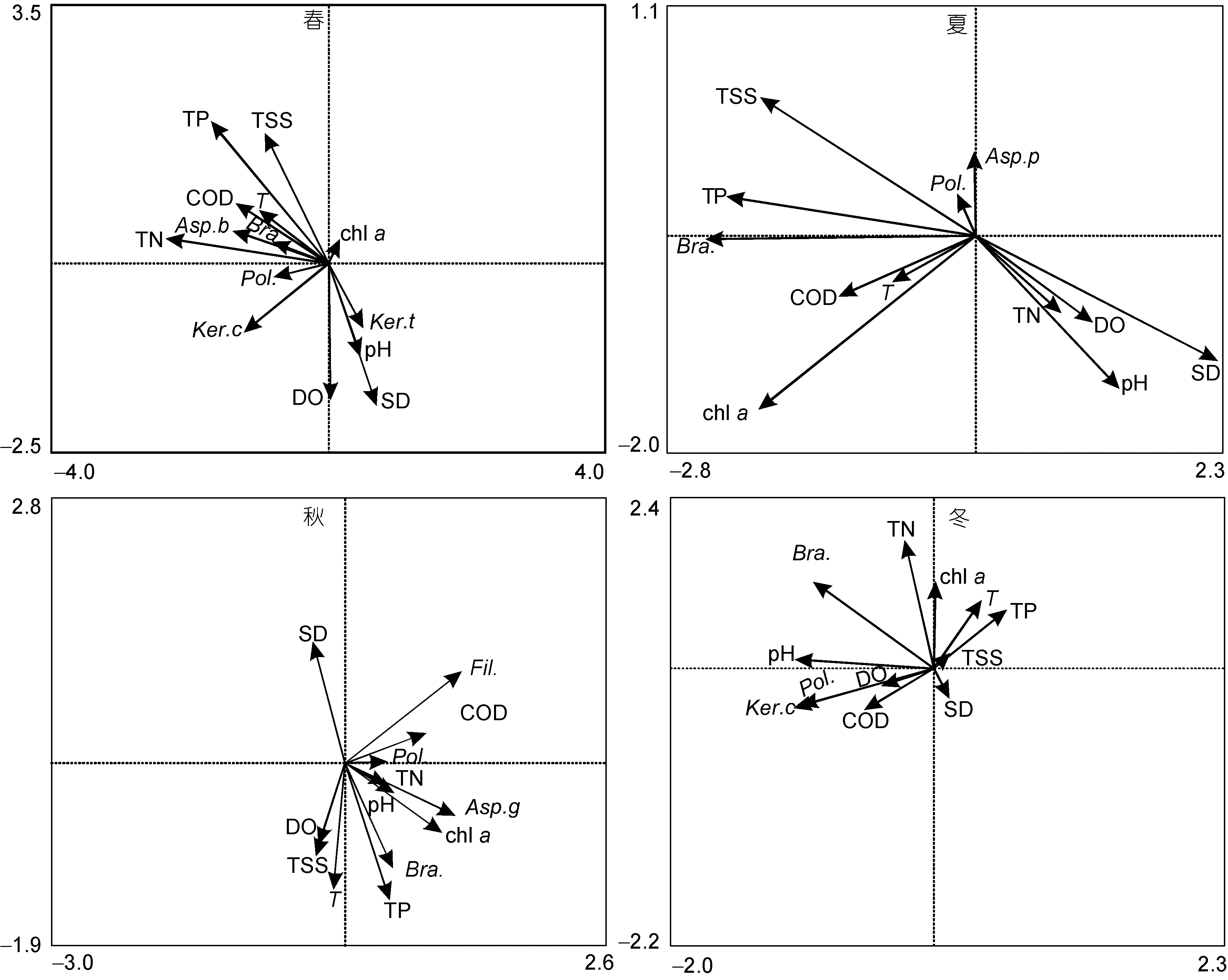
图6 各季节优势种-环境因子相关双序图Fig.6 Bi-plot of redundancy analysis between zooplankton species and environmental factors for each season
长湖浮游动物群落组成中, 小型的轮虫在种类数、丰度方面均占据较大优势, 大型浮游动物(桡足类、枝角类)仅在个别样点出现, 对丰度的贡献很小,且不形成优势种, 这一特点与国内其他湖泊的研究基本相同(吴利等, 2011; 靳萍等, 2013)。浮游动物丰度的季节性差异显著, 呈现为春、夏、秋季高于冬季,这与滴水湖浮游动物的群落结构的研究相同(林青等,2014)。季节间的更替率均大于59%, 但优势种种类的季节变化不明显。各季节浮游动物丰度的多重比较显示仅夏季海子湖区的丰度大于圆心湖区, 其他季节无显著区域性差异, 这可能与浮游动物优势种的结构有关: 浮游动物优势种共8种, 各季节优势种的丰度占总丰度的比值最小为 59.13%, 最高达 95.32%;萼花臂尾轮虫、针簇多枝轮虫是四个季节的共有优势种, 这两种优势种的丰度和与各季节总丰度的比值超过49.68%, 与各样点总丰度的比值最高达96.48%;对各季节这两种优势种的丰度和做多重比较, 结果发现仅夏季显示出海子湖区大于圆心湖区(P=0.041),这表明优势种占据较大优势, 萼花臂尾轮虫和针簇多枝轮虫在各季节湖区间的分布较为均匀。
应用多样性指数评价长湖水质的结果表明, 长湖春、夏、秋季多数样点处于中度污染, 冬季约一半的样点处于重污染。肥度指数评价水质的结果认为长湖全年多数样点处于富营养状态, 且夏季富营养程度有增大的趋势, 这与何勇凤等(2015)应用综合营养指数法与浮游植物细胞丰度指数法对长湖水质评定的结果相同。
3.2 长湖浮游动物群落结构的影响因子
温度一直被认为是影响浮游动物群落结构的重要环境因子之一, Record 等(2010)的研究认为浮游动物多样性季节动态变化主要与温度有关; Ekvall等(2012)的研究认为, 气候变暖会改变浮游动物优势种的结构, 且相较于桡足类, 枝角类更容易占优。高原等(2014)对珠江三角洲浮游轮虫的群落结构研究结果显示水温与轮虫的密度呈显著相关性。轮虫总数的高峰一般在水温20°C出现, 在15—25°C内, 绝大部分轮虫的发育时间会随温度升高而缩短, 轮虫总数高峰一般在20°C以上时出现(黄祥飞等, 1985)。本研究中, 轮虫丰度的年变化趋势与温度的年变化趋势相同, 丰度的最高峰出现在温度最高的夏季; 相关性分析显示浮游动物种类数、丰度、生物量均与水温呈极显著性正相关, 多元逐步回归方程表明水温对这三种指标均产生正向作用; RDA双序图则显示不同季节优势种与水温间的关系并不相同, 但作为主要优势种的萼花臂尾轮虫除冬季与 T的相关性约为零外,其他季节均与水温呈正相关。同时, 水温升高会使浮游植物的密度与生物量增加, 进而使得浮游动物获得充足的食物。故认为水温是影响长湖浮游动物群落结构季节变化的关键因子。
