花铃期干旱胁迫对不同棉花品种光合作用影响
2018-01-22郑巨云王俊铎龚照龙梁亚军艾先涛郭江平段静波李雪源
郑巨云,王俊铎,龚照龙,梁亚军,艾先涛,郭江平, 段静波,莫 明,李雪源
(1.新疆农业科学院经济作物研究所,乌鲁木齐 830091;2.新疆农业大学科学技术学院,乌鲁木齐 830052)
0 引 言
【研究意义】新疆棉区属于中温带至暖温带干旱气候区,棉区大都分布在河流两岸冲积平原和三角洲地带,棉花种植对灌溉的依赖性很强,本区河流水资源尤为匮乏,而且季节分配不均,春、秋、冬季缺水,夏季时有洪水,严重影响了棉花生产,加之林果业、工业的发展及生态用水的保障,造成新疆棉花生产用水更加紧张。新疆已成为我国最大、最有竞争力的棉花生产基地,干旱已成为制约新疆棉花生产持续发展的关键因子,提高棉花耐旱基础研究和应用研究面临的主要任务。干旱是限制棉花生长发育和产量的主要环境因子之一,棉花的营养生长、生殖生长所需的有机物主要来自于叶片的光合作用,因此,通过研究不同品种在抗旱胁迫及正常灌水条件下的光合性状差异,对明确干旱胁迫对棉花光合性状的影响,筛选抗旱能力强的品种及抗旱育种的遗传改良具有重要意义。【前人研究进展】不同棉花品种之间的叶绿素含量、光合速率存在较大差异研究棉花抗旱特性,可节约水源,提高棉花产量。光合作用是植物将太阳能转换为化学能的过程,既是植物体内最重要的化学反应[1]又是植物生长发育的基础和植物体内碳素的重要来源[2]也是干物质积累的基础[3]和决定生产力高低的重要因素[4]花铃期是棉花(GossypiumhirsutumL.)对肥水需求最敏感的时期,亦是最终产量与品质形成的关键时期[5]。随着我国棉花生产区域布局的调整,花铃期季节性干旱已成为棉花生产上主要的自然灾害,严重影响棉花生长,使产量和品质大幅度降低[6-7]。盛蕾期与铃期是棉花肥水最重要的时期,所以主要对盛蕾期与花期在干旱胁迫下光合特性进行研究,应加强研究棉花盛蕾期与花期水分胁迫下光合特性。棉花具有喜光喜温的特点,但超过生长发育适宜的温度,光合作用会受到影响,花铃中期水分亏缺导致群体光合速率比适水下降42.2%,皮棉产量减少741 kg/hm2。【本研究切入点】设立干旱胁迫与正常大田两个水平对棉花光合特性差异进行研究,可为抗旱种质资源鉴定和抗旱育种提供理论依据和技术参考。在干旱胁迫条件下和正常灌水条件下,测定不同品种盛花期、盛铃期光合生理性状各指标,对比分析不同条件下光合性状各指标的差异及相关性。【拟解决的关键问题】研究比较分析光合生理指标的绝对值及抗旱系数,明确干旱胁迫对光合生理指标的影响,筛选抗旱能力强的品种材料。
1 材料与方法
1.1 材 料
选用16N2、16N3、16N4、16N5、源棉6号为材料。这5个品种是经过多年多点筛选出来产量性状突出,品质性状好、抗性好,在生产上示范推广应用的品种材料,具有不同耐旱性的棉花材料。试验在新疆农业科学院第一师16团试验站试验田,三重复随机排列,株系行长8 m,株距11.5 cm,机采模式(10+66+10+66+10) cm,一膜6行种植,种植密度17 950株/667 m2,田间管理同大田生产管理。
1.2 方 法
1.2.1 试验设计
2016年4月在新疆农业科学院阿拉尔16团基地旱池种植,并进行抗旱处理。干旱胁迫条件(W1)浇出苗水,全生育不浇水;正常灌水条件(W2)按大田管理正常灌水,6月10日第一次滴水,花铃期每7~10 d滴1次水,全生育期滴水总量为350 m3。
1.2.2 光合生理指标测定
分别在盛花期和盛铃期,利用LiCpro+便携式手动光合测定仪测定各材料的光合性状相关指标。采用25 mm×25 mm叶室,于晴朗无云天气测定,测定时间为10:30~12:30,打顶前测定主茎倒4叶,打顶后测定倒2叶[3]。每个株系随机选定连续5株,3重复,测试其光合性状基本参数:叶面温度(Tl,℃)、水蒸气的气孔导度(Gs,mol/(m2·s))、单叶净光合速率(Pn,μmol/(m2·s))、蒸腾速率(Tr,μmol/(m2·s))。通过光合速率(Pn)、蒸腾速率(Tr),计算出光合水分利用率(光合水分利用率PWUE=Pn/Tr)。
