褐藻胶裂解酶的基因克隆及表达条件优化
2017-12-28黄桂媛王巧贞卢明倩张荣灿张云开黄庶识
许 超,黄桂媛,王巧贞,卢明倩,张荣灿,廖 威,张云开*,黄庶识
(1.广西科学院 广西海洋天然产物与组合生物合成重点实验室,广西 南宁 530007;2.广西大学 生命科学与技术学院,广西 南宁 530003;3.广西北部湾海洋研究中心,广西 南宁 530007;4.广西职业技术学院,广西 南宁 530226)
褐藻胶裂解酶的基因克隆及表达条件优化
许 超1,2,黄桂媛1,王巧贞1,卢明倩1,张荣灿3,廖 威4,张云开2*,黄庶识1
(1.广西科学院 广西海洋天然产物与组合生物合成重点实验室,广西 南宁 530007;2.广西大学 生命科学与技术学院,广西 南宁 530003;3.广西北部湾海洋研究中心,广西 南宁 530007;4.广西职业技术学院,广西 南宁 530226)
采用大肠杆菌(Escherichia coli)表达海洋弧菌(Vibriosp.)X511的褐藻胶裂解酶基因,以实现对该酶的大量制备及酶学性质研究。通过序列分析,预测该酶的前20个氨基酸为信号肽,去除信号肽后的蛋白质分子质量约为30 254.28 Da,等电点8.75,分子式为C1368H2100N364O401S6,对应的基因序列命名为alg1987。按如下方案构建重组菌:设计特异性核酸引物,PCR扩增得到alg1987;将重组质粒pET-30(a)-alg1987导入大肠杆菌Trans5α进行测序;提取阳性pET-30(a)-alg1987质粒导入大肠杆菌BL21,利用异丙基-β-D-硫代半乳糖苷(IPTG)诱导表达。十二烷基硫酸钠聚丙烯酰胺凝胶电⒕(SDS-PAGE)检测诱导后的重组菌胞液上清,发现异源表达的褐藻胶裂解酶分子质量大小与预测值相近。对重组菌中的褐藻胶裂解酶进行表达条件优化,结果显示,最适参数为诱导温度16℃,诱导时间16 h,诱导剂IPTG浓度0.8 mmol/L。
褐藻胶裂解酶;基因;克隆;条件优化;表达
褐藻胶包括海藻酸及其可溶性盐,是地球上可再生碳水化合物中总储量仅次于纤维素的多糖物质[1-3],广泛存在于海洋褐藻门植物的细胞壁外层及内层,其含量可达到褐藻干质量的10%~40%[4-5]。组成褐藻胶的基本单糖是β-D-甘露糖醛酸(β-D-mannuronic acid,M)和α-L-古罗糖醛酸(α-L-guluronic acid,G),M与G互为差向异构体,仅C5所连羧基的位置存在差异[4]。在褐藻胶中通常存在聚甘露糖醛酸(ploymannuronic acid,pM)、聚古罗糖醛酸(polyguluronic acid,pG)、pM/G、M/G共4种结构单元,以随机比例通过1,4-糖苷键的聚合作用形成直链、无分支、具有负电荷的线性大分子多糖[6]。此外,在少数细菌如假单胞菌和固氮菌属等细菌中也存在褐藻胶,但一般只含多聚甘露糖醛酸[7],而无多聚古罗糖醛酸[8]。褐藻胶广泛应用于饲料、食品、农业及医药[9-11]等多个行业。但是褐藻胶分子质量大,黏度高,不容易被直接使用[12],而通过对褐藻胶进行降解或改性的方法,可得到可发酵的寡糖及的衍生物,且具有特殊生物活性,拓宽了褐藻胶应用领Ⅱ[7]。
目前水解褐藻胶方法可分为3类:①物理降解,包括热解、辐射和超声降解,此方法的操作过程较为繁琐,且存在能耗大及原料转化率低的问题,在工业上应用得不多[13];②化学降解,包括酸碱及氧化降解,酸水解法是目前比较主流的一种方法,该方法操作繁琐,反应过程难以控制,废弃物处理难度大[14-15];③生物酶法,应用褐藻胶裂解酶进行水解作用获得寡糖或者单糖,与物理或者化学法相比,生物酶法工艺效率高,能耗少,环境友好,优势明显。