浮游动物的种类与数量与 pH值密切相关, 轮虫在碱性水体中数量多、种类少, 在酸性水体中的分布则相反; 枝角类一般生活在酸性、中性及微碱性的水体中; 桡足类大多比较适宜在偏碱性的水体中生存(高原等, 2013)。长湖全年各区域pH值相差不大, 属于偏碱性水体, 相关性分析显示浮游动物的种类数、丰度、生物量与pH呈正相关。主要优势种萼花臂尾轮虫能耐受较高 pH, 其致死 pH上限接近 11.0(王金秋等, 1997), 其生物量与pH呈极显著正相关(R=0.32,P=0.004)。pH并未入选多元逐步回归方程, RDA分析中, 春夏季节多数优势种与 pH呈负相关, 秋冬季则正相反。可见, 能耐受较高 pH的萼花臂尾轮虫在长湖偏碱性的水体中有较好的适应性, 且pH对长湖浮游动物的组成有一定影响。
溶解氧的不足可以限制轮虫的生长(杨丽丽等,2011), 夏季长湖溶解氧是四季的最小值, 此时 M 区的溶解氧是全年最小值, 该区域的轮虫丰度也是该季的最小值, 相关性分析显示溶解氧与浮游动物种类数、丰度和生物量呈极显著性负相关, 夏季的RDA分析也显示溶解氧与优势种之间呈负相关, 因此认为夏季M区较低的溶解氧抑制了轮虫的生长。
悬浮物对浮游动物的影响可能是对浮游动物的直接作用, 也可能是通过降低溶氧继而影响浮游植物的生长间接作用(张艳等, 2014)。本研究中TSS与浮游动物种类数、丰度、生物量呈显著正相关, 未入选多元逐步回归方程, RDA分析中多数优势种与之呈正相关, 且chl a与之呈正相关; TSS与DO呈极显著负相关(R=-0.494, P=0.000), 夏季RDA分析中TSS与DO的相关性几乎为-1, 这进一步解释了夏季M区较低的轮虫丰度。因此认为 TSS主要通过间接作用影响长湖浮游动物群落结构。
浮游植物是浮游动物的重要食物来源, 水体中浮游植物的数量与浮游动物的生长繁殖密切相关。不同浮游动物摄取的浮游植物种类也不相同, 萼花臂尾轮虫多数情况下可以利用绿藻和团藻, 基本不食用蓝藻, 针簇多枝轮虫偏向以裸藻、金藻和硅藻为食。长湖四季浮游植物群落结构的研究发现绿藻门的小球藻与硅藻门的扭曲小环藻是主要优势种, 且小球藻的优势度是春、秋、冬季的最大值, 蓝藻门的两栖颤藻在夏季大量增殖成为主要优势种, 氮磷比是影响浮游植物优势种分布的重要因子(郭坤等, 2016)。这与本研究的结果是基本吻合的: 相关性分析显示游动物种类数、丰度与浮游植物现存量的表征值chl a呈现正相关关系, RDA分析中, 萼花臂尾轮虫在四季与chl a呈正相关, 其他季节多数优势种均与chl a呈负相关, 而TP在四季与chl a呈正相关, TN在夏秋冬与chl a呈正相关。因此认为chl a是影响长湖浮游动物群落结构的重要因子, 而TN、TP是通过影响长湖浮游植物的群落结构间接影响浮游动物的组成的。
在分层水体, 透明度是影响轮虫分布的重要因子(吴利等, 2011), 而长湖为浅水湖泊, 不存在分层现象。化学需氧量是一种常用的评价水体污染程度的综合性指标, 化学需氧量越大, 说明水体受有机污染越严重。长湖浮游动物优势种中主要优势种针簇多枝轮虫及在春、冬季作为优势种的螺形龟甲轮虫均是耐污种, 多元逐步回归方程中化学需氧量对浮游动物丰度有正向作用, RDA分析中针簇多枝轮虫与其呈正相关, 而针簇多枝轮虫在各个样点的丰度超过多数种类, 因此认为化学需氧量对长湖浮游动物的丰度产生了正向作用。
