甜菜M14品系盐胁迫转录组数据库的转录因子分析
2018-01-12端木慧子王建慧李海英马春泉
端木慧子,陶 鑫,王建慧,韦 恒,李海英,马春泉,*
(黑龙江大学 a.生命科学学院;b. 黑龙江省普通高等学校分子生物学重点实验室,哈尔滨 150080)
盐胁迫是制约植物生长发育的重要因素之一,盐渍化的土地在中国的东北、华北、西北地区广泛存在,因此培育和研究耐盐作物对提高农作物产量有着重要的意义[1]。在植物抵御非生物胁迫过程中,植物转录因子扮演着重要的角色,它调节基因转录调控的表达,是植物适应外界环境的分子基础,转录调控是基因表达调控的关键步骤[2]。
甜菜M14品系是郭德栋等通过二倍体栽培甜菜和四倍体野生白花甜菜进行杂交和回交得到的甜菜单体附加系,研究表明甜菜M14品系具有野生白花甜菜抗寒、抗旱、耐盐及无融合生殖等优良特性[3-6]。实验室前期通过对0、200、400 mM NaCl处理下甜菜M14品系根和叶的转录组测序得到甜菜M14品系的转录组数据库。
研究发现参与盐胁迫应答的转录因子主要都是来自AP2/ERF、MYB、WRKY、NAC、bZIP、HD-ZIP这6个基因家族。WRKY家族是仅存在于植物中,在非生物胁迫以及生物胁迫中起着各种各样的作用。WRKY 转录因子与其下游靶基因和上游调节基因之间的相互作用和交互作用构成了复杂的 WRKY转录因子调控网络,这是研究界的新兴领域之一[7-9]。前人对拟南芥、棉花、水稻等植物的WRKY家族转录因子做了大量的研究,如小麦中的TaERKY70在小麦抗条锈菌的过程中起着重要的作用,当其被沉默时会导致植株对小麦条锈病的敏感性增高[10];棉花GhWRKY68与ROS的积累、MDA含量升高以及ROS相关基因的表达相关[11];研究发现WRKY在植物激素赤霉素和脱落酸介导信号转导途径中发挥重要作用[12]。棉花、拟南芥、水稻中分别有17、18、26个WRKY家族的转录因子响应盐胁迫[13-15],但很少有人对甜菜WRKY家族的转录因子进行研究。
本研究对课题组前期构建的甜菜M14品系盐胁迫下转录组数据库进行系统的分析,基于文献、植物转录因子数据库等信息对其中的转录因子进行筛选与鉴定,从而获得非生物胁迫应答相关的转录因子。进一步对筛选出的转录因子WRKY家族进行进化分析并构建其蛋白互作网络,采用Real-time PCR技术分析WRKY基因家族转录因子盐胁迫前后表达量,初步探究WRKY家族转录因子在甜菜M14抗盐胁迫机制中的表达模式,为其进一步功能研究奠定基础。
