基于系统发育的燕山东麓植物群落的构建机制
2017-12-22唐丽丽陈国平冯小梅赵铁建石福臣
唐丽丽 陈国平 冯小梅 赵铁建 石福臣*
(1.南开大学生命科学学院,天津 300071; 2.天津八仙山国家级自然保护区管理局,天津 301900)
基于系统发育的燕山东麓植物群落的构建机制
唐丽丽1陈国平1冯小梅2赵铁建2石福臣1*
(1.南开大学生命科学学院,天津 300071;2.天津八仙山国家级自然保护区管理局,天津 301900)
群落构建机制对解释植物物种共存、多样性的维持与发展至关重要。本文以燕山山脉东麓的植物群落为研究对象,野外调查了374个植物样方(20 m×30 m)。通过多元回归树、植物群落亲缘关系指数、高斯核函数密度估计等研究,重点探究了植物群落构建的机制。研究结果表明,燕山东麓的植物群落可划分为以荆条(Vitexnegundovar.heterophylla)、槲栎(Quercusaliena)、蒙古栎(Quercusmongolica)、栓皮栎(Quercusvariabilis)和白蜡树(Fraxinuschinensis)为优势的5个群系。植物最大树高这一功能性状表现出了较强的系统发育信号,此区域植物功能性状保守性较强。荆条灌丛、槲栎、蒙古栎群系的群落构建过程以环境过滤为主导,栓皮栎群系的群落构建过程以负密度制约为主导,白蜡树群落的构建机制有由环境过滤向负密度制约过渡的趋势。另外,本文引用高斯核密度分布估计对亲缘关系指数的分布进行估计,直观、真实地反映了亲缘关系指数的个体值及整体趋势,为探讨群落构建的生态位机制和中性理论间的争论提供更直接的证据。
燕山东麓;多元回归树(MRT);系统发育;高斯核密度估计(Gaussian KDE);群落构建机制
群落构建机制(community assembly rule),即群落物种多样性形成及其维持机制一直是群落生态学研究的核心问题。尤其是在当下全球生境遭到破坏的背景下,理解植物群落是如何构建、什么动力在驱动着植物的多样性及群落功能的变化,对全球植物资源的管理与修复有着至关重要的作用[1]。目前对群落构建机制有两种较为流行的解释分别为经典的生态位机制(niche-assembly)[2~5]和Hubbell提出的中性理论(neutral processes)[6],其中生态位机制包括环境过滤(environmental filtering)[3,5]和负密度制约(competitive interactions)[4~5]。生态位机制和中性理论间的争论由来已久,近年来,得益于基因测序成本的下降、测序技术的标准化、植物基因数据库的壮大、分子生物学、生物信息学及计算机等学科的发展,Webb等[7]提出用谱系结构来解释群落构建过程,并提出用亲缘指数来度量谱系结构,此后越来越多的基于谱系发育得出的关于动物、植物群落构建过程的研究结果也表明了此法用于解释群落构建机制的适宜性及合理性[8~14]。群落生态学与植物谱系发育概念的结合,为解释群落的构建提供了新的理念[15]。
该理念利用净亲缘指数(net relatedness index,NRI)和最近亲缘指数(nearest taxon index,NTI)来计算物种间的系统发育距离[7],并在功能性状保守和趋同两种情况下去讨论群落的构建机制[13~14]。若NTI、NRI>0,说明物种在系统发育结构上聚集,群落由亲缘关系比较密切、性状比较接近的物种构成;若NTI、NRI<0,说明物种系统发育结构发散,群落由亲缘关系较远的物种构成;若NTI、NRI=0,说明物种在系统发育结构上呈现随机状态[7]。在植物功能性状保守的前提下,环境过滤作用会导致群落谱系结构的聚集,负密度制约作用会导致群落谱系结构的发散。在植物功能性状趋同的情况下,环境过滤作用会导致群落谱系结构发散,负密度制约作用会导致群落谱系结构随机或聚集[7,13~14]。而功能性状保守或趋同,在中性作用下,群落谱系结构都呈随机状态。植物功能性状的保守与趋同,则通过植物功能性状的系统发育信号的强弱来反映,系统发育信号目前广泛应用Blomberg等[16]提出的K值法进行检验[17~18]。谱系发育相关理论的引入,为验证中性理论,也为解决两个理论之间的争议提供了一个新的途径[19~20]。
燕山山脉南北衔于大兴安岭、长白山和太行山,横亘于我国东北与华北之间,呈东西走向,气候和植物群落构成均兼具东北与华北地区特征。位于燕山东麓的八仙山国家级自然保护区保存有大面积完整的森林群落,为研究暖温带北部落叶阔叶林形成和演化提供了理想场所。目前有众多植物学、生态学等方面的研究在此开展,对植物群落的研究包括植被的分类、植物多样性指数的计算、群落结构的分析、森林凋落物土壤养分等[21~25]。此前的研究对植被分类、群落结构特征都有论述,但植被分类仅考虑植物本身,并未结合生境信息,而且对植物群落的分析仅仅集中于物种组成、群落外貌等内容,群落的构建机制不明。鉴于此,为明确此地植物群落构建过程,本文对燕山山脉东麓的植物展开了大规模的调查,在对结果进行分析的基础上,得出相应结论。
1 研究区域概况