4 结论
长湖浮游动物以轮虫为主, 其种类数、丰度及生物量均占据较大优势, 枝角类、桡足类不形成优势种。季节更替率高, 但优势种的变化不明显, 优势种在各个区域占据较大比例。萼花臂尾轮虫、针簇多枝轮虫是四季共有优势种, 且在湖区间的分布较为均匀。依据多样性指数认为长湖处于中度污染, 依据肥度指数认为长湖处于富营养状态, 且夏季有增重的趋势。
综合相关性分析、多元逐步回归及RDA分析的结果认为: 水温、chl a是直接影响长湖浮游动物群落结构的关键因子, 总氮、总磷通过影响浮游植物群落结构间接影响浮游动物的组成; 夏季 M 区较低的溶解氧抑制了轮虫的生长, 总悬浮物通过对降低溶解氧对浮游动物产生间接作用; pH与化学需氧量对特定的优势种影响较大。
本文仅对长湖浮游动物群落结构及影响其分布的关键水质因子做了初步探索, 高等水生植物的数量与分布、鱼类的摄食压力等也是不可忽略的因子,这些有待后续研究的补充。
王金秋, 李德尚, 罗一兵等, 1997. pH值对萼花臂尾轮虫种群增长及繁殖的影响. 应用生态学报, 8(4): 435—438
王家楫, 1961. 中国淡水轮虫志. 北京: 科学出版社, 21—289
中国科学院动物研究所甲壳动物研究组, 1979. 中国动物志——节肢动物门 甲壳纲 淡水桡足类. 北京: 科学出版社,53—420
刘建峰, 张 翔, 谢 平等, 2014. 长湖水质演变特征及水环境现状评价. 水资源保护, 30(4): 18—22
杨丽丽, 周小玉, 刘其根等, 2011. 新疆布伦托海轮虫群落结构及其与环境因子的关系. 湖泊科学, 23(5): 806—812
杨亮杰, 吕光汉, 竺俊全等, 2014. 横山水库浮游动物群落结构特征及水质评价. 水生生物学报, 38(4): 720—728
吴 利, 冯伟松, 张堂林等, 2011. 春、秋季武湖浮游动物群落特征及其与环境因子的关系. 水生态学杂志, 32(2): 31—37
吴 利, 冯伟松, 张堂林等, 2011. 湖北省西凉湖浮游动物群落周年动态变化及其与环境因子的关系. 湖泊科学, 23(4):619—625
吴 翠, 唐万鹏, 史玉虎等, 2008. 长湖湿地生态价值评价.湖北林业科技, (1): 45—47
何勇凤, 李昊成, 王旭歌等, 2016. 长湖鱼类群落结构的时空变化. 长江流域资源与环境, 25(2): 265—273
何勇凤, 李昊成, 朱永久等, 2015. 湖北长湖富营养化状况及时空变化(2012-2013年). 湖泊科学, 27(5): 853—864
余明勇, 张海林, 余向京, 2013. 长湖水环境需水量与引清济湖研究. 中国农村水利水电, (6): 21—25
张 艳, 郑 琳, 陈碧鹃等, 2014. 悬浮物对浮游植物和浮游动物的急性毒性效应. 渔业科学进展, 35(2): 16—21