1 材料与方法
1.1 材料
本研究使用的实验材料为甜菜M14品系(公开号:CN1263695A)、甜菜M14品系转录组数据库。
1.2 方法
1.2.1 甜菜M14品系转录组数据库中的转录因子分类
通过植物转录因子数据库、非冗余Nr库和Swissprot数据库中的信息筛选甜菜M14品系转录组数据库中的转录因子,并进行分类。将分类完成的转录因子相关的基因序列一一查找并对应到KEEG注释的代谢通路中所标识的位置。
1.2.2 甜菜M14品系WRKY转录因子的系统发生分析
将甜菜M14品系数据库中的WRKY家族转录因子的序列在NCBI在线Blastp (https://blast.ncbi.nlm.nih.gov/Blast.cgi)进行分析,选取与甜菜WRKY亲缘较近的物种的WRKY核酸序列进行比对分析,用MEGA6.0软件进行系统发育树的构建。
1.2.3 甜菜M14品系WRKY转录因子的蛋白质互作网络构建
根据String10.0数据库(http://www.string-db.org/)构建甜菜M14品系转录组数据库中筛选出的WRKY转录因子的互作网络。
1.2.4 甜菜M14品系WRKY转录因子盐胁迫下基因表达分析
分别提取0、200、400 mM NaCl处理下甜菜M14品系叶组织的总RNA,DNase纯化处理后及浓度稀释均一后,分别取1.0 μL总RNA,采用TaKaRa RNA PCR Kit反转录试剂盒进行反转录合成cDNA。WRKY转录因子基因实时荧光定量PCR反应使用TaKaRa公司的SYBR染料,在Bio-Rad iQ5荧光定量PCR仪上进行。甜菜M14品系WRKY家族转录因子基因的扩增以18S rRNA基因作为内参并采用相同的反应体系,其中上游引物标注为18S,下游引物标注为18AS,WRKY家族转录因子基因上游引物标注为F,下游引物标注为R,数字代表基因编号(表1),每个反应设置3次技术重复和3次生物重复。以0 mM NaCl处理下甜菜M14品系WRKY家族基因的Ct值为对照组,进行数据分析。

表1 Real time PCR引物名称和序列