褐藻胶裂解酶来源广泛,在海洋藻类、海洋软体动物和棘皮动物以及微生物(包括土壤微生物、海洋微生物、噬菌体和少数病毒)中均有报道。在海洋微生物中,又以弧菌属(Vibrio)、芽孢杆菌属(Bacillus)、交替单胞菌(Alteromonas)、假单胞菌属(Pseudomonas)、交替假单胞菌属(Pseudoaltermonas)、鞘氨醇菌属(Sphingosinas)和链霉菌属(Streptomyces)等被研究的最多[16-18]。褐藻胶在褐藻胶裂解酶(alginatelyase)的作用下生成不饱和醛糖酸[19],可经非酶促反应转变为4-脱氧-L-赤式-5-己酮糖糖醛酸(4-deoxy-L-erythro-5-hexoseulose uronate,DEH)[20-21],DEH可被还原酶作用变成2-酮基-3-脱氧-D-葡萄糖酸(2-keto-3-deoxy-D-gluconate,KDG),进而醛酸为丙酮酸进入糖酵解途径被利用[22-23]。
随着基因工程技术的发展,通过构建工程菌对某一特定功能的基因进行大量表达,是实现工业化生产的有效途径。目前国内外对褐藻胶裂解酶的异源表达也做了大量研究,如SHINABARGER D等[24-25]分别将铜绿假单胞菌(Pseudomonas aeruginosa)和褐球固氮菌(Azotobacter chroococcum)的褐藻胶裂解酶基因克隆至大肠杆菌(Escherichia coli)并成功表达;刘航等[26-27]利用大肠杆菌分别表达得到了紫色链霉菌(Treptomyces violaceoruber)和海洋弧菌QY105的褐藻胶裂解酶基因。此外,TAKEDAH等[28]通过整合鞘氨醇菌(Sphingomonassp.)A1和运动发酵单胞菌(Zymomonas mobilis)中褐藻胶的吸收及转化的多个基因,并阻断了代谢通路上副产物的代谢途径,率先在实验室以褐藻胶为唯一碳源利用重组菌株A1生产乙醇,其发酵3d后的乙醇产量达到13.0g/L。WARGACKIAJ等[29]将褐藻胶降解及乙醇合成途径上的多个关键酶基因导入大肠杆菌的染色体,得到了可直接利用褐藻胶生产乙醇的重组菌,其乙醇产量可到达到理论值的80%。ENQUISTNEWMAN M等[30]构建的可直接将褐藻胶转化为乙醇的酵母工程菌株达到了工业生产的要求。
本研究克隆筛自北部湾海Ⅱ的海洋弧菌(Vibriosp.)X511中的褐藻胶裂解酶基因,构建重组质粒导入大肠杆菌(Escherichia coli),通过条件优化,实现该酶的异源大量表达,以期为大量制备褐藻胶裂解酶及进一步研究提供数据支持。
1 材料与方法
1.1 材料与试剂
1.1.1 试剂
海藻酸钠:阿拉丁试剂有限公司;酵母浸膏、蛋白胨:英国Oxoid公司;七水硫酸镁、硫酸铵、氯化钠、磷酸氢二钾、硫酸亚铁:国药集团化学试剂公司;所有试验所用试剂均为分析纯。细菌总DNA提取试剂盒:杭州博日科技公司;PCR高保真ExTaq酶、DNA及RNA Marker、DNA连接试剂盒:大连TakaRa公司;限制性内切酶Nde I、Xho I:美国NEB公司;质粒提取试剂盒、琼脂糖凝胶回收试剂盒:美国Bioflux生物公司。
1.1.2 菌株及质粒
