海拔对泰山侧柏林种子活力的影响
2018-01-17张梦颖王树梅曹帮华毛培利李业宇庞元湘范祖硕秦永建
张梦颖,王树梅,曹帮华,毛培利,李业宇,庞元湘,范祖硕,秦永建
(1.山东省高校森林培育重点实验室,山东泰山森林生态系统国家定位观测研究站,山东农业大学生态与环境重点实验室,山东 泰安271018;2.山东省林木种苗和花卉站,山东 济南250014)
植物群落的演替更新始终是森林生态系统研究的热点之一。天然更新是森林生态系统自我繁衍、恢复的手段和基础。种子能否萌发、产生正常苗是生态系统更新的关键阶段[1]。种子是物种繁殖系统的重要特征,在强大的选择压力下表现出很大的适应性[2],对植物种群的分布格局、种群动态及种群的调控等方面均有重要意义[3]。种子进入土壤后,发芽速度快和发芽率高对幼苗成功建立具有重要意义[4]。这与发芽时间越长,种子越容易被病菌感染导致不能发芽密切相关[5]。
海拔是影响植物种子活力的重要因子。海拔对光照、水分、热量等一系列环境因子有着综合影响,而不同环境变量之间的协同作用对植物生物量的积累和种子活力大小产生重要影响。有研究表明,不同海拔春小麦的种子活力差异显著,高海拔春小麦种子活力明显高于低海拔[6]。急尖长苞冷杉在海拔3 600~4 300 m之间发芽率先增加然后降低,其中3 900 m处发芽率最高[7]。对海拔1 500~1 700 m的独活种子进行质量和重量研究,1 600 m处的种子质量和产量最好,随后随海拔的增加质量降低[8]。说明树种对海拔有一定的最佳适应范围,但不同树种之间的适应范围存在差异。
侧柏(Platycladus orientalis)是泰山景区的主要组成树种之一,在泰山风景林资源中占有重要地位。经营管理好侧柏林对整个泰山风景维持、景区森林资源可持续发展和永续利用有着重要的意义。泰山景区的侧柏林主要是20世纪50年代营造的人工纯林。由于属于风景林,在管理过程中对林分的改造要求严格,极少进行抚育更新,造成林分结构不合理,严重影响了其景观效果。野外调查表明,侧柏具有较好的天然更新能力[9]。天然更新可以大大降低更新成本,同时又有利于保持其生态稳定性和环境效益的持续性。但是目前对于泰山侧柏种子活力与天然更新之间的关系研究并不深入。本试验以泰山不同海拔侧柏林内采集的种子为研究对象,通过光照培养箱观测了种子的发芽过程,分析了种子发芽能力,探讨海拔对侧柏种子活力的影响规律,为泰山侧柏林的管理提供理论支撑。
1 材料与方法
1.1 研究区概况
泰山海拔1 545 m,地形复杂,坡度较大。泰山年均气温10℃,年降水量700~900 mm,年日照总时数为2 893 h,年平均湿度达62%。母岩多为杂性,土壤为中性、微酸性棕色森林土,土层厚度为15~60 cm[10]。森林面积约3.35万hm2,森林覆盖率81.5%,泰山属暖温带气候区,基带植被类型为暖温带落叶阔叶林和温性针叶林。侧柏是泰山主要人工林之一,20世纪50年代营造,以其为主的林分总面积为1 080 hm2,主要分布在海拔800 m以下的山坡上。
1.2 种子采集
2015年9月,分别在海拔200 m(普照寺)和的海拔300 m(万仙楼)以及顺次沿泰山登山东路向上500,800,1 000 m的侧柏林内采集种子。在每个海拔侧柏林内设置10 m×10 m的临时样地3个,对高度>1.5 m的侧柏进行高度、胸径测量,林分具体特征见表1。在海拔1 000 m已经达到了侧柏分布的上线,向上很少有侧柏林,多为零星分布。每块样地选择5株接近平均木的母树,收集种子进行试验。即每个海拔选15株母树收集当年种子进行试验。将种子带回实验室后阴干贮藏至2016年4月,清除杂质,测定种子千粒重。
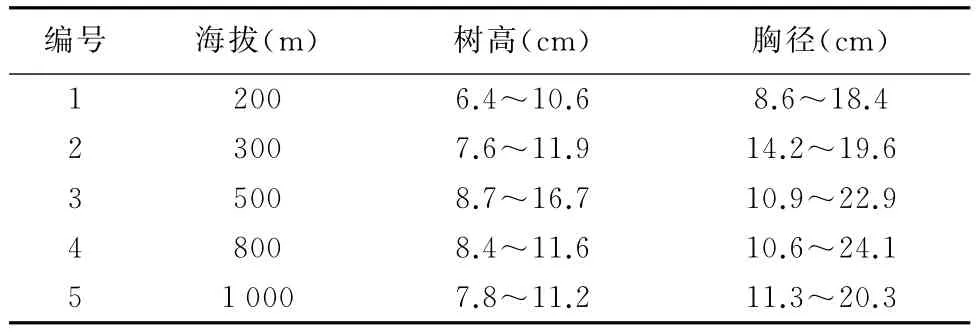
表1 不同海拔侧柏林主要特征
1.3 试验设计
1.3.1 种子萌发测定
2016年4月10日,在光照培养箱内用培养皿进行种子萌发。将种子用40℃温水浸种48 h,按数量要求随机挑选出无病虫害种子作为试验材料,用4‰高锰酸钾溶液对种子消毒30 min,然后用蒸馏水冲洗干净进行摆盘,用滤纸和纱布制作发芽床。每个培养皿摆50粒种子,3次重复。参照《林木种子检验规程》技术[11],将培养皿置于25℃培养箱中,每天对滤纸发芽的种子进行观察,及时补充水分并对发霉种子进行消毒,逐日记录发芽情况。第21天结束发芽实验。
1.3.2 测定指标及方法
1)种子千粒重测定。从不同海拔的纯净种子中随机选出100粒为1组,重复取8组称重,在变异系数不超过6.0的条件下,即可按测量结果计算千粒重。
2)发芽指标测定。发芽试验结束后分别计算种子的发芽率、发芽势、发芽指数、活力指数(VI)及平均发芽时间(MGT)。同时统计每个培养皿的萌发幼苗的重量计算单株幼苗的生物量。
发芽率(%)=发芽种子数/供试种子总数×100%;
发芽势(%)=规定日期内发芽种子数/供试种子总数×100%;
发芽指数(GI)=∑Gt/Dt(式中:Dt为发芽试验第几日;Gt为第几日的发芽数);
平均发芽时间(MGT)=(∑D×n)/∑n(式中:n为相应各日正常发芽粒数;D为从种子置床起的天数);
活力指数(VI)=S∑(Gt/Dt)=S×GI(式中:S为种苗生长量(鲜重,g);GI为发芽指数)。
3)根系活力指标。取第21天摆盘发芽的幼苗,用TTC法测幼苗根系活力。以四氮唑还原强度[mg/(g·h)]作为活力指标[12]。
1.3.3 数据分析
分别对不同海拔侧柏种子的千粒重、发芽率、发芽势、发芽指数、活力指数、简化活力指数、幼苗重量,利用SPSS20.0进行单因素方差分析,并进行LSD多重比较。
2 结果与分析
2.1 千粒重
千粒重是体现种子大小与饱满程度的重要指标。方差分析结果表明,海拔对林内种子千粒重(F=53.20,p<0.01)有着显著影响。由图1看出,侧柏种子千粒重随海拔的升高先降低,然后从300 m开始增加至800 m后再降低。多重比较表明,800 m处千粒重最大(p<0.01),300 m 处千粒重最小(p<0.01)。