本文的研究区域以八仙山国家级自然保护区为中心,向四周山体扩散,此区域属于燕山山脉东侧尾支,位于北京、天津、唐山及承德4市腹地,属于中生代“燕山构造运动”隆起的褶皱山体。此地属于暖温带季风性大陆气候区,雨热同季,年均气温8~10℃,年均降水量700~800 mm,降水多集中于7、8月份。水系发达,多以主峰“聚仙峰”为中心,呈放射状汇入淋河,最终注入于桥水库。地形复杂多样,地势高低起伏,900 m以上的山峰有19余座,主峰“聚仙峰”海拔1 052 m。中、高山地带的土壤多为山地棕壤,土层深厚,富含腐殖质;低山地带的土壤以淋溶褐土为主,土层较瘠薄,多砂砾[23~25]。该地植被处于暖温带植被到温带植被的交错过渡地带,森林植被以暖温带落叶阔叶林为主,植物种类多样,群落结构完整[21~22]。
2 研究方法
2.1 野外调查
植物调查采用样方法,初步辨认不同森林植物群落类型,选取有代表性的地段设置样方,样方多集中于八仙山,并在其邻近的山体散落设置。样方设置方法参照方精云等[26]的植物群落清查的技术规范,森林样方为20 m×30 m,森林乔木层样方内对角位置设置2个10 m×10 m灌木层样方,每乔木层样方内至少选取3个1 m×1 m草本层样方。用手持式智能OREGON 450型GPS并结合HOLUX mo241轨迹仪记录经度和纬度,用手持式海拔气压计并结合当地标准点海拔值矫正后记录海拔高程,用Showa Sokki公司生产的手罗盘测定样方坡度、坡向,对样地群落及主要组成植物进行拍照。乔木层记录树木种名、胸径、树高及健康状况;灌木层记录各种的种名、基径、平均高、株数及盖度;草本层记录种名、盖度、平均高及多度。
本研究中共设置样方374个,其中森林样方(乔、灌、草三层)361个,灌木样方(灌、草两层)13个。根据样地的经度和纬度等信息,通过ArcGIS10.1软件提取样地精确的海拔、坡度、坡向数据,样方的海拔为97~987 m,样方的坡度为2.068°~36.975°,坡向为8.303°~316.562°。
2.2 统计分析
2.2.1 群落数量分类—多元回归树
本文采用多元回归树的方法,对所调查样方的植物进行分类。多元回归树(multivariate regression trees,MRT)是De’Ath[27]于2002年提出,赖江山等[28]于2010年引进于植物群落的群丛分类的方法。此方法以物种—样方信息为因变量,以环境—样方信息为自变量,以自变量的某一最佳值为分类节点,将样方划分为尽可能同质的类别,重复这一过程,直至不能分割。此后将得到众多分类枝杈的结果通过交叉验证来剪枝,作者可以根据自己的实验目的,在“最小的误差±一个相应标准差”的范围内,选择最小的树作为划分结果。通过弄岗山热带喀斯特季节性雨林的研究结果可知,在草本层稀少的森林群落中,用木本植物进行分类的结果更加理想[29]。鉴于此经验,本文利用胸径≥1 cm的101种木本植物的物种信息,及样方的海拔、坡度、坡向信息,对调查的374个样方进行划分归类,并选择了最小的回归树。划分过程在R语言的“mvpart”程序包下进行。
群落命名参照赖江山于2010年提出的指示值法[28],利用植物的多度信息及各群系的样方数量,于R语言的“labdsv”程序包下进行计算,每个群落以指示值最大的植物进行命名,指示值最大的物种不仅在该群落中密度较高,且分布均匀,可以反映所在群落的特征,可以用此物种对群落进行命名。
2.2.2 系统发育信号
按照Blomberg等[16]提出的K值法,选取样方内木本植物的最大树高作为功能性状,进行研究区域内植物系统发育信号地检测。首先计算系统发育树末端物种性状数据的均方误差(MSE0)与基于系统发育树的方差—协方差矩阵(variance-covariance matrix)计算的均方误差(MSE)的比值,然后实际的MSE0/MSE相对于其期望值(基于现有谱系树的拓朴结构和枝长,并假设性状以布朗模型方式进化)的比值即为K值。K<1,则物种表现出适应性进化趋势;K=1,物种则随机进化;K>1,说明物种有显著的系统发育信号,功能性状保守。但系统发育信号在不同群落中的表现不同。K值的计算是在R语言的“ape”和“picante”程序包中计算完成。
2.2.3 构建谱系树
本文参考Webb等[7]的实验方法,构建谱系树。Webb等[7]于2002年首次尝试将谱系树运用于群落生态学的研究,于2005年建立phylomatic平台[30]。多年来,Webb及相关专家多次对相关理论及平台进行完善,目前谱系树理念和phylomatic平台已经广泛应用于群落生态学研究当中,相关的研究在世界范围内得以积极迅速的开展[31~35]。本文采用此方法,将调查的全部种子植物,按照科/属/种的格式,输入到phylomatic平台,生成基于APG Ⅲ分类系统具有进化枝长的系统发育树框架[32]。并在R语言的“picante”程序包中将此框架进行图形化。
2.2.4 亲缘关系指数计算
净亲缘指数(NRI)是指群落内实际得到的所有物种平均成对系统发育距离(MPDobs)相对于零模型(null model)随机值(MPDnull)的标准化效应值(standardized effect size)[7]。
最近亲缘指数(NTI)是指群落内亲缘关系最近的物种之间的平均系统发育距离(MNTDobs)与零模型随机生成值(MNTDnull)的标准化效应值[7]。