海洋弧菌(Vibriosp.)X511为本实验室筛选保存;质粒pET-30(a):上海捷瑞生物公司;大肠杆菌(Escherichia coli)Trans5α、BL21(DE3):北京全式金生物技术有限公司。
1.1.3 培养基
海藻酸钠培养基:海藻酸钠6 g/L,NaCl 15 g/L,硫酸铵5 g/L,磷酸氢二钾1 g/L,硫酸镁1 g/L,硫酸亚铁0.001 g/L,pH 7.5。
活化培养基:海藻酸钠培养基中的硫酸铵用蛋白胨替代,其他成分保持不变。
LB培养基:胰蛋白胨10 g/L,酵母提取物5 g/L,氯化钠10 g/L,pH调至7.0。
LA培养基:在LB培养基的基础上添加1.5%的琼脂。以上培养基均在121℃条件下灭菌20 min备用。
1.2 仪器与设备
TProfessionalstandard96GradientPCR仪:德国Biometra公司;UNIVERSAL 32R台式冷冻离心机:德国Hettich科学仪器公司;TU-1901双光束紫外可见分光光度仪:北京普析通用仪器公司;OmegaFlour凝胶成像仪:美国Aplegen公司。
1.3 方法
1.3.1 序列分析
借助SignalP 4.1 Server(http://www.cbs.dtu.dk/services/SignalP/)分析褐藻胶裂解酶基因的信号肽;利用ExPASy(http://web.expasy.org/compute_pi/)预测目的蛋白质的分子质量及等电点信息;在美国国家生物技术信息中心(National Center of Biotechnology Information,NCBI)(https://www.ncbi.nlm.nih.gov/)蛋白质数据库中Blast目的蛋白,下载同源序列,利用MEGA6.0软件基于邻接法(Neighbor-Joining,NJ)法构建系统发育树。
1.3.2 基因克隆
取1 mL于-80℃冰箱中保藏的菌种X511至活化培养基中活化,按1%的接种量转入海藻酸钠培养基培养过夜,培养条件均为30℃、200 r/min。根据细菌总脱氧核糖核酸(deoxyribonucleic,DNA)提取试剂盒的操作步骤,提取菌株X511的总DNA经稀释后作为聚合酶链反应(polymerase chain reaction,PCR)模板。
去除信号肽后的目的基因长度为798 bp,采用Vector NTI Advance 11.5软件设计引物,分别在引物两段添加限制性酶切位点及保护碱基:alg1987-F:GGAATTC CATATG TCTGATGTAAATAATGGTATCG(下划线部分为Nde玉酶切位点);alg1987-R:CCG CTCGAG TTACTTGCCGTTTAGCTTAA(下划线部分为Xho玉酶切位点)。委托北京奥科生物公司进行引物合成。PCR总反应体系为:ExTaq酶1μL,6xExTaq酶Buffer5μL,脱氧核糖核苷三磷酸(deoxyribonucleoside triphosphate,dNTP)混合物2.0 μL,上游引物(F)2.0 μL,下游引物(R)2.0 μL,DNA模板2.0 μL,加ddH2O至25μL。PCR扩增条件为:98℃预变性5min,98℃变性10s,50℃退火15 s,68℃延伸50 s,68℃延长5 min,16℃保存;循环数为30次。
1.3.3 表达载体的构建
