引起核桃种子休眠的主要因素及其休眠类型
2018-01-22阿卜杜许库尔牙合甫古丽尼沙沙依明尼亚孜海利力库尔班吐逊古丽托乎提加帕尔卡迪尔
阿卜杜许库尔·牙合甫,古丽尼沙·沙依明尼亚孜,海利力·库尔班, 吐逊古丽·托乎提,何 丹,加帕尔·卡迪尔
(1.新疆林业科学院经济林研究所,乌鲁木齐 830063;2.吐鲁番地区鄯善县林业局,新疆鄯善 838200; 3. 新疆安居丹生物科技有限公司,乌鲁木齐 830011;4.新疆农业大学园艺学院,乌鲁木齐 830052)
0 引 言
【研究意义】核桃(JuglansregiaL.)也称胡桃,胡桃科(Juglandaceae),胡桃属(Juglans L.)[1]。全世界胡桃属(JuglansL.)有21种,主要分布于南北美洲,西印度,以及西南欧洲至东亚和日本等地区[2]。核桃属植物原产于我国的有4个种,即普通核桃(J.regiaL.)、核桃楸(J.mandshuricaMaxim.)、铁核桃(J.sigillataDode.)、河北核桃(J.hopeiensisHu.)[3]。新疆是我国核桃的主产区之一,该地区拥有丰富的种质资源和广阔的适合发展核桃产业的空间(新疆野生核桃是栽培核桃的直系祖先[4])。而发展核桃育种及苗木标准化繁育,需从最基本的种子生理研究开始。核桃种子春播经预处理,才能达到较好的出苗效果。因此,对核桃种子休眠方面的研究能在一定程度上弥补前人研究成果的同时,说明休眠的成因,对高效繁殖实生苗、保护核桃种质资源有重要的实践意义。【前人研究进展】种子是种子植物所特有的延存器官,其休眠和萌发是植物体重要的生命活动之一。种子休眠通常是指完整的具生活力的种子在适宜萌发的条件下不萌发(发芽)的现象[5]。虽然种子休眠是植物在长期演化过程中形成的一种对不良环境的适应机制。很多果树、林木、药材和某些农作物的种子,在自然条件下需要经过很长一段休眠期才能发芽,因此未经解除休眠的种子不能萌发或发芽率很低,往往给生产造成一定困难[6-8]。根据休眠的机制可将种子休眠类型分为物理休眠、化学休眠、生理休眠3种,其原因大致可归为两大类:第一类是内源因素,即胚本身的原因,包括胚的形态发育未完成、生理上未成熟、缺少必需的激素或存在抑制萌发的物质;第二类是外源因素,即胚以外的各种组织,即种壳(种皮、果皮或胚乳等)的限制,包括种壳的机械阻碍、不透水性、不透气性以及种壳中存在抑制萌发的物质等[5,9]。有研究认为,休眠与体内ABA的浓度以及有机体对抑制物质的敏感度有关[10],向日葵种胚内生的ABA能诱导休眠[11]。Schmitz 等(2002)认为,改变ABA代谢途径以及胚对ABA敏感性下降是逊扁柏(ChamaecyparisnootkatensisSpach)种子终止休眠的两个因素,但仅改变这两个因素之一不足以提高萌发率。化学处理可能直接激活ABA氧化酶,而通过低温层积或GA3影响ABA受体或影响下游信号传递,进而调控水解酶等生理过程[10,12]。解除种子休眠方法大体可以分为物理、化学和生物方法3类,物理方法包括温度处理、机械处理、射线、超声波处理和电场处理、干燥后熟和层积处理;化学处理包括激素处理、无机化学药剂处理和有机化学药物处理。