1.2.3 表型性状
盛花期及盛铃期光合性状的抗旱系数计算参照Blum[8]、冯方剑[9]、郑巨云[10]等方法。计算公式:CD=Ti,S/Ti,W. 式中,Ti,S为某一性状(i)在水分胁迫(S)环境下的测定值,Ti,W为为某一性状(i)在正常灌水(W)环境下的测定值。
1.3 数据处理
数据分析采用EXCLE2010软件和SAS9.0软件对田间测试数据进行统计分析。
2 结果与分析
2.1 亲本及处理之间差异
研究表明,盛花期各品种之间叶片温度、气孔导度、蒸腾速率、P均小于0.05表现差异显著,只有光合速率、光合水分利用率表现差异不显著。重复之间表现差异均不显著,说明重复之间误差在合理试验范围之内,适于进一步分析。干旱与正常灌水条件不同处理之间差异除光合速率P>0.05差异不显著,叶片温度、气孔导度、蒸腾速率、光合水分利用率P<0.05,达到显著差异水平。品种与处理间互作结果为,叶片温度、光合速率P>0.05,表现差异不显著,气孔导度、蒸腾速率、光合水分利用率P<0.05,表现差异显著。表1
研究表明,盛铃期光特性分析中,不同品种之间叶片温度、蒸腾速率、光合速率P>0.05表现差异不显著,气孔导度、光合速率、光合水分利用率P<0.01差异极显著。重复之间除叶片温度P<0.05差异显著,气孔导度、光合速率、蒸腾速率及光合水分利用率均P>0.05差异不显著。不同处理之间蒸腾速率及光合水分利用率P<0.05差异显著,叶片温度、气孔导度及光合速率P>0.05差异不显著。品种与处理之间互作,叶片温度、蒸腾速率P>0.05差异不显著,气孔导度、光合速率及光合水分利用率P<0.05差异显著。表2
表1 盛花期光合性状双因素方差
Table 1 Double-factor variance analysis on photosynthetic traits at the flowering stage

性状TraitP品种PvaluesforvarietiesP重复PvaluesforrepeatsP处理PvaluesfordifferentenvironmentsP品种与处理互作Pvaluesforgenotype-environmentinteractionofvarieties叶片温度 Tl000320374200001006气孔导度 Gs0001905634<00001<00001光合速率 Pn0140609703011607185蒸腾速率 Tr000406798<00001<00001光合水分利用率 PWUE00732072280004500207
注:P为0.05水平双因素方差分析概率值,P≤0.05表示差异显著,P>0.05表示差异不显著
Note:Pare probability values of two-factor variance analysis;P≤0.05 indicates significant difference, orP>0.05 indicates no significant difference
表2 盛铃期光合性状双因素方差
Table 2 Double-factor variance analysis on photosynthetic traits at the full-boll stage

性状TraitP品种PvaluesforvarietiesP重复PvaluesforrepeatsP处理PvaluesfordifferentenvironmentsP品种与处理互作Pvaluesforgenotype-environmentinteractionofvarieties叶片温度 Tl02512000860050408277气孔导度 Gs00008050940261300006光合速率 Pn0006016730285900225蒸腾速率 Tr0166201645<0000104861光合水分利用率 WUSE<0000103219<00001<00001
注:P为0.05水平双因素方差分析概率值,P≤0.05表示差异显著,P>0.05表示差异不显著