本文将植物在谱系树上的枝长等信息,结合各样方物种信息,在R语言的“picante”程序包下,计算得出各样方的NRI、NTI值。
2.2.5 各群落亲缘关系指数分布估计
本文根据多元回归树的分类结果,以各群落内样方的NRI、NTI值对该群落内亲缘关系指数分布进行估计。采用高斯核密度估计方法(Gaussian kernel density estimation)绘制各群落亲缘关系指数密度分布曲线,并估算NRI、NTI在大于0和小于0区域内的面积,以此对各群落亲缘关系指数的正负趋势进行判断,进而探究各群落谱系结构聚集、发散还是随机。此计算过程在R语言的“ggplot2”和“zoo”程序包下完成。
3 结果与分析
3.1 植物群落的数量分类及命名
3.1.1 群落划分依据
根据聚类结果,将调查的样方共分为5个群系(图1)。按照如果群系A的样方被群系B的若干样方包围,则可判断该样方归属于群系B的标准[28],对分类结果进行校正,结果表明群系Ⅰ样方有13个,群系Ⅱ的样方为29个,群系Ⅲ的样方为100个,群系Ⅳ的样方为108个,群系Ⅴ的样方为124个。海拔、坡度、坡向均是群落分类的节点,但海拔是群落分类的决定性因素。

图1 植物群落数量与分类——多元回归树Fig.1 Multivariate regression tree in the number and classification of phytoecommunity
3.1.2 植物群落的命名
按照指示值法对划分的5个群系进行命名(表1),依次为:
Ⅰ为荆条灌丛,包括13个样方,海拔均在475 m以下,并位于不大于7.002°的平稳缓坡上。灌木层的主要优势种为荆条,指示值为100,伴生有胡枝子(Lespedezabicolor)、扁担杆(Grewiabiloba)、小叶鼠李(Rhamnusparvifolia)、酸枣(Ziziphusjujubavar.spinosa)、雀儿舌头(Leptopuschinensis)等;草本层主要优势种为北京隐子草(Cleistogeneshancei)、矮丛苔草(Carexcallitrichosvar.nana),伴生种有大丁草(Gerberaanandria)、油芒(Spodiopogoncotulifer)、稗(Echinochloacrusgalli)、葎草(Humulusscandens)、狼尾草(Pennisetumalopecuroides)、委陵菜(Potentillachinensis)、益母草(Leonurusjaponicus)等。
Ⅱ为栓皮栎群系,包括29个样方,海拔在475 m以下,坡度在7.002°以上。该群系乔木层主要优势种为栓皮栎,指示值为62.1,伴生种有辽东栎(Quercuswutaishanica)、胡桃楸(Juglansmandshurica)、槲树(Quercusdentata)、油松(Pinustabuliformis)等;灌木层主要优势种为荆条,伴生种有扁担杆、小花溲疏(Deutziaparviflora)、小叶鼠李、三裂绣线菊(Spiraeatrilobata)等;草本层主要优势种为求米草(Oplismenusundulatifolius),伴生种有矮丛苔草、薯蓣(Dioscoreapolystachya)、秋苦荬菜(Ixeridiumsonchifolium)、香茶菜(Isodonamethystoides)、羊须草(Carexcallitrichos)、茜草(Rubiacordifolia)等。
Ⅲ为白蜡树群系,包括100个样方,多位于海拔475 m以上的北坡。乔木层主要优势种为白蜡,指示值为61.6,伴生有蒙古栎、紫椴(Tiliaamurensis)、中国黄花柳(Salixsinica)、油松、元宝槭(Acertruncatum)、五角枫(Acerpictumsubsp.mono)、大果榆(Ulmusmacrocarpa)等;灌木层主要优势种为三裂绣线菊,伴生有锦带花(Weigelaflorida)、大花溲疏(Deutziagrandiflora)、小叶鼠李、迎红杜鹃(Rhododendronmucronulatum)等,草本层主要优势种为矮丛苔草,伴生种有三脉紫菀(Asterageratoides)、银背风毛菊(Saussureanivea)、穿龙薯蓣(Dioscoreanipponica)等。
Ⅳ为槲栎群系,包含108个样方,多位于475~597.5 m的山坡。该群系乔木层主要优势种为槲栎,指示值为63.8,伴生有白蜡、鹅耳枥(Carpinusturczaninowii)、栓皮栎、胡桃楸、紫椴、栾树(Koelreuteriapaniculata)、油松等;灌木层主要优势种为小花溲疏,伴生种主要有雀儿舌头、鼠李(Rhamnusdavurica)、卫矛(Euonymusalatus)等;草本层主要优势种为求米草,伴生种主要有大叶铁线莲(Clematisheracleifolia)、矮丛苔草、羊须草、香茶菜等。