根据试剂盒操作步骤,切胶回收PCR扩增产物,在37℃条件下对目的基因及pET-30(a)载体进行双酶切,体系为:CutSmartTM 5 μL,Nde玉2 μL,Xho玉2 μL,加入目的基因至50μL,反应时长2h;分别切胶回收目的基因及载体的双酶切产物,于16℃按以下条件过夜连接,体系为:Solution玉10 μL,pET-30a 4 μL,目的DNA 12 μL;将重组载体pET-30(a)-alg1987转化至大肠杆菌感受态细胞Trans5α,挑选阳性转化子送上海生工公司测序,提取序列正确的重组质粒转化至感受态细胞大肠杆菌BL21(DE3)[31]。
1.3.4 优化诱导表达条件
挑取阳性转化子接至LB培养基中培养过夜,得到种子液;以1%的接种量将种子液转接至LB培养液中,振荡培养至OD600nm介于0.6~0.8之间,加入异丙基-β-D-硫代半乳糖苷(isopropyl-β-D-thiogalactoside,IPTG)至终浓度为0.4 mmol/L,分别为16℃、25℃、37℃三组不同温度条件下诱导表达,在诱导时间为0、2 h、4 h、8 h、16 h、24 h时取样,测定重组菌破胞液中的褐藻胶裂解酶粗酶活性,选取粗酶活性最高时对应的条件作进一步优化。
IPTG可诱导重组菌中异源基因的表达,但其自身对微生物细胞体具有一定的毒害作用,过高的浓度甚至会杀死细胞,影响重组菌的表达效率。为得到较优的添加量,分别对IPTG终浓度为0、0.2 mmol/L、0.4 mmol/L、0.6 mmol/L、0.8 mmol/L、1.0 mmol/L进行优化,选择褐藻胶裂解酶粗酶活性最高时对应的IPTG浓度。
1.3.5 褐藻胶裂解酶活性测定
将培养好的菌液于6 000 r/min、4℃条件下离心10 min收集菌体,用浓度为0.1mol/L,pH值为7.5的Tris-HCl等体积重悬;在冰浴条件下超声20 min破碎菌体,8 000 r/min,4℃离心5 min,取上清作为粗酶。酶活测定方法为:向900 μL底物(海藻酸钠溶于0.1mol/LpH值为7.5的Tris-HCl至质量体积比为0.2%)中加入100μL粗酶液,30℃水浴反应10min,12 000 r/min条件下离心2 min,测定上清液在波长235 nm条件下的OD235nm值[26],定义光密度值每分钟上升0.01为一个酶活力单位(U)。
相对酶活是指定义同一批试验中酶活性最高一组的为100%,其他试验组的相对酶活性为该试验组与最高组之间酶活性的比值。
1.3.6 SDS-PAGE电泳
将诱导后的重组菌破胞液液进行十二烷基硫酸钠聚丙烯酰胺凝胶电⒕(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE),以初步确定重组菌表达的褐藻胶裂解酶的分子质量。
2 结果与分析
2.1 序列分析结果
采用SignalP4.1Server分析褐藻胶裂解酶基因(见图1A)对应的蛋白质序列(全长共285个氨基酸),预测该蛋白质的前20个氨基酸为信号肽序列(见图1B下划线部分)。

图1 目的基因及蛋白质序列Fig.1 Target gene and protein sequence
利用ExPASy分析不含信号肽的褐藻胶裂解酶序列,预测其理论的分子质量大小为30 254.28 Da,等电点8.75,分子式为C1368H2100N364O401S6。在NCBI中进行蛋白质序列比对,发现该酶与褐藻胶裂解酶PL-7家族中多个酶的结构Ⅱ较为相似,下载同源序列,利用MEGA6.0软件构建的系统发育树,结果见图2。

图2 目的蛋白的系统发育树Fig.2 Phylogenetic tree of the target protein