Note:Pare probability values of two-factor variance analysis;P≤0.05 indicates significant difference, orP>0.05 indicates no significant difference
2.2 干旱胁迫条件下盛花期光合性状指标
研究表明,盛花期抗旱条件下光合特性各项指标值是不同的。在盛花期叶片温度16N4、16N5、源棉6号较高是处于相同水平,其次是16N3号,16N2号最低,叶片温度最低,蒸腾速率16N2、16N3、16N4、源棉6号较高处于同一水平,在盛花期升腾速率较快,16N5蒸腾速率较其他四个品种相对较低。气孔导度16N2最高16N3比16N2较低,16N4与源棉6号处同一水平,比16N5高较16N2、16N3较低。光合速率16N3最高,16N2较16N3稍低,其他三个品种处同一水平,水分利用率16N5最大,其他四个品种处同一水平比16N5较低。表3
表3 盛花期抗旱条件下各性状指标

Table 3 Photosynthetic character indexes of different variety under W1 at flowering stage
注:表中数字后的小写英文字母表示5%显著水平
Note: The different lowercase letters mean significant differences at 5%
研究表明,盛花期棉花光合性状抗旱系数中叶片温度CD值16N4最高,其次16N3和16N5处同一水平,16N2最低。蒸腾速率CD值16N3号最高,16N2、16N4、源棉6号较低处相同水平,16N5最低。气孔导度CD值16N3最高,16N2、源棉6号、16N4、16N5从高到低逐渐降低。光合速率CD值在盛花期都处于相同水平较高。水分利用率CD值16N5最高,其他四个品种处于相同水平相对较低。表4
表4 盛花期抗旱系数
Table 4 Drought resistance coefficient of photosynthetic traits at flowering stage
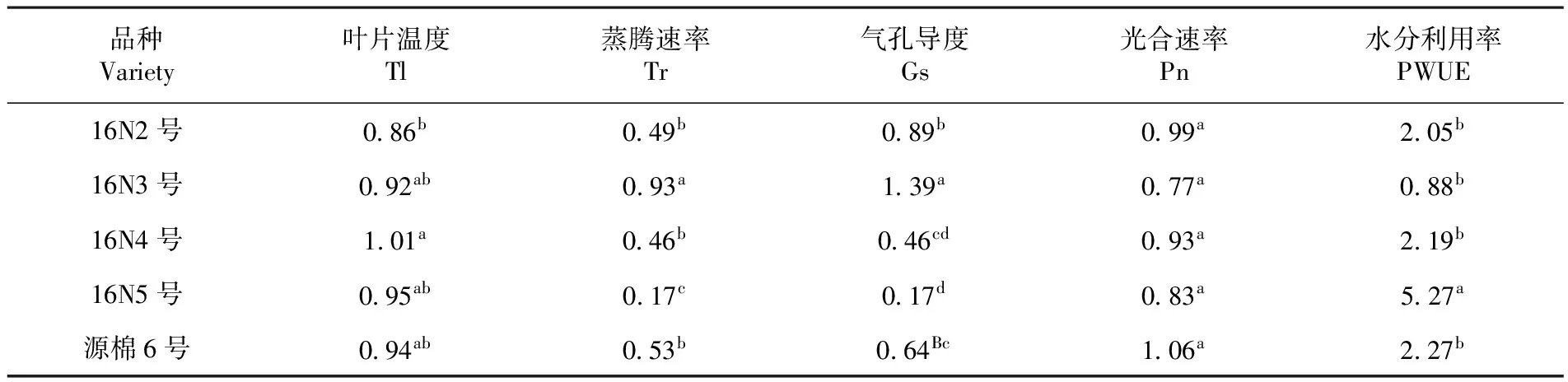
品种Variety叶片温度Tl蒸腾速率Tr气孔导度Gs光合速率Pn水分利用率PWUE16N2号086b049b089b099a205b16N3号092ab093a139a077a088b16N4号101a046b046cd093a219b16N5号095ab017c017d083a527a源棉6号094ab053b064Bc106a227b
注:表中数字后的小写英文字母表示5%显著水平
Note: The different lowercase letters mean significant differences at 5%
2.3 干旱胁迫条件下盛铃期光合性状指标及抗旱系数
研究表明,盛铃期抗旱干旱胁迫条件下各项光合性状指标存在不同差异,叶片温度,16N5、源棉6号最高,16N4、16N3处同一水平较低,16N2最低;蒸腾速率,16N5最高,16N2、16N3最低,16N4和源棉6号处同一水平位于最高与最低之间;气孔导度,16N2最高,其他四个品种较低且处同一水平;光合速率16N2最高,其他四个品种较低且处同一水平;水分利用率16N2最高,其他四个品种较低且处同一水平。表5
表5 盛铃期抗旱条件下各性状指标

Table 5 Photosynthetic character indexes of different variety under W1 at full-boll stage
注:表中数字后的小写英文字母表示5%显著水平
Note: The different lowercase letters mean significant differences at 5%