2 结果与分析
2.1 甜菜M14品系转录组数据库中转录因子家族分类
参照植物转录因子数据库和文献,对甜菜M14品系转录组数据库中的转录因子进行分类。在甜菜M14品系转录组数据库中,Nr功能注释下的数据库表格中筛选出了768个转录因子,Swissprot功能注释下的数据库表格中筛选出了541个转录因子,将Nr与Swissprot的转录因子进行桥接获得了623个转录因子,包含34个转录因子家族(表2)。

表2 甜菜M14品系数据库转录因子家族分类
根据植物转录因子数据库网站(http://planttfdb.cbi.pku.edu.cn)将这623个转录因子按功能分类可分为5类,即生长发育、生物与非生物胁迫、信号传导、染色质及染色体形成和其它(表3)。
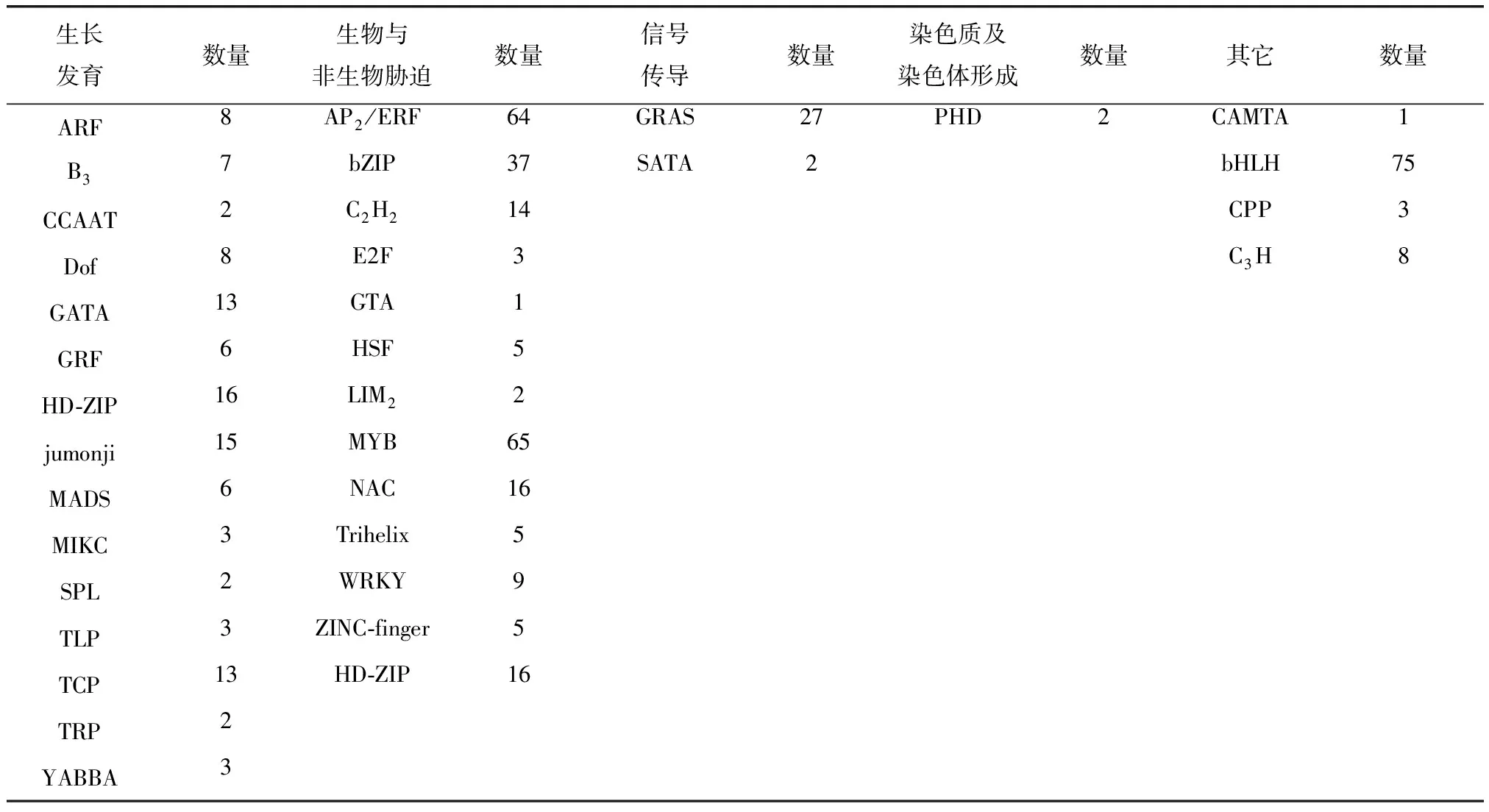
表3 甜菜M14品系转录因子家族功能分类
2.2 数据库中转录因子的KEGG代谢通路分析
甜菜M14品系转录因子KEEG注释的结果显示转录因子参与植物的各种代谢途径,其中参与调节植物昼夜节律和植物抵抗病原体通路的转录因子最多,WRKY家族的转录因子则在植物切除修复的通路上发挥着重要作用(表4)。通过对应KEGG的代谢通路,可以初步了解不同转录因子家族在不同的代谢通路中的作用,为以后实验验证,以及改良物种有很好的作用。
2.3 WRKY转录因子及近缘物种系统进化树
对甜菜M14品系的WRKY家族转录因子及其他物种的WRKY家族转录因子进行系统发育分析,结果显示甜菜M14品系的WRKY家族转录因子与拟南芥WRKY家族转录因子亲缘关系最近(图1)。
2.4 WRKY转录因子的蛋白质互作网络构建
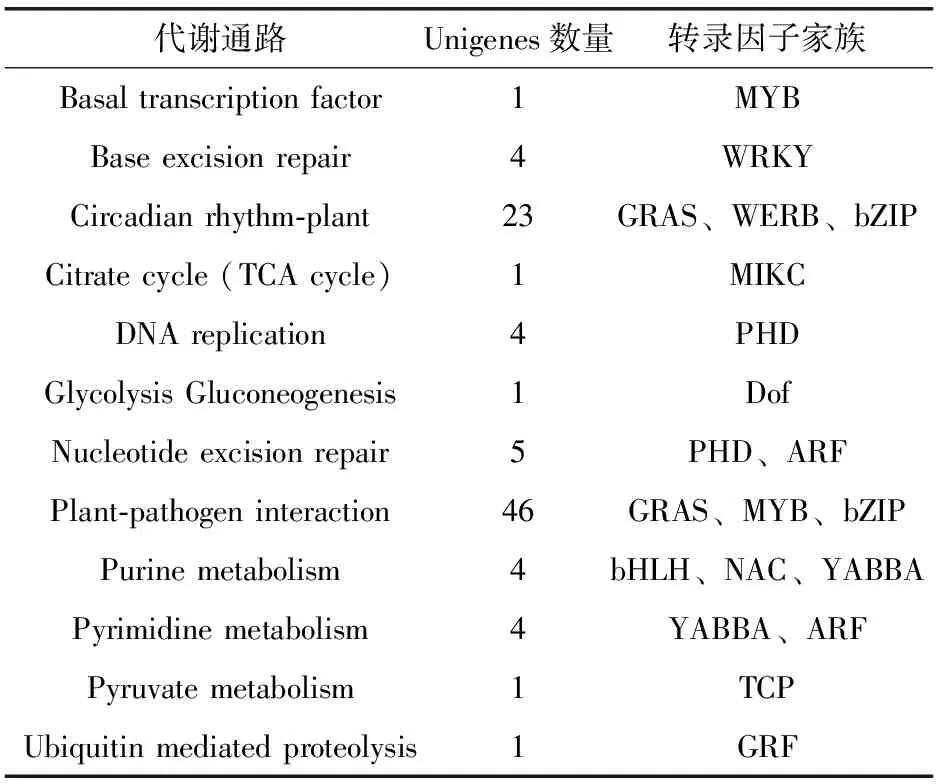
表4 甜菜M14品系转录因子的KEGG代谢通路
转录因子是高等植物生命活动中不可或缺的调控因子,具有复杂的网络调控关系。根据甜菜M14品系中WRKY转录因子的拟南芥同源基因,利用STRING数据库构建WRKY蛋白质互作网络。结果显示,甜菜WRKY3、WRKY13、WRKY15、WRKY32的拟南芥同源基因均具有多个互作的蛋白质,形成复杂的蛋白质互作网络(图2),因此这些WRKY转录因子可能行使比较重要的功能。