由图2可知,褐藻胶裂解酶Alg1987与噬琼胶菌属菌株Agarivoranssp.JAM-Alm中的褐藻胶裂解酶BAH70324.1以及白色噬琼胶菌属菌株Agarivorans albus中的褐藻胶裂解酶WP 040307007.1和Agarivorans albusMKT 106中的褐藻胶裂解酶GAD01352.1一致性最高,为81%,远低于同种酶之间的95%,说明Vibriosp.X511 Alg1987可能是一种新型褐藻胶裂解酶。
利用SWISS-MODEL对褐藻胶裂解酶Alg1987的第29~259个氨基酸进行在线同源建模,得到多个推荐模型。由图3可知,与Alg1987同源性最高的模型为2za9.1.A,两者的相似性仅为23.08%,该模型富的C端富含β折叠,其余部分为α螺旋。

图3 褐藻胶裂解酶Alg1987的三维结构模拟图Fig.3 Three-dimensional structure simulation diagram of alginate lyase Alg1987
2.2 基因克隆及表达载体的构建
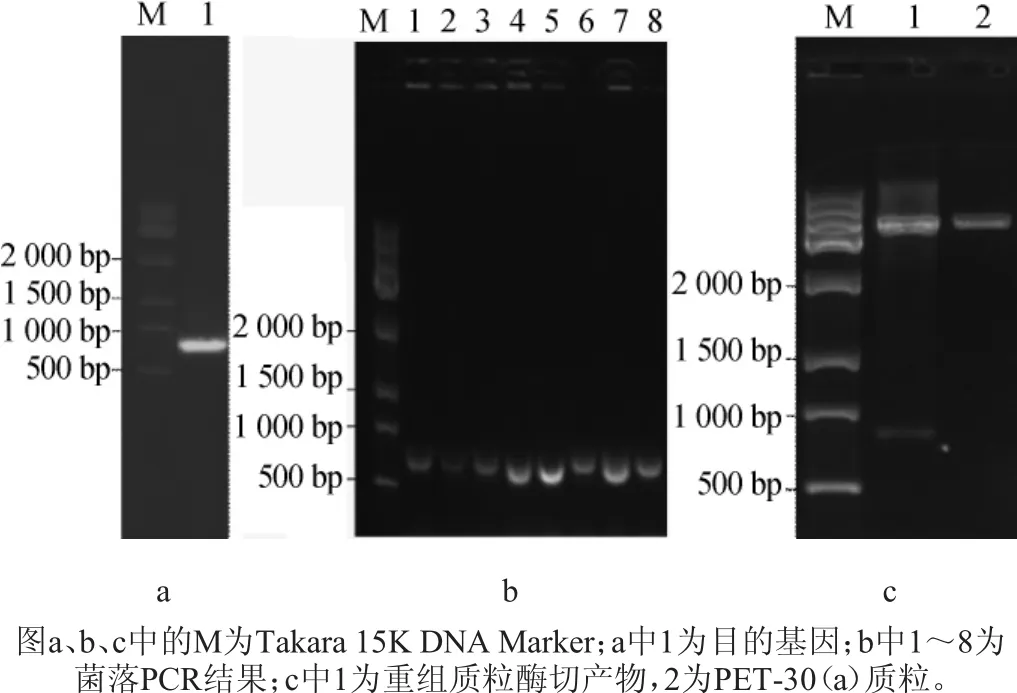
图4 琼脂糖凝胶电⒕结果Fig.4 Results of agarose gel electrophoresis
以海洋弧菌Vibriosp.X511的总DNA为模板进行PCR扩增,结果见图4。由图4a可知,得到的扩增产物长度约800bp,采用NCBI将目的基因的序列与测序结果进行在线比对,结果显示两者的相似度为100%,说明已成功获得目的基因;由图4b可知,菌落PCR的凝胶电⒕结果显示,多个大肠杆菌转化子均能扩出与目的基因长度一致的条带,初步证明目的基因已成功导入受体菌。由图4c可知,提取菌落PCR验证结果为阳性转化子的质粒,用Nde玉和Xho玉双酶切质粒,以空载的pET-30(a)质粒为对照,经0.4%琼脂糖凝胶电⒕后发现重组质粒中含有与目的基因大小一致的片段(图4c中1号⒕道),进一步说明目的基因可能已成功导入了质粒中。将质粒送至生物公司测序并比对,显示该质粒中存在与目的基因相似度为100%的DNA片段,重组质粒构建成功,-80℃甘油保藏含目的基因的Trans5α克隆载体。
2.3 褐藻胶裂解酶的表达优化
2.3.1 诱导温度及时间
分别优化重组菌的诱导剂添加量及诱导时间,结果见图5。由图5可知,在诱导时间4 h以内,3组不同的诱导温度对重组菌表达褐藻胶裂解酶几乎没有影响;随着培养时间的延长,重组菌在较低诱导温度下表达的褐藻胶裂解酶粗酶活性较高;在诱导温度为16℃时,重组菌经16 h的诱导后,其超声破碎细胞液上清中的褐藻胶裂解酶活性达到最大值,继续诱导至24 h,酶活性无明显变化;在诱导温度分别为25℃和37℃条件下诱导8h时,重组菌的破碎细胞液中开始出现包涵体,且褐藻胶裂解酶粗酶活性明显较低。故选择诱导温度16℃,诱导时间16 h为诱导条件。
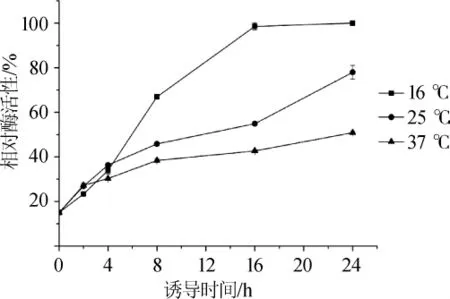
图5 不同诱导温度对重组菌表达褐藻胶裂解酶的影响Fig.5 Effects of different induction temperatures on the expression of aginate lyase in recombinant bacteria
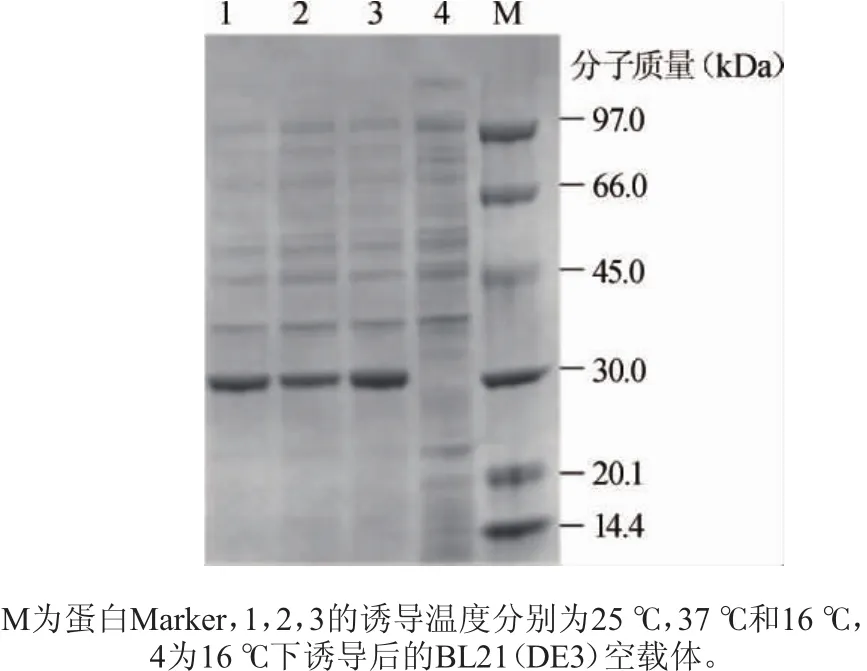