生物方法包括低温层积、胚胎后熟等[5,13]。【本研究切入点】对核桃种子休眠成因和可能引起核桃种子休眠的GA3/ABA比值、抑制物质的存在,胚的后熟以及硬壳种皮的障碍等诸多因素中的主导因素及其休眠类型至今了解甚少。研究对核桃种子进行短期层积处理、外源赤霉素处理、清水浸泡处理和浸水后日光暴晒处理等多种处理,再结合核桃壳浸泡液进行生物检测等试验,对数据进行综合分析,揭示引起核桃种子休眠的主要因素并确定核桃种子休眠的所属类型。【拟解决的关键问题】探明核桃种子休眠的成因并确定核桃种子的休眠类型;解决目前农业生产所面临的播种前必需长达40 d以上的低温层积或必须在晚秋播种越冬的难题。
1 材料与方法
1.1 材 料
核桃种子属普通核桃(JuglansregiaL.),实生核桃种子,采集地喀什叶城县,经检验该批种子的饱满度为100%。
1.2 方 法
1.2.1 处理
首先将核桃种子随机分成6份、每份200粒核桃种子。设计5个处理,一个对照,未经处理直接播种为对照(CK);核桃干种子直接进行低温沙藏层积处理21 d为处理1(T1),处理条件为,含水量为70%、沙层厚度为10 cm、温度≤4℃;核桃种子用清水浸泡24 h再进行21 d低温沙藏为处理2(T2),处理条件同上;清水浸泡15 d 为处理3(T3),置于室温条件,每天换水1次;清水浸泡15 d后日光暴晒2 h为处理4(T4);核桃种子以浓度为480 mg/L的GA3溶液浸泡48 h为处理5(T5),处理温度为室温(15~20℃),并保证种子浸水均匀。
1.2.2 播种
将处理好的核桃种子4月初分别在温室播种到直径为20 cm的营养钵,自制营养土里。粘土+细沙+新鲜锯末以3∶1∶2混合制成营养土。每个营养钵种播种2粒核桃种子,把种子缝合线与地面垂直,种尖向一侧摆放,埋土厚度为5 cm。为避免水分蒸发过快,播种后浇透水营养钵上覆盖塑料薄膜,温室温度在20~25℃,每天观察湿度及发芽情况。
1.2.3 测定指标
核桃种子开始萌发后,每天统计发芽数,分别用公式1、2、3分别计算发芽率、发芽势和发芽指数[14]。发芽率是指样本种子中发芽种子的百分数;种子发芽势是指逐日发芽量已达到发芽高峰值,并将开始下降时即为发芽势的期限,以%表示[15]。发芽指数是发芽日数。分别用如下公式来计算:
公式1:

1.2.4 核桃壳浸泡液的制备及生物检测
去除核桃种仁保留壳(内果皮),将其按质量和体积的比例加入蒸馏水置于烧杯中,震荡浸泡24 h后过滤备用。将上述浸提液分别稀释制备0(对照)、0.5、1.0、1.5、2.0 g 干重/mL浓度的核桃壳浸泡液。吸取10 mL加入直径为9 cm培养皿中,放两张滤纸,整齐摆放10粒健康小麦种子,设3个重复,在25℃培养箱里萌发,每12 h记录发芽数(G)、芽长(BL)和根长(RL)至不再增加为止,并用公式1分别计算不同浓度下的小麦发芽率(%)。
公式2:
发芽势GE(%)=
公式3:
发芽指数(GI) = [T×G1+ (T-1) ×G2+ (T- 2) ×G3… + 1 ×GT]/T.