研究表明,5个品种叶片温度CD值处同一水平;5个品种的蒸腾速率CD值也出同一水平,差异较小,气孔导度CD值16N2最高,其他较低处同一水平。光合速率CD值16N2最高,其他四个品种较低处同一水平。水分利用率CD值16N2最高,源棉6号较低,16N3、16N4、16N5、最低处同一水平。表6
表6 盛铃期抗旱系数
Table 6 Drought resistance coefficient of photosynthetic traits at full-boll stage

品种Variety叶片温度Tl蒸腾速率Tr气孔导度Gs光合速率Pn水分利用率PWUE16N2号102a018a513a154a866a16N3号103a017a036b063b373b16N4号104a026a052b072b297b16N5号106a027a071b082b338b源棉6号102a020a048b095b551ab
注:表中数字后的小写英文字母表示5%显著水平
Note: The different lowercase letters mean significant differences at 5%
3 讨 论
干旱适应性是植物对干旱胁迫的一种生理响应机制,而不同的植物对胁迫的敏感度不同,形成不同的干旱适应性机制。花铃期是棉花肥水最重要的时期,所以主要对花铃期干旱胁迫下和正常灌水条件下的光合生理特性进行研究。叶面温度是在进行光合测定时棉花叶片的表面温度,叶面温度越高则说明光合作用越明显[11]。研究发现盛花期叶片温度抗旱系数除16N4外,其他均小于1,说明盛花期干旱胁迫对叶片温度影响较大,叶片温度降低造成光合作用能力下降,而盛铃期叶片温度抗旱系数约为1,说明盛铃期干旱胁迫对叶片温度影响较小,基本可以忽略。
气孔导度是植物与外界环境进行物质交换的主要通道气孔通过调节其开闭状态,控制着CO2进入植物体内,以及水分从体内的散失[12]。研究发现除16N3盛花期气孔导度抗旱系数和16N2盛铃期气孔导度抗旱系数大于1以外,其它材料花铃期气孔导度抗旱系数均小于1,这说明干旱胁迫会降低气孔导度,减少了水分的散射和CO2进入,从而降低棉花光合速率。
植物蒸腾是土壤到植物到大气连续体水热传输过程中一个极为重要的环节,是植物和环境之间物质交换和能量平衡的重要生理过程[13],研究发现5份材料盛花期、盛铃期蒸腾速率抗旱系数均小于1,说明干旱胁迫会明显降低植物叶片的蒸腾速率,减少水分散失,而不同品种降低的幅度不同。
作物水分利用率指消耗单位水分所生产的同化物质的量它实质上反映了作物耗水与其干物质生产之间的关系,是评价作物生长适宜程度的综合生理生态指标[14]。研究发现5份材料盛花、铃期水分利用率抗旱系数均大于1,说明干旱胁迫会明显提高棉花叶片的水分利用效率,在水分匮乏条件下,通过提高水分利用率,保持棉花叶片的一定的光合作用,但不同品种的水分利用率抗旱系数差异显著。
前人研究发现陆地棉或海岛棉种内品种(系)之间的光合速率也存在一定差异[15],研究发现除源棉6号盛花期单叶净光合率抗旱系数和16N2盛铃期单叶净光合率以外,其它材料盛花期、盛铃期单叶净光合速率抗旱系数均小于1,说明干旱胁迫会显著降低棉花叶片的单叶净光合速率,但不同品种的单叶净光合速率抗旱系数存在显著差异,16N2盛花期单叶净光合速率抗旱系数约为1,盛铃期单叶净光合速率抗旱系数大于1,说明干旱胁迫对16N2盛花期单叶净光合速率负影响较小,持续胁迫会提高16N2盛铃期单叶净光合速率。
戴茂华等[16]研究发现棉花在不同生育时期对水分的需求量不同,抵抗干旱胁迫的内在机制也有所不同。研究发现在花铃期干旱胁迫下,抗旱性强的品种单叶净光合速率降幅小,水分利用率较高,能够保持较高水分利用率,维持较高的光合作用以产生更多的光合有机物,维持棉花正常生长发育,如干旱胁迫下16N2单叶光合速率于盛花期至盛铃期逐渐上升,保持较高的水分利用率,维持基本的光合生理功能,表现出良好的抗旱能力。
4 结 论