图6 不同诱导温度下的重组菌破碎细胞液上清Fig.6 Supernatant of recombinant bacteria cytosol at different induction temperatures
分别取16℃、25℃及37℃诱导温度条件下诱导16 h的重组菌破胞液上清进行SDS-PAGE电⒕,结果见图6。由图6可知,重组菌在3个不同诱导温度下大量表达的蛋白质分子质量大小一致且与预测值(约30 kDa)相近,检测发现样品1,2,3均有褐藻胶裂解酶活性,而样品4(空载)无酶活性,说明重组大肠杆菌正确表达了褐藻胶裂解酶基因alg1987。
2.3.2 诱导剂浓度
将重组菌在IPTG终浓度分别为0、0.2 mmol/L、0.4mmol/L、0.6 mmol/L、0.8 mmol/L、1.0 mmol/L条件下下于16℃诱导16 h,收集菌体,超声波破碎细胞,离心后得到的上清液中褐藻胶裂解酶粗酶活性结果见图7。由图7可知,当诱导剂浓度介于0~0.2mmol/L时,不同IPTG浓度下破胞液上清中检出的粗酶活性较为稳定,酶活性较低,约为最高值的45%;随着IPTG浓度的升高,重组菌的褐藻胶裂解酶活性也呈上升趋势,并在浓度为0.8 mmol/L时达到最大;当IPTG浓度高于0.8 mmol/L时,酶活性迅速下降,推测0.8 mmol/L可能为该重组菌耐受的IPTG临界浓度。
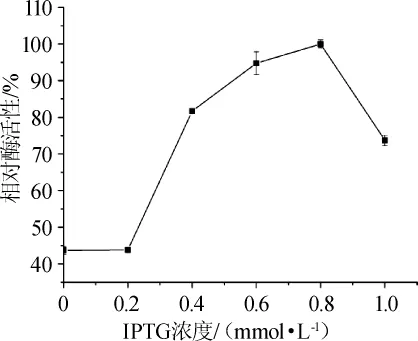
图7 IPTG浓度对表达的影响Fig.7 Effect of IPTG concentration on expression
综合上述的优化结果,选择诱导温度16℃、诱导时间16 h、诱导剂浓度为0.8 mmol/L为重组菌的表达褐藻胶裂解酶的最佳诱导条件。
3 结论
褐藻胶及其寡糖具有很大的应用潜力,但目前从自然环境中筛选到的大多数产褐藻胶裂解酶菌株的酶活性较低,产物复杂,分离纯化难度大,生产成本高,不利于褐藻胶裂解酶的规模化应用。利用基因工程手段将褐藻胶裂解酶基因导入遗传背景清晰的工程菌株获得高产的重组菌,使褐藻胶裂解酶的工业化大规模生产成为可能。
以本实验室筛选的海洋弧菌X511的基因组DNA为模板,克隆得到了褐藻胶裂解酶基因alg1987;经NCBI数据库同源比,下载与Alg1987同源性较高的序列,构建了进化树,发现该酶同源性最高(79%)的蛋白属于褐藻胶裂解酶PL7家族,推测Alg1987可能为褐藻胶裂解酶PL7家族中的一个新型酶;采用生物信息学工具预测褐藻胶裂解酶Alg1987的分子质量为30 254.28 Da,等电点8.75,分子式为C1368H2100N364O401S6;选用PET-30(a)为载体构建重组质粒并转化大肠杆菌BL21,利用IPTG诱导重组菌的表达,SDSPAGE凝胶电⒕检测诱导后重组菌的破碎细胞液上清,显示异源表达的褐藻胶裂解酶Alg1987与预测的分子质量大小一致。优化表达条件以提高重组菌中褐藻胶裂解酶的表达效率,得到的最适条件为:诱导温度16℃,诱导时间16 h,诱导剂IPTG浓度0.8 mmol/L。本研究为菌株海洋弧菌Vibriosp.X511产褐藻胶裂解酶Alg1987的大量制备及酶学性质研究打下了基础。
[1]TAKEDA H,YONEYAMA F,KAWAI S,et al.Bioethanol production from marine biomass alginate by metabolically engineered bacteria[J].Energ Environ Sci,2011,4(7):2575-2581.