Ⅴ为蒙古栎群系,包括124个样方,多位于海拔597.5 m以上的山体。该群系乔木层主要优势种有蒙古栎,指示值为54.4,伴生种主要有油松、臭檀(Tetradiumdaniellii)、紫椴、胡桃楸、元宝槭等;灌木层主要优势种为小花溲疏,伴生有大花溲疏、蚂蚱腿子(Myripnoisdioica)、迎红杜鹃等,草本层主要优势种为大叶铁线莲,伴生有臭草(Melicascabrosa)、糙苏(Phlomisumbrosa)、矮丛苔草、歪头菜等。
表15个群系内优势物种指数值
Table1Theindicatorvalueofspeciesinfivevegetationformations
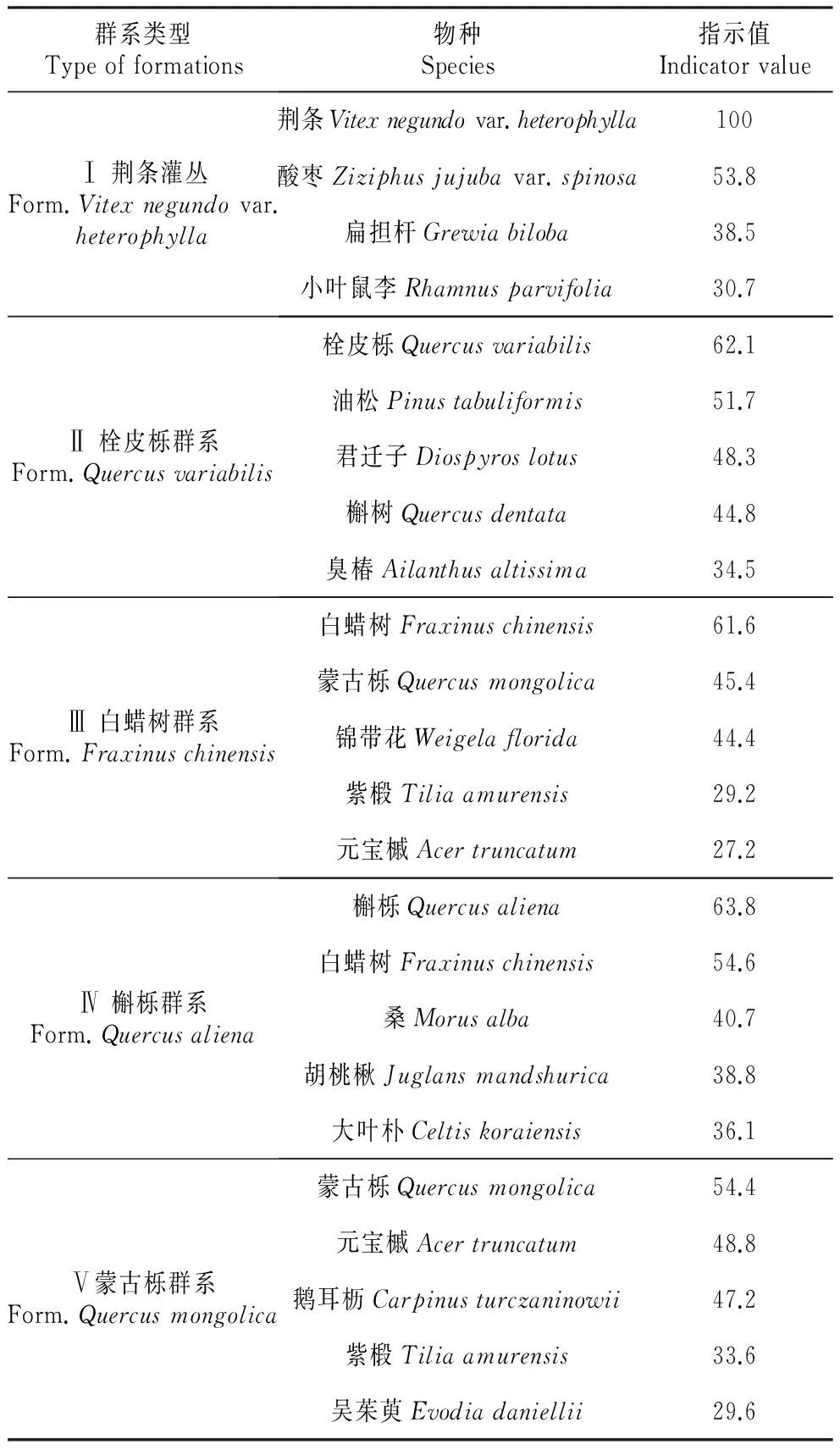
群系类型Typeofformations物种Species指示值IndicatorvalueⅠ荆条灌丛Form.Vitexnegundovar.heterophylla荆条Vitexnegundovar.heterophylla100酸枣Ziziphusjujubavar.spinosa53.8扁担杆Grewiabiloba38.5小叶鼠李Rhamnusparvifolia30.7Ⅱ栓皮栎群系Form.Quercusvariabilis栓皮栎Quercusvariabilis62.1油松Pinustabuliformis51.7君迁子Diospyroslotus48.3槲树Quercusdentata44.8臭椿Ailanthusaltissima34.5Ⅲ白蜡树群系Form.Fraxinuschinensis白蜡树Fraxinuschinensis61.6蒙古栎Quercusmongolica45.4锦带花Weigelaflorida44.4紫椴Tiliaamurensis29.2元宝槭Acertruncatum27.2Ⅳ槲栎群系Form.Quercusaliena槲栎Quercusaliena63.8白蜡树Fraxinuschinensis54.6桑Morusalba40.7胡桃楸Juglansmandshurica38.8大叶朴Celtiskoraiensis36.1Ⅴ蒙古栎群系Form.Quercusmongolica蒙古栎Quercusmongolica54.4元宝槭Acertruncatum48.8鹅耳枥Carpinusturczaninowii47.2紫椴Tiliaamurensis33.6吴茱萸Evodiadaniellii29.6
3.2 系统发育信号
最大树高是反映植物吸收和利用光资源最重要的功能性状,因此我们选择利用最大树高这一功能性状来分析。经计算结果得知最大树高这一功能性状表现出了显著的系统发育信号(K=1.70,P=0.001),说明在此区域最大树高这一功能性状上,系统发育保守性较强,功能性状与物种的进化历史有着密切的关系,亲缘关系较近的物种间性状的相似性更高。
3.3 群落谱系结构及构建机制分析
根据亲缘关系指数分布估计的结果(表2,图2~3)可知,本文所采用的高斯核密度分布估计方法,直观、全面的反映出各群系NRI、NTI值正负的趋势。各群系的NRI、NTI正负趋势如下:
美国次级制裁措施中有很多是针对向受制裁者名单上所列伊朗主体(包括伊朗国家石油公司及其分支机构)提供的某些支持。
荆条灌丛(Ⅰ)、槲栎群系(Ⅳ)、蒙古栎群系(Ⅴ)中,NRI、NTI均表现出正值趋势,即谱系结构聚集,荆条灌丛的构建机制可认为是环境过滤。但荆条灌丛(Ⅰ)在NRI值的水平上正负的面积相差不大;槲栎群系(Ⅳ)的NRI、NTI值在正负范围内的面积相差不大。
栓皮栎群系(Ⅱ)中,NRI、NTI值均表现出负值趋势,即谱系结构发散,栓皮栎群系的构建机制可认为是负密度制约。但在NTI水平上,其正负面积相差不大。
白蜡树群系(Ⅲ)的NRI值表现出正值趋势,谱系结构聚集;但其NTI值表现出了明显的负值趋势,由此得出谱系结构发散的结果。且在NRI水平上,正负面积相差不大。
另外,理论上NRI、NTI在正值、负值两个区域内面积的估算结果之和应为1,但由于此结果是通过估算而得,且本文想利用的是其正、负趋势,估计面积时的误差对亲缘关系指数的正、负趋势影响不大,所以可以接受表2结果中NRI、NTI在正、负范围内的面积和不等于1。
表25个群系NRI、NTI值在正负范围内的面积
Table2PositiveandnegativeareaundercurveofNRIandNTIinfivevegetationformations

NRINTI<0>0<0>0荆条灌丛Form.V.negundovar.heterophylla0.40033230.59806090.1192990.880119栓皮栎群系Form.Q.variabilis0.65269220.34632340.54034020.4586158白蜡树群系Form.F.chinensis0.45455370.54429630.6963390.302834槲栎群系Form.Q.aliena0.44646750.55193070.43029750.5687292蒙古栎群系Form.Q.mongolica0.21622950.78303230.3800650.6191014

图2 NRI在5个群系的分布Fig.2 NRI Distribution in five vegetation formations