T表示需发芽完成而持续的时间(T=d),G1,G2,…GT表示在1, 2,…T内的发芽率;
1.3 数据处理
实验的样本量较大,数据采用两个总体比率的大样本做U检验方法来进行统计。两个总体比率的大样本做U检验方法公式4[16]:


n指样本量(供试种子数),X指样本的发生次数(发芽种子数)
当|U| ≥ 1.96时两个样本之间的差异显著,当|U| ≥ 2.58时两个样本之间的差异极显著,当|U| ≤ 1.96时两个样本之间的差异不显著。
统计分析采用SPSS13.0软件对小麦种子发芽率、芽长及根长的抑制率进行ANOVA分析,LSD多重比较,Excel2010绘图。
2 结果与分析
2.1 不同处理对种子发芽率的影响
核桃种子发芽率在不同处理之间存在明显的差异。研究表明,T4的种子发芽率最高,播种第50 d达到69.5%,比CK(34%)高104%,比其他处理及CK均有显著差异(|U| ≥2.58)。其次为T3,播种第50 d的发芽率比CK高 67.6%,比其他处理及CK均有显著的差异(|U| ≥2.58)。处理T5的种子发芽率发现为最低,在播种第50 d的发芽率仅为44.3%,与CK 、T1和T2之间均没有显著差异(|U| ≤ 1.96)。图1,表1

图1 不同处理下种子发芽率
Fig.1 The seed germination percentage of different treatment表1 核桃种子发芽率U检验结果(U值)
Table 1 U test results (U value) of walnut seed germination percentage
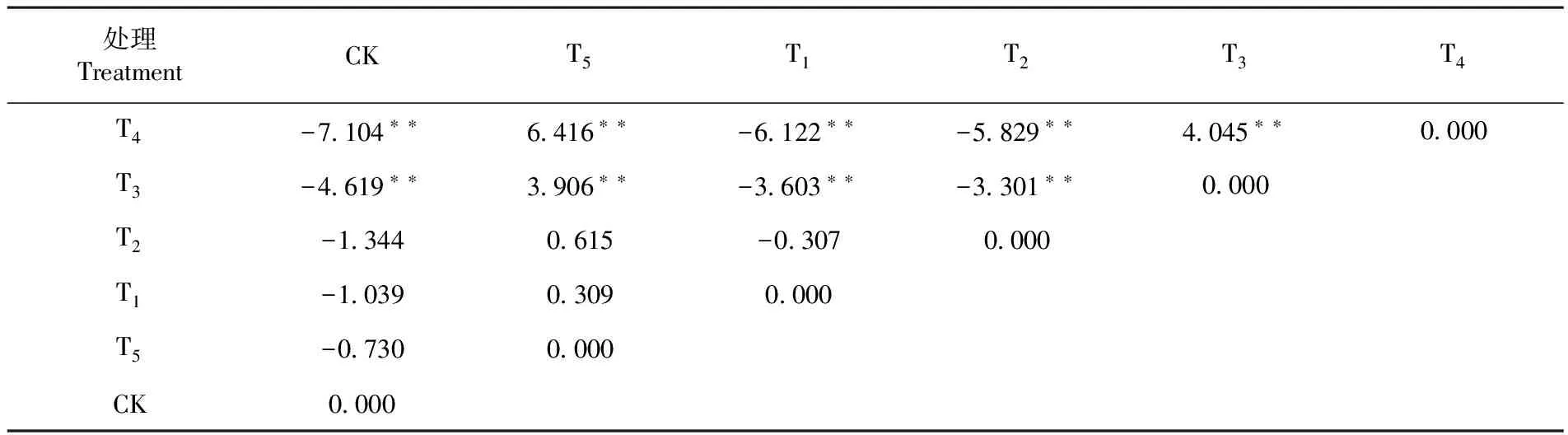
处理TreatmentCKT5T1T2T3T4T4-7104∗∗6416∗∗-6122∗∗-5829∗∗4045∗∗0000T3-4619∗∗3906∗∗-3603∗∗-3301∗∗0000T2-13440615-03070000T1-103903090000T5-07300000CK0000
注:当|U| ≥ 1.96时两个样本之间的差异显著,|U| ≥ 2.58时差异极显著,|U| ≤ 1.96时差异不显著。**指极显著性差异
Note: |U| ≥ 1.96 means significant difference at the 0.05 level, |U| ≥ 2.58 means significant difference at the 0.01 level, |U| ≤ 1.96 means there no significant difference.**means significant difference at the 0.01 level
2.2 不同处理下的种子发芽势和发芽指数
发芽势可以表示种子的发芽能力、反映种子活力的一个重要指标,而种子发芽指数是发芽日数,发芽指数高就说明该种子发芽所用的时间短、发芽速度快。研究表明,不同处理下的种子发芽势和发芽指数均有明显的差异。其中,T4的发芽势和发芽指数均为最高,分别为48%和19.52%,比CK高100%和307%;其次为T3,种子发芽势和发芽指数分别为35.5%和13.93%、比CK高39.58%和190%。处理T5的发芽势和发芽指数均为最低,分别为23.5%和7.48%,与CK没有明显的差异。表2
2.3 不同处理对种子萌发起始时间影响
不同处理下不仅种子发芽率有差异,而且种子的萌发起始时间也明显的不同。研究表明,处理T4与T2的种子萌发起始时间最早(第20 d),比CK早16 d;其次为T3,比CK早15 d。而处理T1与T5的种子萌发起始时间最晚(第24 d)。表2
表2 不同处理下种子发芽势和发芽指数变化
Table 2 The effect of different treatment on germination energy and germination index