图1 WRKY转录因子家族相关物种系统进化树Fig.1 Phylogenetic tree of WRKY transcriptional factors

图2 拟南芥WRKY转录因子家族的STRING分析图Fig.2 Phylogenetic tree of WRKY transcriptional factors
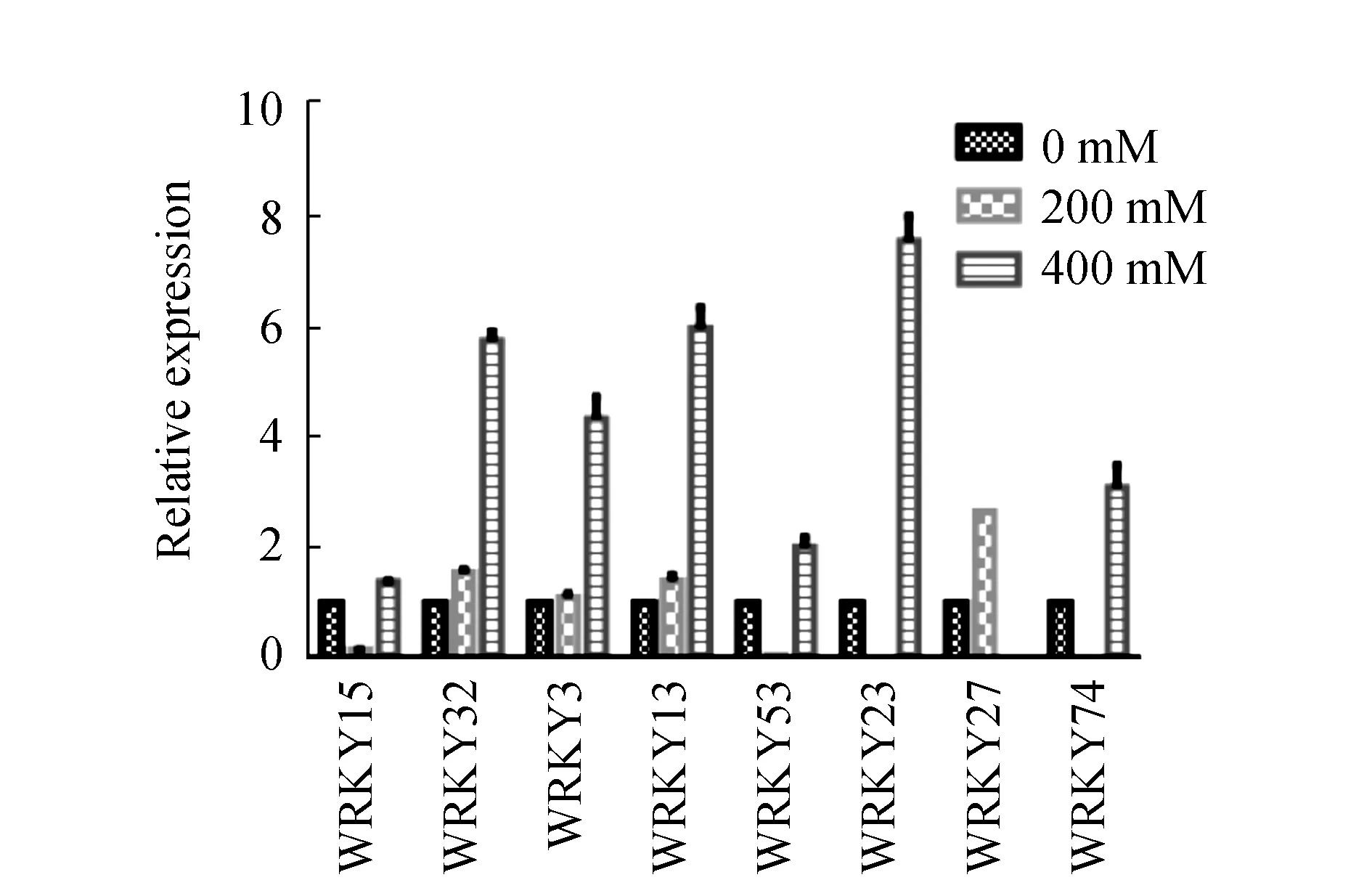
图3 盐胁迫下WRKY家族转录因子基因Real-time定量结果Fig.3 Real-time PCR results of WRKY transcriptional factors of sugar beet M14 under NaCl stress
2.5 盐胁迫下WRKY家族转录因子基因的表达模式分析
根据Real time PCR的结果,计算2-△△Ct制作柱形图,见图3。由图3可见,WRKY15和WRKY53受到低浓度盐胁迫的抑制,在高浓度盐胁迫下上调;WRKY32、WRKY3、WRKY13在低浓度和高浓度盐胁迫下表达量都上调。WRKY23、WRKY27、WRKY74在低浓度盐胁迫下不表达,高浓度盐胁迫下表达上调。
3 讨 论
本研究在甜菜M14品系0 、200、400 mM NaCl浓度胁迫下的转录组数据库中,共筛选到了623个转录因子,分属34个家族。按照功能可将其分为参与植物生长发育、响应生物与非生物胁迫、参与信号传导、调控染色质及染色体的形成和其它5大类,其中有9个WRKY转录因子具有响应生物与非生物胁迫功能。
WRKY家族转录因子在拟南芥、玉米、大豆等作物响应胁迫的过程中发挥着举足轻重的作用,但很少有人对甜菜中的WRKY家族转录因子进行过研究。因此,本实验对甜菜M14品系中的WRKY家族转录因子WRKY15、WRKY32、WRKY3、WRKY13、WRKY53、WRKY23、WRKY27和WRKY74进行了系统的分析。结果显示,WRKY3、WRKY13、WRKY15和WRKY32同源的拟南芥WRKY家族转录因子具有复杂的蛋白互作网络,具有多个互作蛋白质;WRKY15、WRKY32、WRKY3、WRKY13、WRKY53、WRKY23、WRKY27和WRKY74的表达均受到NaCl的诱导,而且可以推测甜菜M14品系WRKY32、WRKY3、WRKY13在NaCl胁迫下稳定的诱导表达,WRKY23、WRKY74和WRKY27的表达则具有相互补充的作用,即在低浓度NaCl(200 mM)胁迫下WRKY27受到大量诱导,在高浓度NaCl(400 mM)胁迫是WRKY23、WRKY74受到大量诱导。