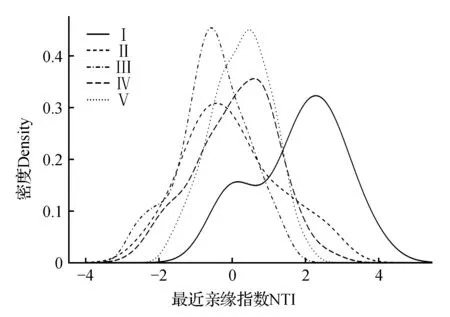
图3 NTI在5个群系的分布Fig.3 NTI distribution in five formations
4 讨论
4.1 基于多元回归树的群落数量分类
相较于传统的重要值法对群落的划分,本文依据多元回归树进行的植物群落数量分类,结合了植物与环境信息的多元回归树法划分的群落类型,更能简单直观的反映此地植被类型种类及其分布规律,且能据此得出此地植被分异的主要驱动因子为海拔。分类更加直观,可操作性、可利用性更强。
自De’Ath提出多元回归树可用于生态学相关领域的研究后,国外众多学者采用此法对荒漠、海滨滩涂、湿地植物进行了相关研究[36~40]。赖江山等[28]于2010年首次将此法引进于群落植物学的群丛分类,至此,国内学者也就此对法古田山、弄岗山、小秦岭、古尔班通古特沙漠等地植物群落的进行了群丛分类,从他们的研究中可知,海拔、坡度、坡向、纬度等因子的变化是区域植被分异的主要原因[28~29,41~42]。因此,本文结合海拔、坡度、坡向等环境因子,重新对涵盖八仙山国家级自然保护区在内的燕山山脉东麓的植物进行了分类,得到的分类结果可直观、简洁的反映研究区域植被的实际情况。
另外,由植物群落分类的节点可知,燕山山脉东麓的植被差异主要来源于海拔的变化,坡度和坡向也是植被分异的影响因素,但影响程度弱于海拔。虽然植物根据海拔、坡度、坡向的变化有所分异,但各群系的物种组成不存在明显界线,并不是截然不同的,而是存在很多共同种,特别是在森林群落当中,植物组成上几乎没有区别,有所区别的只是它们的优势程度。
4.2 高斯核密度估计方法在解释群落谱系结构中的应用
本文采用高斯核密度估计的方法对亲缘关系指数个体值及整体趋势进行表达,直接、有效的避免了在基于系统发育方法判断群落构建机制过程中假中性理论现象的产生。
本研究中各群系的亲缘关系指数虽有正负区别,但荆条灌丛(群系Ⅰ)、栓皮栎群系(群系Ⅱ)、白蜡树群系(群系Ⅲ)和槲栎群系(群系Ⅳ)的亲缘关系指数在正、负区域的面积均有趋近于相等的状况(荆条灌丛、白蜡树群系的NRI,栓皮栎群系的NTI,槲栎群系的NRI和NTI)。若正、负区域内面积趋近于相等,即NRI、NTI值的整体趋势趋近于0,则群落构建过程应判定为中性理论。而本文通过高斯核密度估计法做的亲缘关系指数分布曲线,保留了NRI、NTI的个体值,通过曲线的波峰分布可知,之所以会出现正负区域面积接近相等的情况,是因为群系Ⅰ、Ⅲ、Ⅳ的NRI值和群系Ⅱ的NTI值在正、负值区域内都出现了明显的波峰,也就是此群系中相当一部分的样方的NRI、NTI值均表现为正值,也有一部分样方的NRI、NTI值均表现为负值,即不是所有样方的NRI、NTI值为0,也就证实了并不是中性理论在各群系构建过程中起作用。
4.3 群落构建机制
荆条灌丛(Ⅰ)、槲栎群系(Ⅳ)、蒙古栎群系(Ⅴ)在群落构建过程中环境过滤起主导作用,栓皮栎群系(Ⅱ)中群落构建过程主要受负密度制约影响,在白蜡树群系(Ⅲ)中环境过滤和负密度制约共同影响着群落的构建过程。
荆条灌丛(Ⅰ)、槲栎群系(Ⅳ)、蒙古栎群系(Ⅴ)的谱系结构均表现为聚集,其NRI、NTI均大于0,尤其是蒙古栎群系的NRI、NTI值大于0的趋势更明显,表现出了更为强烈的聚集。由此说明这三个群系的构建机制为环境过滤,即在物种组成的过程中,环境对物种的筛选起主导作用。荆条灌丛(Ⅰ)多分布于低山的缓坡,槲栎群系(Ⅳ)、蒙古栎群系(Ⅴ)则多分布于海拔较高、坡向偏阳的山体。其分布区域阳光、土壤有机质含量都较为丰富[23~25],能满足亲缘关系较近的物种对相同资源的需求,因此由于环境的过滤导致了亲缘关系较近的物种聚集分布,表现出谱系结构聚集的现象。
栓皮栎群系(Ⅱ)的谱系结构较为发散(NRI、NTI均小于0),群系的构建机制表现为负密度制约,即栓皮栎群系在构建过程中,物种间竞争排斥较为激烈。此地栓皮栎群系分布的区域土层一般较为瘠薄,土壤有机质含量较少[23~25],植物可利用的资源有限。尤其是亲缘关系较近的物种,对资源的需求也较为相近[49~50]。为获取有限的资源,亲缘关系较近的物种间的相互排斥、相互竞争会尤为强烈,因此在栓皮栎群系构建过程中出现负密度制约现象较为合理。
在白蜡树群系(Ⅲ)中出现了NRI趋近于正值,谱系结构较为聚集,但其NTI趋近于负值,谱系结构表现发散的现象。NRI度量的是整个群落物种间系统发育的差异,而NTI度量的是亲缘关系相近的物种间系统发育的差异,即在白蜡树群系(Ⅲ)中,虽然整体上谱系关系较为相近,但亲缘关系较近的物种并不倾向于聚集在一起,而是散落分布于整个群系当中,因此白蜡树群系(Ⅲ)在物种形成过程中很有可能在初期经历了环境过滤,后期经历了负密度制约。即在物种形成初期,环境过滤起主导作用,使得亲缘关系较近的物种生长于此,但白蜡树群系(Ⅲ)位于此地山体的北坡,光照、土壤有机质含量等资源稍差于南坡[23~25],随着物种数量的增加,资源总量有限,物种间竞争越来越激烈,排斥现象加剧,亲缘关系最近的物种无法再聚集生长,便分散于整个群系当中,由此出现了NRI虽呈大于0的趋势,但NTI却明显小于0。
5 结论