处理Treatment萌发起始时间Startingdayofgermination(d)发芽势Germinationpotential(%)发芽指数GerminationindexCK362448T124185828T2203451064T3213351393T420481952T524235748
2.4 核桃种壳浸泡液对小麦种子萌发和幼苗生长的影响
桃种壳(内果皮)浸提液对小麦种子萌发具有强烈的抑制作用,随处理浓度的提高发芽率有明显的下降趋势。当提取液浓度1.5 g/mL时小麦种子发芽率仅有25.56%(抑制率达74%),与CK(100%)和0.5 g/mL处理(67%)之间有极显著性差异(P< 0.01),与1.0 g/mL浓度处理间有显著差异(P< 0.05)。不同浓度浸提液对小麦幼苗生长同样有显著的抑制作用,当浓度1.5 g/mL时浸泡液对幼苗芽长和根长的抑制效果较对照和0.5 g/mL处理极显著高,即抑制率分别高达94%、95%,在此处理下幼苗芽长和根长的抑制效果与1.0 g/mL浓度间有显著差异(P< 0.05)。图2,表3
表3 核桃壳浸泡液对小麦种子发芽率和芽长及根长的抑制作用
Table 3 Inhibitory effect of walnut endocarp extracts on germination percentage and shoot and root length of wheat seeds

处理浓度Treat(g/mL)发芽率GP平均值Mean±Std.D(%)芽长ShootLength平均值Mean±Std.D(cm)根长RootLength平均值Mean±Std.D(cm)0(CK)10000±000Cd690±047Cc753±105Cd056667±577Bc230±020Bb390±014Bc103889±839Ab060±008Aa145±037Ab152556±962Aa045±006Aa035±006Aa
注:图中不同小写字母表示差异显著(P< 0.05),不同大写字母表示差异极显著(P< 0.01)
Note: Letters in capital and lower case indicate extremely significant difference level (P<0.01) and significant difference level (P<0.05) respectively

图2 核桃壳浸泡液对小麦种子发芽率芽长及根长的抑制作用
Fig.2 Inhibitory effect of walnut endocarp extracts on germination percentage, shoot and root length of wheat seeds
3 讨 论
种子休眠是自然界普遍存在的现象,而不同种(品种)种子的休眠程度和原因有所不同[5],同一种(品种)种子的休眠可能由多种因素控制[17-19]。低温层积处理打破休眠的效应在不同种类核桃之间有很大差异,这取决于种子的休眠类型和休眠深度[20]。 Vahdati等[21]提出6~8周低温处理最适合打破核桃种子休眠并克服由于休眠引起的生理矮生。研究表明,核桃干种子直接湿沙低温(4℃)层积处理21 d(T1)或浸水24 h再进行湿沙低温(4℃)层积处理21 d(T2)均对核桃种子的发芽有一定的促进作用,但T1和T2种子发芽率与其CK之间没有显著差异(|U| ≤ 1.96)。结果表明,层积处理21 d不足以满足有效打破普通核桃种子休眠所需。实验结果与Vahdati等[21]的结果一致,在层积处理过程中任一涉及调控种子休眠的因素都有可能发生变化。