[2]RAVANAL M C,PEZOA-CONTE R,SCHOULTZ S V,et al.Comparison of different types of pretreatment and enzymatic saccharification of Macrocystis pyrifera,for the production of biofuel[J].Algal Res,2016,13:141-147.
[3]THOMAS F,LUNDQVIST L C,JAM M,et al.Comparative characterization of two marine alginate lyases fromZobellia galactanivoransreveals distinct modes of action and exquisite adaptation to their natural substrate[J].J Biol Chem,2013,288(32):23021-23037.
[4]WONG T Y,PRESTON L A,SCHILLER N L.ALGINATE LYASE:review of major sources and enzyme characteristics,structure-function analysis,biological roles,and applications[J].Annu Rev Microbiol,2000,54(1):289-340.
[5]LINKER A,JONES R S.A new polysaccharide resembling alginic acid isolated from pseudomonads[J].J Biol Chem,1966,241(16):3845-3851.[6]周燕霞.褐藻胶裂解酶分泌菌株的分离鉴定及Tamlana holothuriorum s12T中褐藻胶裂解酶的研究[D].济南:山东大学,2016.
[7]GORIN P A J,SPENCER J F T.EXOCELLULAR ALGINIC ACID FROM AZOTOBACTER VINELANDII[J].Can J Chem,1966,44(9):993-998.
[8]CASWELL R C,GACESA P,LUTRELL K E,et al.MolecuLar cloning and heterologous expression of aKlebsiella pneumoniaegene encoding alginate lyase[J].Gene,1989,75(1):127-134.
[9]CHATER P I,WILCOX M D,BROWNLEE I A,et al.Alginate as a protease inhibitorin vitroand in a model gut system;selective inhibition of pepsin but not trypsin[J].Carbohyd Polym,2015,131:142-151.
[10]HOUGHTON D,WILCOX M D,CHATER P I,et al.Biological activity of alginate and its effect on pancreatic lipase inhibition as a potential treatment for obesity[J].Food Hydrocolloid,2015,49(11):18-24.
[11]SCHAEFFER D J,KRYLOV V S.Anti-HIV activity of extracts and compounds from algae and cyanobacteria[J].Ecotox Environ Safe,2000,45(3):208-227.
[12]REMMINGHORST U,REHM B H.Bacterial alginates:from biosynthesis to applications[J].Biotechnol Lett,2006,28(21):1701-1712.
[13]YANAKI T,NISHII K,TABATA K,et al.Ultrasonic degradation of Schizophyllum communepolysaccharide in dilute aqueous solution[J].J Appl Polym Sci,2010,28(2):873-878.
[14]WANG D M,KIM H T,YUN E J,et al.Optimal production of 4-deoxy-L-erythro-5-hexoseuLoseuronicacidfromalginate for brown macro algae saccharification by combining endo-and exo-type alginate lyases[J].Bioproc Biosyst Eng,2014,37(10):2105-2111.
[15]ASPINALL G O.The Polysaccharides[M].New York:Academic Press,1982:25-30.
[16]TANG J C,TANIGUCHI H,CHU H,et al.Isolation and characterization of alginate-degrading bacteria for disposal of seaweed wastes[J].Lett Appl Microbiol,2009,48(1):38-43.
[17]FARRELL E K,TIPTON P A.Functional characterization of AlgL,an alginate lyase fromPseudomonas aeruginosa[J].Biochemistry,2012,51(51):10259-66.
[18]WONG T Y,PRESTON L A,SCHILLER N L.Alginate lyase:review of major sources and enzyme characteristics,structure-function analysis,biologicalroles,andapplications[J].Annul Rev Microbiol,2000,54(1):289-340.
[19]YOON H J,HASHIMOTO W,MIYAKE O,et al.Overexpression in Escherichia coli,purification,and characterization ofSphingomoassp.A1 alginate lyases[J].Protein Expres Purif,2000,19(1):84-90.
[20]HASHIMOTO W,MIYAKE O,MOMMA K,et al.Molecular identification of oligoalginate lyase ofSphingomonassp.strain A1 as one of the enzymes required for complete depolymerization of alginate[J].J Bacteriol,2000,182(16):4572-4577.
[21]MIYAKE O,HASHIMOTO W,MURATA K.An exotype alginate lyase inSphingomonassp.A1:overexpressioninEscherichiacoli,purification,and characterization of alginate lyase IV (A1-IV)[J].Protein Expres Purif,2000,19(1):84-90.