在本研究中,燕山山脉东麓的植物群落可以分为以荆条、栓皮栎、白蜡树、槲栎和蒙古栎为优势的5个群系,海拔是此地植被分异最主要的影响因子。植物最大树高这一性状的系统发育信号较强,功能性状保守性较强。
本文采用高斯核密度分布估计方法对样方的NRI、NTI值进行的解读,更直观、形象、具体的反映了NRI、NTI的个体值的分布趋势;曲线面积的计算,也为判定NRI、NTI的整体正负趋势,提供了更有力的手段。个体及整体水平的NRI、NTI值的同时表达,为探讨群落构建中的生态位机制理论和中性理论的争论提供了直观证据。
荆条灌丛、槲栎群系和蒙古栎群系的谱系结构聚集,栓皮栎的谱系结构发散。白蜡树群系整体的谱系结构趋近于聚集,但亲缘关系相近的物种谱系结构发散。说明荆条灌丛、槲栎群系和蒙古栎群系的群落构建机制是环境过滤,栓皮栎群系的群落构建机制是负密度制约,而白蜡树群系在群落构建过程中可能经历了环境过滤到负密度制约的过程。
1.Cavender-bares J,Kozak K H,Fine P V A,et al.The merging of community ecology and phylogenetic biology[J].Ecology Letters,2009,12(7):693-715.
2.Diamond J M.Assembly of species communities[M].//Cody M L,Diamond J M.Ecology and evolution of communities.Cambridge:Harvard University Press,1975.
3.Weiher E,Keddy P.Ecological assembly rules:perspectives,advances,retreats[M].Cambridge:Cambridge University Press,1999.
4.Hubbell S P.The unified neutral theory of biodiversity and biogeography[M].Princeton,Oxford:Princeton University Press,2001.
5.Bazzaz F A.Habitat selection in plants[J].The American Naturalist,1991,137:S116-S130.
6.Tilman D.Resource competition and community structure[M].Princeton:Princeton University Press,1982.
7.Webb C O,Ackerly D D,Mcpeek M A,et al.Phylogenies and community ecology[J].Annual Review of Ecology & Systematics,2002,33:475-505.
8.Cavender-bares J,Ackerly D D,Baum D A,et al.Phylogenetic overdispersion in Floridian oak communities[J].The American Naturalist,2004,163(6):823-843.
9.Gillespie R.Community assembly through adaptive radiation in Hawaiian spiders[J].Science,2004,303(5656):356-359.
10.Fine P V A,Miller Z J,Mesones I,et al.The growth-defense trade-off and habitat specialization by plants in Amazonian forests[J].Ecology,2006,87(sp7):S150-S162.
11.Strauss S Y,Webb C O,Salamin N.Exotic taxa less related to native species are more invasive[J].Proceedings of the National Academy of Sciences of the United States of America,2006,103(15):5841-5845.
12.Davies J T,Meiri S,Barraclough T G,et al.Species co-existence and character divergence across carnivores[J].Ecology Letters,2007,10(2):146-152.
13.Kraft N J B,Cornwell W K,Webb C O,et al.Trait evolution,community assembly,and the phylogenetic structure of ecological communities[J].The American Naturalist,2007,170(2):271-283.