植物内源激素参与休眠的调控,其中脱落酸(ABA)和赤霉素(GA3以及细胞分裂素(CTK)是涉及调控植物休眠的激素[22]。种子在低温层积处理过程中,随处理时间的延长其ABA浓度逐次下降[22-24],而内源GA3的浓度逐渐上升[25-26],使得GA3/ABA比值上升[24,27,28]。因此被认为,低温层积过程中改变GA3/ABA的比值是解除种子休眠的主要原因。黑核桃种子休眠的研究结果显示,黑核桃处于休眠的种子GA3含量较低,ABA含量最高,随层积时间延长GA3明显上升,ABA含量急剧下降到零,似乎GA3/ABA比值是调控黑核桃种子休眠和萌发的内部因子[29]。通过GA3对模式植物拟南芥(Arabidopsis)萌发的作用机制研究中发现,GA3处理可促进胚根的扩展并有利于种胚细胞壁的松弛和水解[30,31],这可能是用外源赤霉素改变体内GA3/ABA比值可解除休眠的原因之一。但外源赤霉素有效的发挥打破休眠的作用是以细胞分裂素的存在或萌发抑制物的不存在为条件,如果存在萌发抑制物并缺少细胞分裂素,外源赤霉素就不能有效发挥解除休眠的作用[32]。研究中,普通核桃种子在480 ppm的赤霉素(GA3)溶液中浸泡处理48 h(T5),可缩短发芽时间并有提高种子发芽率的趋势,但处理与CK之间没有显著差异(|U| ≤ 1.96)。结果表明,核桃种子休眠由GA3/ABA比值参与调控外,还有萌发抑制物质的影响。
实验结果显示,核桃种子浸水15 d处理(T3)或浸种15 d再暴晒阳光2 h(T4)均可提高核桃种子发芽率,与CK存在极显著差异(|U| ≥2.58);T3和T4播种第50 d的发芽率分别比CK高67.6%和104%;发芽势分别比CK高39.58%、190%;发芽指数分别比CK高100%、307%。对萌发抑制物质的存在或种皮的原因处于休眠的种子,播种前进行浸水处理洗脱水溶性萌发抑制物质,同时软化种壳、增强通透性,并能减小核桃种子硬壳对种胚生长的机械障碍可有效解除种子休眠[14,33]。阿卜杜许库尔等[14]研究结果表明,阜康阿魏种子含有水溶性的萌发抑制物质,进行20 d流水浸泡处理可去除该抑制物质,萌发率高达85%以上。荆条种子的萌发受到种皮和内部抑制物质的共同阻碍与抑制作用[34],黄香柏种皮含有萌发抑制物质[10],用清水漂洗都可有效地去除该抑制物质而解除休眠。近年来核桃种子休眠研究结果显示,不同种核桃种子休眠可能有差异。贾彩霞等[35]研究结果显示,核桃壳抑制物质中存在易溶于水的成分,也有易溶于乙醚的成分。曲芬霞等[36]研究认为,西藏核桃的种壳并不是引起休眠的主要原因,外壳、种仁可能均含有抑制种子发芽的物质。湖南山核桃种子休眠研究却显示种子没有休眠特性[37]。王沙沙等[38]的研究显示,6-BA可代替低温打破铁核桃种子休眠,浸种时间对铁核桃种子萌发均有显著影响。上述核桃种子休眠研究中,除湖南山核桃种子没有显示休眠特性之外,其它都涉及水溶性萌发抑制物质,这和该实验中T3的结果一致,据此确定普通种子的休眠主要受水溶性萌发抑制物质的调控。
为进一步证明水溶性萌发抑制物质在核桃种子休眠中的作用,实验用核桃壳浸提液萌发小麦种子结果显示,不同浓度的浸泡液对小麦的发芽率、芽长及根长随浓度的增加抑制作用逐渐增强,并在各浓度之间有极显著的差异,这一结果与阿卜杜许库尔等[14]研究结果一致。可见,核桃种壳中含有水溶性萌发抑制物存在,种子进行浸泡处理洗脱萌发抑制物质能使种子萌发。显然,水溶性萌发抑制物质在调控种子休眠中起主要作用。
将T3和T4之间相比,T4的发芽率、发芽势和发芽指数均比T3明显提高,并且有显著的统计学意义。这是浸水加阳光暴晒处理使核桃种子缝合线松弛,进一步削弱种壳对胚根突破种皮的机械阻力的结果。据此,确定核桃种子的硬壳是抑制种子萌发的另一个重要因素。Vahdati 等[21]用去壳和未去壳核桃种子进行低温层积处理,并对比研究其发芽率和发芽指数等生理指标也发现,硬壳阻碍引起核桃种子的机械休眠。
4 结 论
4.1 低于三周的短期层积处理不能满足核桃种子解除休眠的需要;
4.2 核桃种子解除休眠过程中虽有內源GA3/ABA比值的变化,但不是主要调控因素;
4.3 水溶性萌发抑制物质和硬壳的机械障碍是操纵核桃种子休眠的重要因素。
4.4 根据研究结果确定,核桃种子休眠属于由內源活性物质和抑制物质所调控的生理休眠型和受种壳调控的机械休眠型,也可称之为综合休眠型。
4.5 核桃种子播种前无需进行6周以上长期低温层积处理,而只有15 d浸水(每天需要更换清水)加2 h日光暴晒处理即可达到解除休眠培育良苗的目的。
)
[1] 郗荣庭,张毅萍.中国果树志.核桃卷[M].北京:中国林业出版社,1996.