花铃期干旱胁迫处理会降低棉花叶片蒸腾速率、气孔导度及光合速率等光合生理指标,但可以提高光合水分利用率,不同品种光合性状指标差异显著。干旱条件下盛花期16N3单叶净光合速率最高为10.31 μmol/(m2·s),16N2、源棉6号其次均为8.93 μmol/(m2·s),其它2份材料单叶净光合速率均小于8.0 μmol/(m2·s),盛花期源棉6号、16N2单叶净光合速率抗旱系数最高,分别为1.06、0.99。盛铃期16N2的单叶净光合速率、气孔导度、光合水分利用率最高分别为0.2 mmol/(m2·s)、14.75 μmol/(m2·s)、48.11 g/kg,源棉6号单叶净光合速率、光合水分利用率次之,分别为8.16 μmol/(m2·s)、21.40 g/kg,其它三个品种较低。花铃期干旱胁迫环境下,16N2光合性状指标抗旱系数最高,叶片温度CD值、气孔导度CD值、单叶净光合速率CD值、光合水分利用率CD值分别为1.02、5.13、1.54、8.66,具有较强的抗旱能力,可以作为抗旱育种的中间材料,为抗旱聚合育种奠定基础。
)
[1] 佘诚棋,程鹏,季琳琳,等. 油茶光合作用光响应曲线的拟合[J].经济林研究,2012,(1):118-120,137.
SHE Chen-qi, CHENG Peng, JI Lin-lin, et al. (2012). Light-response curves fi tting of photosynthesis inCamelliaoleifera[J].NonwoodForestResearch,(1):118-120,137. (in Chinese)
[2]韦铄星,文仕知,张希,等. 不同林龄四川桤木的光合生理特性分析[J]. 中南林业科技大学学报,2011,(7):102-108.
WEI Shuo-xing, WEN Shi-zhi, ZHANG Xi, et al. (2011). Analysis on photosynthesis physiological characteristics of Alnus cremastogyne at different stand ages [J].JournalofCentralSouthUniversityofForestry&Technology, (7):102-108. (in Chinese)
[3]王怡.水分胁迫对3个油橄榄品种光合及生理特性的影响[D]. 雅安:四川农业大学硕士学位论文,2012.
WANG Yi. (2012).StudyonEffectsofWaterStressonPhotosynthesisandPhysiologicalCharacteristicsofThreeOliveVarieties[D]. Master Thesis. Sichuan Agricultural University, Ya'an. (in Chinese)
[4]高超,闫文德,田大伦,等.杜仲光合速率日变化及其与环境因子的关系[J].中南林业科技大学学报,2011,(5):100-104.
GAO Chao, YAN Wen-de, TIAN Da-lun, et al. (2011). Diuranal change in photosynthetic rate ofEucommiaulmoidesand its relationship with environmental factors [J].JournalofCentralSouthUniversityofForestry&Technology, (5):100-104. (in Chinese)
[5]余渝,陈冠文,田笑明,等.北疆高产棉花光合特性研究[J],棉花学报,2001, 13(4): 230-233.
YU Yu, CHEN Guan-wen, TIAN Xiao-ming, et al. (2001). Research on the Photosynthesis Characteristic of the High Yield Cotton in North Xinjiang [J].CottonScience,13(4): 230-233. (in Chinese)
[6]郭纪坤. 陆地棉抗旱耐盐及产量、形态性状的QTL定位[D]. 乌鲁木齐: 新疆农业大学硕士学位论文, 2007.