14.Vamosi S M,Heard S B,Vamosi J C,et al.Emerging patterns in the comparative analysis of phylogenetic community structure[J].Molecular Ecology,2009,18(4):572-592.
15.Mcpeek M A.The macroevolutionary consequences of ecological differences among species[J].Palaeontology,2007,50(1):111-129.
16.Blomberg S P,Garland JR T,Ives A R.Testing for phylogenetic signal in comparative data:behavioral traits are more labile[J].Evolution,2003,57(4):717-745.
17.曹科,饶米德,余建中,等.古田山木本植物功能性状的系统发育信号及其对群落结构的影响[J].生物多样性,2013,21(5):564-571.
Cao K,Rao M D,Yu J Z,et al.The phylogenetic signal of functional traits and their effects on community structure in an evergreen broad-leaved forest[J] .Biodiversity Science,2013,21(5):564-571.
18.宫骁.基于群落系统发育对沿坡向梯度上亚高寒草甸群落构建的分析[D].甘肃:兰州大学,2016.
Gong X.The research of community assembly along a slope aspect gradient in an alpine meadow based on community phylogeny[D].Gansu:Lanzhou University,2016.
19.Cavender-bares J,Pahlich A.Molecular,morphological,and ecological niche differentiation of sympatric sister oak species,QuercusvirginianaandQ.geminata(Fagaceae)[J].American Journal of Botany,2009,96(9):1690-1702.
20.牛红玉,王峥峰,练琚愉,等.群落构建研究的新进展:进化和生态相结合的群落谱系结构研究[J].生物多样性,2011,19(3):275-283.
Niu H Y,Wang Z F,Lian J Y,et al.New progress in community assembly:community phylogenetic structure combining evolution and ecology[J].Biodiversity Science,2011,19(3):275-283.
21.丛明旸,石会平,张小锟,等.八仙山国家级自然保护区典型森林群落结构及物种多样性研究[J].南开大学学报:自然科学版,2013,46(4):44-52.
Cong M Y,Shi H P,Zhang X K,et al.Analyses on community structure and species diversity of typical forest in Baxianshan national natural reserve[J].Acta Scientiarum Naturalium Universitatis Nankaiensis,2013,46(4):44-52.
22.王天罡.天津八仙山自然保护区植物多样性及其保护研究[D].北京:北京林业大学,2007.
Wang T G.Studies on plant diversity and conservation in Tianjin Baxianshan nature reserve[D].Beijing:Beijing Forestry University,2007.
23.陈国平,俎丽红,高张莹,等.八仙山不同立地落叶阔叶林凋落物养分特征及土壤肥力评价研究[J].植物研究,2016,36(6):878-885.
Chen G P,Zu L H,Gao Z Y,et al.The characteristics of forest floor nutrients and soil fertility assessment for deciduous broad-leaved forest with different site conditions[J].Bulletin of Botanical Research,2016,36(6):878-885.
24.陈国平,程珊珊,丛明旸,等.三种阔叶林凋落物对下层土壤养分的影响[J].生态学杂志,2014,33(4):874-879.
Chen G P,Cheng S S,Cong M Y,et al.Effects of litter decomposition on soil nutrients in three broad-leaved forests[J].Chinese Journal of Ecology,2014,33(4):874-879.
25.陈国平.天津典型森林群落和湿地群落对土壤生态特征的影响[D].天津:南开大学,2014.
Chen G P.Effects of typical forest communities and wetland communities on soil ecological characteristics in Tianjin[D].Tianjin:Nankai University,2014.
26.方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533-548.
Fang J Y,Wang X P,Shen Z H,et al.Methods and protocols for plant community inventory[J].Biodiversity Science,2009,17(6):533-548.
27.De’ath G.Multivariate Regression Trees:a new technique for modeling species-environment relationships[J].Ecology,2002,83(4):1105-1117.
28.赖江山,米湘成,任海保,等.基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例[J].植物生态学报,2010,34(7):761-769.
Lai J S,Mi X C,Ren H B,et al.Numerical classification of associations in subtropical evergreen broad-leaved forest based on multivariate regression trees-a case study of 24 hm2Gutianshan forest plot in China[J].Chinese Journal of Plant Ecology,2010,34(7):761-769.
29.黄甫昭,王斌,丁涛,等.弄岗北热带喀斯特季节性雨林群丛数量分类及与环境的关系[J].生物多样性,2014,22(2):157-166.
Huang P Z,Wang B,Ding T,et al.Numerical classification of associations in a northern tropical karst seasonal rain forest and the relationships of these associations with environmental factors[J].Biodiversity Science,2014,22(2):157-166.
30.Webb C O,Donoghue M J.Phylomatic:tree assembly for applied phylogenetics[J].Molecular Ecology Notes,2005,5(1):181-183.
31.Bremer B,Bremer K,Chase M W,et al.An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants:APG III[J].Botanical Journal of the Linnean Society,2003,141(4):399-436.
32.Zanne A E,Tank D C,Cornwell W K,et al.Three keys to the radiation of angiosperms into freezing environments[J].Nature,2014,506(7486):89-92.
33.Webb C O,Ackerly D D,Kembel S W.Phylocom:software for the analysis of phylogenetic community structure and trait evolution[J].Bioinformatics,2008,24(18):2098-2100.
34.Kress W J,Erickson D L,Jones F A,et al.Plant DNA barcodes and a community phylogeny of a tropical forest dynamics plot in Panama[J].Proceedings of the National Academy of Sciences of the United States of America,2009,106(44):18621-18626.
35.Swenson N G.The assembly of tropical tree communities-the advances and shortcomings of phylogenetic and functional trait analyses[J].Ecography,2013,36(3):264-276.