XI Rong-ting,ZHANG Yi-ping.(1996).ChinaFruitTrees.WalnutVolume[M].Beijing: China Forestry Publishing House.(in Chinese)
[2]Manning, W. E. (1978). The classification within the juglandaceae.AnnalsoftheMissouriBotanicalGarden, 65(4): 1,058-1,087.
[3]吴国良,刘群龙,郑先波,等.核桃种质资源研究进展[J].果树学报,2009,26(4):539-545.
WU Guo-liang,LIU Qun-long,ZHENG Xian-bo,et al.(2009).Advances in research on the worldwide walnut germplasm [J].JournalofFruitScience,26(4): 539-545.(in Chinese)
[4]王磊,催乃然,张汉斐.新疆野核桃的研究[J].干旱区研究,1997,14(1):17-27.
WANG Lei,CUI Nai-ran,ZHANG Han-fei.(1997).Research report of Xinjiang wild walnuts [J].AridZoneResearch,14(1):17-27.(in Chinese)
[5]侯冬花,萨拉木·艾尼瓦尔,海利力·库尔班.种子休眠与休眠解除的研究进展[J].新疆农业科学,2007,44(3):349-354.
HOU Dong-hua,Salam Aniwar,Halil Kurban.(2007).Studies on seed dormancy and dormancy breaking [J].XinjiangAgriculturalSciences,44(3): 349-354.(in Chinese)
[6]赵鑫,马小军,凯撒·苏来曼,等.低温解除阜康阿魏种子休眠和内源激素变化规律的研究[J].中草药,2006,37(2):268-270.
ZHAO Xin,MA Xiao-jun,Kaisa Sulaiman,et al.(2006).Rule of breaking Ferulafukanensisseed dormancy under low temperature and content changes of endogenous hormone [J].ChineseTraditionalandHerbalDrugs,37(2): 268-270.(in Chinese)
[7]宁伟,范文丽,李宏博,等.变温及GA3处理对辽东木种子解除休眠过程中代谢调控的影响[J].园艺学报,2006,33(3): 649-652.
NING Wei,FAN Wen-li,LI Hong-bo,et al.(2006).Effect of alternating temperature and gibberellin (GA3) on metabolic control of Japanese Aralia (Araliaelata) Seed in dormancy breaking process [J].ActaHorticulturaeSinica,33(3):649-652.(in Chinese)
[8]阿卜杜许库尔·牙合甫,玉山江·依布拉音,海利力·库尔班.阜康阿魏种子打破休眠及其人工种植研究[J].新疆农业科学,2008,45(3):494-497.
ABuduxukuer Yahapu,Yushanjiang Yibulayin,Hailili Kuerban.(2008).A study on breaking dormancy ofFerulafukanensisShen. and its artificial cultivation [J].XinjiangAgriculturalSciences,45(3): 494-497.(in Chinese)
[9]Turner, S., & Merritt, D. (2009). Seed germination and dormancy. Plant Cell, 9(7): 1,055-1,066.
[10]Schmitz, N., Abrams, S. R., & Kermode, A. R. (2002). Changes in aba turnover and sensitivity that accompany dormancy termination of yellow-cedar (chamaecyparis nootkatensis) seeds.JournalofExperimentalBotany, 53(366): 89-101.
[11]Le, P. D. M., & Garello, G. (1992). In situ abscisic acid synthesis : a requirement for induction of embryo dormancy in helianthus annuus.PlantPhysiology, 98(4): 1,386-1,390.
[12]王蕾,海利力·库尔班,萨拉木·艾尼瓦尔.野生杏种子对外源赤霉素的生理响应[J].干旱区研究,2009,(5):708-713.