[22]MURATA K,KAWAI S,MIKAMI B,et al.Superchannel of bacteria:Biologicalsignificanceandnewhorizons[J].Biosci Biotechnol Biochem,2008,72(2):265-677.
[23]TAKASE R,OCHIAI A,MIKAMI B,et al.Molecular identification of unsaturated uronate reductase prerequisite for alginate metabolism in Sphingomonassp.A1[J].BBA Protein Proteom,2010,1804(9):1925-1936.
[24]SHINABARGER D,MAY T B,BOYD A,et al.Nucleotide sequence and expression of thePseudomonas aeruginosaalgF gene controlling acetylation of alginate[J].Mol Microbiol,1993,9(5):1027-1035.
[25]PECIÑA A,PASCUAL A,PANEQUE A.Cloning and expression of the algL gene,encoding theAzotobacter chroococcumalginate lyase:purification,and characterization of the enzyme[J].J Bacteriol,1999,181(5):1409-1414.
[26]刘 航,曹海龙,岳 敏,等.褐藻胶裂解酶基因的克隆、表达载体构建以及表达条件的研究[J].华中师范大学学报:自然科学版,2012,46(4):456-460.
[27]王 莹,张 兰,王 亚,等.海洋弧菌QY105中褐藻胶寡糖酶OalA的基因克隆、重组表达与性质研究[J].中国海洋药物,2016,35(4):47-52.
[28]TAKEDA H,YONEYAMA F,KAWAI S,et al.Bioethanol production from marine biomass alginate by metabolically engineered bacteria[J].Energ Environ Sci,2011,4(7):2575-2581.
[29]WARGACKI A J,LEONARD E,WIN M N,et al.An engineered microbial platform for direct biofuel production from brown macroalgae.[J].Science,2012,335(6066):308-313.
[30]ENQUISTNEWMAN M,FAUST A M E,BRAVO D D,et al.Efficient ethanol production from brown macroalgae sugars by a synthetic yeast platform[J].Nature,2014,505(7482):239-243.
[31]韩 伟,林 娟,谢 永,等.褐藻胶裂解酶基因的克隆表达及重组酶酶学性质[J].微生物学通报,2017,44(5):1074-1080.
Conditions optimization of gene cloning and expression of alginate lyase
XU Chao1,2,HUANG Guiyuan1,WANG Qiaozhen1,LU Mingqian1,ZHANG Rongcan3,LIAO Wei4,ZHANG Yunkai2*,HUANG Shushi1
(1.Guangxi Key Laboratory of Marine Natural Products and Combinatorial Biosynthesis Chemistry,Guangxi Academy of Sciences,Nanning 530007,China;2.College of Life Science and Technology,Guangxi University,Nanning 530003,China;3.Guangxi Beibu Gulf Marine Research Centre,Nanning 530007,China;4.Guangxi Vocational and Technical College,Nanning 530226,China)
The alginate lyase gene in marineVibriosp.X511 was expressed byEscherichia colito realize the large-scale preparation of alginate lyase and research enzymatic properties.By analyzing the sequence,the first 20 amino acids of the enzyme were predicted as the signal peptide,the protein molecular mass without the signal peptide was 30 254.28 Da,the isoelectric point was 8.75,the molecular formula was C1368H2100N364O401S6,and the corresponding gene sequence was named asalg1987.The construction program of bacteria recombination was as follows:the specific nucleic acid primers were designed and thealg1987was obtained by PCR;the recombinant plasmid pET-30(a)-alg1987was introduced intoE.coliTrans5α for sequencing;the positive plasmid pET-30(a)-alg1987was extracted and introduced intoE.coliBL21,then expressed by the induce of IPTG.The supernatant of recombinant bacteria cytosol was induced and detected by SDS-PAGE,it was found that the molecular mass of the alginate lyase by heterologous expression was similar to the predicted value.The expression conditions of alginate lyase in recombinant bacteria were optimized.The results showed that optimum parameters were as follows:induction temperature 16℃,time 16 h,induction agent IPTG concentration 0.8 mmol/L.
alginate lyase;gene;cloning;conditions optimization;expression
Q814
0254-5071(2017)12-0120-06
10.11882/j.issn.0254-5071.2017.12.025
2017-09-13
国家自然科学基金(31560017);广西自然科学基金重点项目(2014GXNSFDA118012);广西科技计划重点研发项目(桂科AB16380071);广西科技计划科技基地和人才专项(桂科AD17129019)
许 超(1991-),男,硕士研究生,研究方向为海洋微生物。
*通讯作者:张云开(1972-),男,副研究员,博士,研究方向为微生物工程。