36.Zhang R,Liu T,Zhang J L,et al.Spatial and environmental determinants of plant species diversity in a temperate desert[J].Journal of Plant Ecology,2016,9(2):124-131.
37.Arieira J,Penha J,Da Cunha C N,et al.Ontogenetic shifts in habitat-association of tree species in a neotropical wetland[J].Plant and Soil,2016,404(1):219-236.
38.Obolewski K,Gotkiewicz W,Strzelczak A,et al.Influence of anthropogenic transformations of river bed on plant and macrozoobenthos communities[J].Environmental Monitoring and Assessment,2011,173(1-4):747-763.
39.Sommer B,Froend R.Phreatophytic vegetation responses to groundwater depth in a drying Mediterranean-type landscape[J].Journal of Vegetation Science,2014,25(4):1045-1055.
40.Gapare W J,Ivkovic M,Liepe K J,et al.Drivers of genotype by environment interaction in radiata pine as indicated by multivariate regression trees[J].Forest Ecology and Management,2015,353:21-29.
41.陈云,王海亮,韩军旺,等.小秦岭森林群落数量分类、排序及多样性垂直格局[J].生态学报,2014,34(8):2068-2075.
Chen Y,Wang H L,Han J W,et al.Numerical classification,ordination and species diversity along elevation gradients of the forest community in Xiaoqinling[J].Acta Ecologica Sinica,2014,34(8):2068-2075.
42.张荣,刘彤.古尔班通古特沙漠南部植物多样性及群落分类[J].生态学报,2012,32(19):6056-6066.
Zhang R,Liu T.Plant species diversity and community classification in the southern Gurbantunggut Desert[J].Acta Ecologica Sinica,2012,32(19):6056-6066.
43.Gastauer M,Meira-neto J A.Interactions,Environmental sorting and chance:phylostructure of a tropical forest assembly[J].Folia Geobotanica,2014,49(3):443-459.
44.Piwczyński M,Puchaka R,Ulrich W.Influence of tree plantations on the phylogenetic structure of understorey plant communities[J].Forest Ecology and Management,2016,376:231-237.
45.Liu X H,Zhu X X,Niu Y,et al.Phylogenetic clustering and over-dispersion for alpine plants along elevational gradient in the Hengduan Mountains Region,southwest China[J].Journal of Systematics and Evolution,2014,52(3):280-288.
46.Freilich M A,Connolly S R.Phylogenetic community structure when competition and environmental filtering determine abundances[J].Global Ecology and Biogeography,2015,24(12):1390-1400.
47.Martins C A,Roque F O,Santos B A,et al.Correction:what shapes the phylogenetic structure of anuran communities in a seasonal environment? the influence of determinism at regional scale to stochasticity or antagonistic forces at local scale[J].PLoS One,2016,11(3):e0151734.
48.Brunbjerg A K,Cavender-bares J,Eiserhardt W L,et al.Multi-scale phylogenetic structure in coastal dune plant communities across the globe[J].Journal of Plant Ecology,2014,7(2):101-114.
49.童鑫.从种群遗传和群落组成的空间结构研究群落维持机制[D].上海:华东师范大学,2015.
Tong X.Exploring community assembly through the lens of spatial structure:from population genetics to community composition[D].Shanghai:East China Normal University,2015.
50.Tilman D,Wedin D,Knops J.Productivity and sustainability influenced by biodiversity in grassland ecosystems[J].Nature,1996,379(6567):718-720.
National Science and Technology Basic Project of the Ministry of Science and Technology of China(2011FY110300);The Project of Wild Plant Resource Inventory of Tianjin(20120024002)
introduction:TANG Li-Li(1989—),female,Ph.D,major in plant resource and plant ecology.
date:2017-04-29
CommunityAssemblyRulesoftheEastofYanshanMountainBasedonPhylogeny
TANG Li-Li1CHEN Guo-Ping1FENG Xiao-Mei2ZHAO Tie-Jian2SHI Fu-Chen1*
(1.College of Life Science,Nankai University,Tianjin 300071; 2.Baixian Mountian National Nature Reserves,Tianjin 301900)
Analysis of community assembly rules plays a crucial role in understanding species coexistence and the maintenance of biodiversity of the community. We conducted field investigations and statistical analysis on 374 plant plots(20 m×30 m) in the east of Yanshan Mountain, and used Multiple Regression Tree(MRT), phylogenetic index and Gaussian kernel density estimation(Gaussian KDE) on data collected from study sites to explore its community assembly rules. The whole community could be divided into five formations, includingVitexnegundovar.heterophylla,Quercusaliena,Q.mongolica,Q.variabilisandFraxinuschinensis. Among these forms, environmental filtering mainly influenced the community assembly process inV.negundovar.heterophylla,Q.alienaandQ.mongolica, competition controlled the speciation inQ.variabilis, while inF.chinensis, the pattern had been changing from environmental filtering to competition. In addition, Gaussian KDE was introduced to illustrate net relatedness index(NRI) and nearest taxon index(NTI) values, providing a new intuitive perspective to explore the argument between niche-assembly and neutral processes.
the east of Yanshan Mountain;Multiple Regression Tree(MRT);phylogeny;Gaussian kernel density estimation(Gaussian KDE);community assembly rules
国家科技部基础性工作专项(2011FY110300);天津市野生资源植物调查项目(20120024002)资助
唐丽丽(1989—),女,博士研究生,主要从事植物资源与植物生态学研究。
* 通信作者:E-mail:fcshi@nankai.edu.cn
2017-04-29
* Corresponding author:E-mail:fcshi@nankai.edu.cn
S718.54
A
10.7525/j.issn.1673-5102.2017.06.002