WANG Lei,Hailili Kurban,Salam Ainiwar.(2009).Physiological response of wild apricot seeds to exogenous gibberellins [J].AridZoneResearch,(5): 708-713.(in Chinese)
[13]鱼小军,王芳,龙瑞军.破除种子休眠方法研究进展[J].种子,2005,24(7):46-49.
YU Xiao-jun,WANG Fang,LONG Rui-jun.(2005).Studies on breaking seed dormancy [J].Seed,24(7):46-49.(in Chinese)
[14]阿卜杜许库尔·牙合甫,海利力·库尔班.生物碱参与阿魏种子休眠的调控[J].干旱区研究,2013,30(5):882-886.
Abduxukur Yakup,Hailili Kurban.(2013).Regulation of Alkaloid inFerulaSeeds Dormancy [J].AridZoneResearch,30(5):882-886.(in Chinese)
[15]郑光华.种子生理研究[M].北京:科学出版社,2004:623-661.
ZHENG Guang-hua.(2004).SeedPhysiologyResearch[M].Beijing: Science Press: 623-661.(in Chinese)
[16]杜荣骞.生物统计学[M].北京:高等教育出版社,2003.
DU Rong-qian.(2003).Biostatistics[M].Bei Jing:Higher Education Press.(in Chinese)
[17]Copete, E., Herranz, J. M., Ferrandis, P., Baskin, C. C., & Baskin, J. M. (2011). Physiology, morphology and phenology of seed dormancy break and germination in the endemic iberian species narcissus hispanicus (amaryllidaceae).AnnalsofBotany, 107(6): 1,003-1,016.
[18]杨期和,尹小娟,叶万辉.硬实种子休眠的机制和解除方法[J].植物学通报,2006,23(1):108-118.
YANG Qi-he,YIN Xiao-juan,YE Wan-hui.(2006).Dormancy mechanism and breaking methods for hard seeds [J].ChineseBulletinofBotany,23(1): 108-118.(in Chinese)
[19]Ross, J. D. (1995). Seeds: physiology of development and germination.Phytochemistry, 38(6): 1,551.
[20]Chandler, W.H., Kimball, M.H., Phillip, G.L., Tufts, W.P., Weldon, G.P., (1937).Chilling requirements for opening of buds on deciduous orchard trees and some other plants in California. Calif. Agric. Exp. Bull. 611.
[21]Vahdati, K., Aslamarz, A. A., Rahemi, M., Hassani, D., & Leslie, C. (2012). Mechanism of seed dormancy and its relationship to bud dormancy in persian walnut.Environmental&ExperimentalBotany,75(4): 74-82.
[22]Powell, L. E. (1987). Hormonal aspects of bud and seed dormancy in temperate-zone woody plants.Hortscience, 22(5): 845-850.
[23]Rodríguez, A., Canal, M. J., & Sánchez-Tamés, R. (1991). Seasonal changes of plant growth regulators in corylus.JournalofPlantPhysiology, 138(1): 29-32.
[24]Hilhorst, H. W. M., & Karssen, C. M. (1992). Seed dormancy and germination: the role of abscisic acid and gibberellins and the importance of hormone mutants.PlantGrowthRegulation, 11(3): 225-238.
[25]Hoad, G. V. (1992). Gibberellins. Edited by Takahashin. Phinneyb. o. and Macmillanj. Berlin: Springer-Verlag, (1991), pp. 426, dm178. ISBN 3-540-97259-5.ExperimentalAgriculture, 28(1): 426-129.
[26]Wiebel, J., Downton, W. J. S., & Chacko, E. K. (1992). Influence of applied plant growth regulators on bud dormancy and growth of mangosteen (garcinia mangostana, l.).ScientiaHorticulturae, 52(1-2): 27-35.
[27]罗弦,潘远智,杨学军,等.低温层积处理对4种苔草种子休眠与萌发的影响[J].草业学报,2010,19(3):117-123.