GUO Ji-kun. (2007).QTLsassociatedwithdroughttolerance,saltresistance,yieldandmorphologicaltraitsinuplandcotton[D]. Master Thesis. Xinjiang Agricultural University, Urumqi. (in Chinese)
[7]李少昆, 肖璐, 黄文华. 不同时期干旱胁迫对棉花生长和产量的影响Ⅱ.棉花生长发育及生理特性的变化[J]. 石河子大学学报(自然科学版), 1999, 3(4): 259-264.
LI Shao-kun, XIAO Lu, HUANG Wen-hua, et al. (1999). Effect of drought stress on cotton growth and lint yield at different growing stage II. The change of cotton growth and physiological characteristics to water stress [J].JournalofShiheziUniversity(NaturalScience) , 3(4): 259-264. (in Chinese)
[8] Abraham Blum, & Wayne R. Jordan. (1985). Breeding crop varieties for stress environments.CriticalReviewsinPlantSciences, 2(3): 199-238.
[9]冯方剑,宋敏,陈全家,等.棉花苗期抗旱相关指标的主成分分析及综合评价[J].新疆农业大学学报, 2011,34(3):211-217.
FENG Fang-jian, SONG Min, CHEN Quan-jia, et al. (2011). Analysis and Comprehensive Evaluation on Principal Component of Relative Indices of Drought Resistance at the Seedling Stage of Cotton [J].JournalofXinjiangAgriculturalUniversity, 34(3):211-217. (in Chinese)
[10]郑巨云,GeorgeOluoch,Khan Muhammad Kashif Riaz, 等.毛棉苗期抗旱性状的 QTL定位[J].棉花学报,2017,29(1):29-39.
ZHENG Ju-yun, George Oluoch,Khan Muhammad Kashif Riaz, et al. (2017). Mapping QTLs for Traits Related to Drought Tolerance at Seedling Stage in an Inter-Specific Gossypium hirsutum × G. tomentosum Cotton F2:3 Population [J].CottonScience, 29(1):29-39. (in Chinese)
[11]卫林,江爱良,蒋世逵.农田叶面温度计算方法的研究[J].中国农业气象,1981,(4):37-42.
WEI Lin, JIANG Ai-liang, JIANG Shi-kui. (1981). The study of leaf temperature of farmland. Agriculture Weather [J],ChineseJournalofAgrometeorology, (4):37-42. (in Chinese)
[12]钟楚,朱勇.烟草气孔导度对光强的响应[J].中国生态农业学报,2013,21(8):966-972.
ZHONG Chu, ZHU Yong. (2013). Response of stomatal conductance to light in tobacco plants [J].ChineseJournalofEco-Agriculture, 21(8): 966-972. (in Chinese)
[13]赵春彦,司建华,冯起,等.风对极端干旱区胡杨蒸腾速率的影响[J].冰川土,2015,(4):1 104-1 111.
ZHAO Chun-yan, SI Jian-hua,FENG Qi,et al.(2015). Effect of wind speed on transpiration rate ofPopuluseuphraticain extreme arid deserts [J].JournalofGlaciologyandGeocryology, 37(4):1,104-1,111.(in Chinese)
[14]江云,马友华,陈伟,等.作物水分利用率的影响因素及其提高途径探讨[J].中国农学通报,2007,(9):269-273.
JIANG Yun, MA You-hua, CHEN Wei, et al. (2007). Review on Influence Factor and Improvement Way of Crop Water Use Efficiency [J].ChineseAgriculturalScienceBulletin, (9):269-273. (in Chinese)
[15]梁哲军,赵海祯,齐宏立,等.基因型差异对棉花光合产物生产和分配的影响及遗传改良研究[J].棉花学报,2005,17(1):18-22.
LIANG Zhe-jun, ZHAO Hai-zhen, QI Hong-li, et al. (2005). Studies on Genetic Improvement and Effects on Production and Distribution of Photosynthate among Different Genotypes of Cotton [J].CottonScience, 17(1): 18-22. (in Chinese)
[16]戴茂华,岳秀琴,刘丽英,等.花铃期干旱胁迫对棉花光合特性的影响[J].安徽农业科学,2015,(19):4-5,20.
DAI Mao-hua, YUE Xiu-qin, LIU Li-ying,et al. (2015). Effects of Drought During Flowering and Boll-forming Stages on Photosynthetic Characteristics of Cotton [J].JournalofAnhuiAgri.Sci.,43(19): 4-5,20. (in Chinese)