2008年Zhou等将大豆中的3个转录因子GmWRKY13、GmWRKY21和GmWRKY54在拟南芥中过表达, 发现在200 mM NaCl胁迫下GmWRKY13的过表达植株可抗盐害[16]。笔者的结果同样显示,甜菜M14品系中的转录因子WRKY13在200、400 mM NaCl处理下表达量均发生上调,表明甜菜M14品系中转录因子WRKY13可能是响应盐胁迫的正调控因子。Wei 等将遏蓝菜中的TcWRKY53转录因子在烟草中过表达后发现TcWRKY53是响应渗透胁迫的负调控因子[17]。众所周知,NaCl胁迫往往会伴随着渗透胁迫的发生,笔者发现在200 mM NaCl胁迫下转录因子WRKY53的表达量下调,表明甜菜M14品系中转录因子WRKY53可能是响应低盐胁迫的负调控因子。
另外,甜菜M14品系中的转录因子WRKY15、WRKY3、WRKY32、WRKY23、WRKY27和WRKY74在盐胁迫下表达量也均发生变化,但在其它作物中并没有报道,后续笔者将会对其功能进行进一步验证。
4 结 论
本研究通过对前期构建的甜菜M14品系盐胁迫转录组数据库进行分析,结合已知的转录因子数据库与注释信息,对甜菜M14品系中的转录因子进行了分类与注释,并对筛选到的响应盐胁迫的甜菜WRKY转录因子进行了蛋白质互作网络分析与表达分析,最终获得了可能参与盐胁迫应答的甜菜WRKY转录因子。本研究将为甜菜盐胁迫响应研究提供理论基础,为甜菜中转录因子研究提供数据支持,并为WRKY家族转录因子功能的进一步研究奠定基础。
[1] Khan M A, Ungar I A, Showalter A M. Effects of salinity on growth, ion content, and osmotic relations inHalopyrummucronatum(L.) Stapf[J]. Journal of Plant Nutrition, 1999, 22(1): 191-204.