LUO Xian,PAN Yuan-zhi,YANG Xue-jun,et al.(2010).Effect of cold stratification on dormancy and germination of four Carex species seeds [J].ActaPrataculturaeSinica, 19(3):117-123.(in Chinese)
[28]Wang, W. Q., Song, S. Q., Li, S. H., Gan, Y. Y., Wu, J. H., & Cheng, H. Y. (2009). Quantitative description of the effect of stratification on dormancy release of grape seeds in response to various temperatures and water contents.JournalofExperimentalBotany, 60(12): 3,397-3,406.
[29]裴东,张俊佩,石永森,等.层积催芽对美国黑核桃种子发芽和苗木生长的影响[J].林业科学,2002,38(5):73-77.
PEI Dong,ZHANG Jun-pei,SHI Yong-sen,et al.(2002).Seed germination and seedling growth associated with stratification ways onJuglansNigra[J].ScientiaSilvaeSinicae,38(5):73-77.(in Chines)
[30]Puype, M., Demol, H., Vandekerckhove, J., & Job, D. (2002). Proteomics of arabidopsis seed germination. a comparative study of wild-type and gibberellin-deficient seeds.PlantPhysiology, 129(2): 823-837.
[31]Groot, S. P. C., & Karssen, C. M. (1987). Gibberellins regulate seed germination in tomato by endosperm weakening: a study with gibberellin-deficient mutants.Planta, 171(4): 525-531.
[32]李宏博,刘延吉,李天来.GA3对辽东木楤种子解除体眠中信号分子变化的影响[J].园艺学报,2006,33(2):410-414.
LI Hong-bo,LIU Yan-ji,LI Tian-lai.(2006).The effect of GA3on the relative signal molecule changes during the breaking dormancy in Aralia elata seed [J].ActaHorticulturaeSinica,33(2):410-416.(in Chinese)
[33]罗旭辉,詹杰,陈义萍,等.浸种处理对闽引羽叶决明种子萌发的影响[J].草原与草坪,2010,30(2):47-49,55.
LUO Xu-hui,ZHAN Jie,CHEN Yi-ping,et al.(2010).Effect of soaking treatments on germination ofChamaecristarotundifoliacv. Minyin seeds [J].GrasslandandTurf,30(2):47-49,55.(in Chinese)
[34]李义强,宋桂龙,郭宇.水浸与赤霉素处理对荆条种子萌发影响研究[J].种子,2012,31(3):10-13.
LI Yi-qiang,SONG Gui-long,GUO Yu.(2012).Study on effect of water immersion and gibberellin treatments on seed germination ofVitexnegundovar.heterophylla[J].Seed,31(3):10-13.(in Chinese)
[35]贾彩霞,翟梅枝,宋艳枝.核桃壳内源抑制物质活性的初步研究[J].西北林学院学报,2006,21(3):98-100.
JIA Cai-xia,ZHAI Mei-zhi,SONG Yan-zhi.(2006).A Preliminary study on the intrinsic inhibitor activities from walnut endocarp [J].JournalofNorthwestForestryUniversity,21(3):98-100.(in Chinese)
[36]曲芬霞,姚霞珍,郭建斌,等.西藏核桃休眠及萌芽特性研究[J].种子,2011,30(3):44-47.
QU Feng-xia,YAO Xia-zhen,GUO Jian-bin,et al.(2011).Study on seed dormancy and germination characteristic ofJuglansregiain Tibet [J],Seed,30(3):44-47.(in Chinese)
[37]樊卫国,马文涛,龙登楷,等.保湿冷藏对湖南山核桃种子萌发及生理的影响[J].果树学报,2011,28(5):831-836.
FAN Wei-guo,MA Wen-tao,LONG Deng-kai,et al.(2011).Effects of moisture-preserved refrigeration and natural dehydration on the seed germination and physiology ofCaryahunanensis[J].JournalofFruitScience,28(5):831-836.(in Chinese)
[38]王沙沙,潘学军,张文娥,等.铁核桃Juglanssigillata种子萌发的影响因素研究[J].西南师范大学学报(自然科学版),2013,38(11):70-75.
WANG Sha-sha,PAN Xue-jun,ZHANG Wen-e,et al.(2013).On Influential Factors of Seed Germination ofJuglansSigillata[J].JournalofSouthwestChinaNormalUniversity(NaturalScienceEd.) ,38(11):70-75.(in Chinese)