图1 侧柏种子千粒重随海拔的变化
2.2 种子萌发特征
由图2可看出,海拔200,300,500 m侧柏种子发芽进程相似,在第6天均有部分种子发芽,而后发芽数一直增加;800 m种子第6天才开始发芽,到第13天发芽数上升趋势明显;1 000 m在第9天开始发芽,发芽数少变化趋势不明显。
方差分析结果表明,海拔对侧柏种子发芽率(F=48.07,p<0.01)、发芽势(F=22.86,p<0.01)、发芽指数(F=54.41,p<0.01)、平均发芽时间(F=29.99,p<0.01)和活力指数(F=63.86,p<0.01)都有着极显著影响。随着海拔的增加,发芽率、发芽势、发芽指数和活力指数都是单峰型变化(图3)。多重比较表明,在发芽率和发芽势上,海拔300 m与500 m之间对于发芽指数和活力指数,则是海拔300 m处最大(p<0.01)。在上述指标中,海拔800 m和1 000 m处的数值最低,显著低于其他海拔(p<0.05)。在平均发芽时间上,800 m与1 000 m差异不显著(p>0.05),显著高于其他海拔(p<0.01)。

图2 侧柏林内求同海拔种子发芽进程
2.3 幼苗根系活力
植物根系中的脱氢酶能引起TTC的还原,生成红色而不溶与水的TTF,可以用染色情况及吸收TTC的量来表示根系活力。由图4看出,幼苗根尖酶活力随海拔的变化呈单峰型变化。方差分析结果表明,海拔对根系活力(F=2.493,p=0.11)无显著影响。
2.4 幼苗生物量
方差分析结果表明,海拔对侧柏种子幼苗重量(F=36.75,p<0.01)和单株幼苗重量(F=8.33,p<0.01)有着极显著的影响。由图4看出,侧柏种子幼苗重量随海拔的增加先升高再降低。多重比较结果差异不显著(p>0.05),显著高于其他海拔(p<0.05)。表明,200 m和300 m之间幼苗重量差异不显著(p=0.16),200 m和500 m之间幼苗重量差异不显著(p=0.39),1 000 m 处幼苗重量最小(p<0.01)。单株幼苗重量随海拔升高先下降后升高再下降。多重比较表明,200 m和300 m之间单株幼苗重量差异不显著(p=0.08),显著高于其他海拔(p<0.01),300 m 和800 m之间单株重量差异不显著(p=0.16),500 m和1 000 m之间单株重量差异不显著(p=0.29)。