陈非洲, 2016. 关于淡水浮游动物和原生动物分开研究的建议.湖泊科学, 28(3): 691
林 青, 由文辉, 徐凤洁等, 2014. 滴水湖浮游动物群落结构及其与环境因子的关系. 生态学报, 34(23): 6918—6929
国家环境保护总局, 2003. 地表水和污水监测技术规范(HJ/T 91—2002). 北京: 中国环境科学出版社, 15—20
国家环境保护总局自然生态司, 2010. 全国淡水生物物种资源调查技术规定(试行). 北京: 国家环境保护部, 167—176
金相灿, 屠清瑛, 1990. 湖泊富营养化调查规范. 北京: 中国环境科学出版社, 25—28
郝孟曦, 杨 磊, 孔祥虹等, 2015. 湖北长湖水生植物多样性及群落演替. 湖泊科学, 27(1): 94—102
姜会超, 陈海刚, 宋秀凯等, 2015. 莱州湾金城海域浮游动物群落结构及与环境因子的关系. 生态学报, 35(22):7308—7319
柴 毅, 彭 婷, 李昊成等, 2014. 长湖海子湖夏季浮游植物群落结构及环境影响因子. 湖北农业科学, 53(19):4568—4573
高 原, 李新辉, 赖子尼等, 2014. 珠江三角洲河网浮游轮虫的群落结构. 应用生态学报, 25(7): 2114—2122
高 原, 赖子尼, 李 捷等, 2013. 连江浮游动物多样性的空间分布. 生物多样性, 21(6): 699—708
郭 坤, 杨德国, 彭 婷等, 2016. 湖北省长湖浮游植物优势种生态位分析. 湖泊科学, 28(4): 825—834
黄祥飞, 胡春英, 伍焯田, 1985. 武汉东湖的轮虫. 水生生物学报, 9(2): 129—143
蒋燮治, 堵南山, 1979. 中国动物志——节肢动物门 甲壳纲淡水枝角类. 北京: 科学出版社, 79—273
靳 萍, 代克岩, 杨 程等, 2013. 牧野湖浮游动物群落结构及其与环境因子的相关性研究. 水生态学杂志, 34(2):53—61
薛 毅, 陈立萍, 2007. 统计建模与 R软件. 北京: 清华大学出版社, 1—371
Borcard D, Gillet F, Legendre P编著, 赖江山译, 2014. 数量生态学——R 语言的应用. 北京: 高等教育出版社,138—177
Ekvall M K, Hansson L A, 2012. Differences in recruitment and life-history strategy alter zooplankton spring dynamics under climate-change conditions. PLoS One, 7(9): e44614
Record N R, Pershing A J, Jossi J W, 2010. Biodiversity as a dynamic variable in the Gulf of Maine continuous plankton recorder transect. Journal of Plankton Research, 32(12):1675—1684