[3] 郭德栋,芳晓华,刘丽萍,等.无融合生殖甜菜单体附加系的获得和鉴定[J]. 云南大学学报:自然科学版, 1999, 23(S3): 180-181.
[4] Li H, Cao H, Wang Y, et al. Proteomic analysis of sugar beet apomictic monosomic addition line M14[J]. Journal of Proteomics, 2009,73: 297-308.
[5] 吕笑言,金英.利用生物信息学技术构建分析甜菜M14品系盐胁迫下参考转录组数据库[J].黑龙江大学自然科学学报,2017,34(2):208-212.
[6] 蒋德生,李海英,于冰,等.甜菜M14品系BvM14-APX、BvM14-DHAR3、BvM14-MDAR基因响应NaCl胁迫的表达分析[J].黑龙江大学工程学报,2016,7(2):64-71.
[7] Berri S, Abbruscato P, Faivre-Rampant O, et al. Characterization of WRKY co-regulatory networks in rice and Arabidopsis [J]. BMC Plant Biology, 2009, 9(1):120-131.
[8] Zentgraf U, Laun T, Miao Y. The complex regulation of WRKY53, during leaf senescence ofArabidopsisthaliana[J]. European Journal of Cell Biology, 2010, 89(2-3): 133-137.
[9] Banerjee A, Roychoudhury A. WRKY proteins: signaling and regulation of expression during abiotic stress responses[J]. The Scientific World Journal, 2015, 807560-807576.
[10] Wang J J, Tao F, An F, et al. Wheat transcription factor TaWRKY70 is positively involved in high-temperature seedling plant resistance toPucciniastriiformisf. sp.tritici[J]. Molecular Plant Pathology, 2016, 18(5): 649- 661.
[11] Jia H, Wang C, Wang F, et al. GhWRKY68 reduces resistance to salt and drought in transgenic Nicotiana benthamiana[J]. PLoS One, 2015, 10(3): e0120646.
[12] 李冉, 娄永根. 植物中逆境反应相关的 WRKY 转录因子研究进展[J]. 生态学报, 2011, 31(11): 3223-3231.
[13] Cai C P, Niu E L, Du H, et al. Genome-wide analysis of the WRKY transcription factor gene family inGossypiumraimondiiand the expression of orthologs in cultivated tetraploid cotton [J]. Acta Agron Sin, 2014, 2(z1): 87-101.
[14] Deyholos Michael K, Jiang Y Q. Comprehensive transcriptional profiling of NaCl-stressed Arabidopsis roots reveals novel classes of responsive genes [J]. BMC Plant Biology, 2006, 6(1):25-44.
[15] Ramamoorthy R, Jiang S Y, Kumar N, et al. A comprehensive transcriptional profiling of the WRKY gene family in rice under various abiotic and phytohormone treatments [J]. Plant and Cell Physiology, 2008, 49(6):865-879.
[16] Zhou Q Y, Tian A G, Zou H F, et al. Soybean WRKY-type transcription factor genes, GmWRKY13, GmWRKY21, and GmWRKY54, confer differential tolerance to abiotic stresses in transgenic Arabidopsis plants[J]. Plant Biotechnology Journal, 2008, 6(5):486-503.
[17] Wei W, Zhang Y, Han L, et al. A novel WRKY transcriptional factor from Thlaspi caerulescens negatively regulates the osmotic stress tolerance of transgenic tobacco[J]. Plant Cell Reports, 2008, 27(4):795-803.