图4 幼苗根系酶活力随海拔的变化
3 结论和讨论
3.1 海拔对侧柏千粒重的影响

图3 侧柏种子发芽率、发芽势、发芽指数、平均发芽时间和活力指数随海拔的变化

图5 侧柏林内种子幼苗重量和单株重量随海拔的变化
目前关于海拔高度对侧柏种子千粒重影响的报道并不多见。从本研究的结果来看,海拔对于侧柏种子千粒重有显著影响,随海拔的增加呈先降低后升高的趋势,到海拔800 m最大后降低。相关研究表明,野生芥菜型油菜与野生白菜型油菜千粒重随海拔升高而增加,具有垂直分布的生态适应性[13]。垂穗披碱草,相对较低海拔(3 500,3 000 m和2 500 m)的3个种群有良好的适应性,而高海拔(4 000 m)的垂穗披碱草在黄土高原地区的生产性能较低,但4个海拔的种子千粒重表现较小的变异系数[14],这表明千粒重受海拔的影响较小。因此,海拔是植物种子的形成和种子大小的重要影响因素之一,但与植物的适应性密切相关。
3.2 海拔对侧柏发芽能力的影响
高的发芽能力是体现种子活力高的直接指标。天然更新过程中,发芽能力强能够增大野外幼苗建立的可能性[15]。本研究结果表明,随海拔升高,发芽率、发芽势、发芽指数、活力指数都是呈“低-高-低”的变化趋势,中等海拔的林分侧柏种子发芽能力明显优于低海拔和高海拔。这与普布次仁[7]和李续荣[8]的研究结果一致。同时,发芽时间短,种子可以快速的进入到生长阶段,降低腐烂和被微生物侵染,有利于幼苗的建立[4]。本研究发现,在海拔200~500 m的平均发芽时间显著低于海拔800 m和1 000 m,这表明过高的海拔不利于侧柏种子的萌发,这可能与母树长期生长的外界环境因子有关。
3.3 海拔对幼苗生物量的影响
幼苗生物量与天然更新密切相关。个体大的幼苗有着高的成活率。本研究表明,整个培养皿的幼苗生物量在海拔200~500 m之间差异不大,但海拔500 m的单株生物量显著低于海拔200 m。幼苗个体大小的差异可能与母树长期生长的环境有关。
3.4 小 结
种子萌发是天然更新过程中幼苗建立的关键过程。研究表明,随着海拔增加,侧柏种子活力先增加再降低,种子质量最优的海拔范围为300~500 m,种子发芽率超过60%,这与野外调查发现的侧柏有着较好的天然更新能力一致。但在野外更新环境中,侧柏种子萌发与幼苗建立的过程与关键影响因素并不清楚,如母树的年龄、土壤水分、大气温度等,需要在今后做进一步的研究。
[1]GonzálezRodríguez V,Villar R,Casado R,et al.Spatiotemporal heterogeneity effects on seedling growth and establishment in fourQuercusspecies[J].Annals of Forest Science,2011,68:1 2171 232.
[2]Wheeler NC,Guries RP.Population structure,genic diversity,and morphological variation in Pinus contorta Dougl[J].Canadian Journal of Forest Research,1982,12:595606.
[3]Steven D D.Experiments on mechanisms of establishment in oldfield succession:seedling emergence[J].Ecology,1991,72:1 0661 075.
[4]Cendán,C.,L.Sampedro and R.Zas.The maternal environment determines the timing of germination inPinus pinaster.[J].Environ.Exp.Bot,2013,94:6672.
[5]Beckstead,J.,S.E.Meyer,B.M.Connolly,M.B.Huck and L.E.Street.Cheatgrass facilitates spillover of a seed bank pathogen onto native grass species.J.[J].Ecol,2010,98:168177.
[6]阿里别里根·哈孜太.不同海拔对不同春小麦种子活力的影响[D].新疆农业大学,2015.
[7]普布次仁,赵垦田,杨小林.温度对不同海拔梯度急尖长苞冷杉种子发芽的影响[J].西部林业科学,2014(4):132135.
[8]李续荣,程小林,曹亚凤,等.海拔高度对独活种子产量和质量的影响[J].种子科技,2016(8):120122.
[9]鲁法典,韩峻,李超.泰山侧柏林下幼树分布规律及天然更新研究[J].河北林果研究,2006(1):2932.
[10]李健,孙仲序.泰山侧柏[J].山东林业科技,1998(6):2627.
[11]国家质量技术监督局.林木种子检验规程[M].北京:中国标准出版社,1999.
[12]张志良,瞿伟菁.植物生理学实验指导(第3版)[M].北京:高等教育出版社,2003.
[13]卓嘎,旦巴,何燕,等.西藏野生油菜种质资源千粒重分析[J].种子,2008,27(12):7276.
[14]刘婷娜,王彦荣,胡小文.不同海拔垂穗披碱草种子产量及其构成因素[J].草业科学,2014(3):468473.
[15]毛培利,韩广轩,张志东,等.黑松海岸防护林径级分布及其与球果和种子特性的关系[J].东北林业大学学报,2010,38(11):2123.
