景宁木兰(Magnolia sinostellata)远缘杂交授粉过程障碍研究
2017-12-22陈翔翔范李节王倩颖尹增芳申亚梅
陈翔翔 卢 璐 范李节 王倩颖 尹增芳 申亚梅*
(1.浙江农林大学风景园林与建筑学院,临安 311300; 2.南京林业大学生物与环境学院,南京 210037)
景宁木兰(Magnoliasinostellata)远缘杂交授粉过程障碍研究
陈翔翔1卢 璐1范李节1王倩颖1尹增芳2申亚梅1*
(1.浙江农林大学风景园林与建筑学院,临安 311300;2.南京林业大学生物与环境学院,南京 210037)
以景宁木兰(Magnoliasinostellata)为亲本,分别与望春玉兰(M.biondii)、紫玉兰(M.liliiflora)、红运二乔(M.soulangeana‘Red lucky’)、丹馨玉兰(M.soulangeana‘Danxin’)进行种间远缘杂交,并以景宁木兰自交为对照,研究了景宁木兰自交和杂交亲和性。结果表明:(1)景宁木兰具有较高的花粉活力与柱头可授性,花粉管能正常进入杂交母本花柱,自交结实率为2%;(2)以景宁木兰为父本,紫玉兰柱头对其花粉具有较好的识别性,花粉萌发率和花粉管生长速度均高于以红运二乔和丹馨玉兰做母本的组合,花粉管生长较为正常,结实率为25%,而与红运玉兰杂交的结实率为4%,与丹馨玉兰杂交的结实率为0%;(3)以景宁木兰为母本,望春玉兰花粉在其柱头上的萌发比率高于其他4个组合,但花粉管生长过程中出现扭曲,景宁木兰花柱内出现胼胝质沉积,且最终并未获得种子。花粉管发育镜检结果表明:受精前障碍即花粉管进入花柱基部前生长异常是引起景宁木兰与丹馨玉兰、望春玉兰杂交不结实的主要原因;并进一步推测花粉管进入子房后的异常状态是导致景宁木兰自交,与红运二乔杂交结实率低的主要原因。
景宁木兰;人工杂交;花粉萌发;结实率;杂交亲和性
景宁木兰(Magonliasinostellata)为木兰属(Magonlia)玉兰亚属1新种,目前仅在浙江有发现,因其分布范围狭窄、种群数量少、结实少,且种子萌发率低,处于极度濒危状态,2012年被列为浙江省保护植物[1~2]。在观赏特性方面,该物种为木兰属灌木类植物,花期3月,花瓣数量9~18枚不等,花初开时粉红色,盛花时近白色,具有较高的观赏价值。
目前对于木兰属内种间人工杂交已有报道,其中玉兰亚属内杂交多为亲和,并获得杂交优异后代,如二乔玉兰(M.soulangeana)系列品种等[3~4]。但在自然生境中,木兰属植物虽然有足够的花粉量,但不同物种花粉活力、萌发率均存在差异,最终影响了结实率,存在杂交障碍。因此为探讨景宁木兰与木兰科木兰属其他物种间的杂交亲和性,开发其育种潜力,本研究以其为亲本,分别与望春玉兰(M.biondii)、紫玉兰(M.liliiflora)、红运二乔(M.soulangeana‘Red lucky’)、丹馨玉兰(M.soulangeana‘Fragrant cloud’)进行杂交,对授粉过程进行观察,并统计结实率,研究景宁木兰与其他玉兰的杂交亲和性,同时为开发木兰科优异新种质提供基础理论依据。
1 材料与方法
1.1 试验材料
本试验于2013年2月底至4月以浙江安吉刘家塘林场长势良好且花量丰富的10年生景宁木兰及浙江农林大学校园内长势良好、正常开花的10年生望春玉兰、紫玉兰、红运二乔、丹馨玉兰为试验材料,分别于开花期进行杂交试验。
1.2 试验方法
各试验材料花期略有不同(望春玉兰2月底3月初,景宁木兰、丹馨玉兰、红运二乔玉兰3月,紫玉兰4月),且木兰科植物具有雌蕊先熟的特性。望春玉兰雄蕊与景宁木兰雌蕊成熟期一致,与丹馨玉兰、红运二乔可花期相遇,与紫玉兰花期不遇。因景宁木兰花量有限,本试验只设计了景宁木兰×望春玉兰、紫玉兰×景宁木兰、丹馨玉兰×景宁木兰、红运二乔×景宁木兰4个杂交组合,以景宁木兰自交作为对照(CK)。2013年3月,望春玉兰雄蕊与景宁木兰雌蕊均成熟时,选择晴朗天气收集新鲜的望春玉兰花粉对去雄后的景宁木兰进行授粉;同时在景宁木兰雄蕊成熟时,收集新鲜的景宁木兰花粉进行储藏,用于丹馨玉兰、红运二乔与紫玉兰雌蕊成熟时进行授粉,并在授粉后不同时间对花粉在柱头内生长情况进行荧光观察。
1.2.1 花粉采集、贮藏与活力测定
望春玉兰新鲜花粉采集后,直接进行活力测定,并进行授粉试验。景宁木兰花粉采集后,分摊开置于硫酸纸上,放入硅胶干燥器中干燥2 h后,收集到离心管或者冷冻管中并盖紧管盖,置于-80℃冰箱中进行保存。分别对未干燥新鲜花粉(A)、干燥2 h后新鲜花粉(B)、未干燥低温贮藏15 d后花粉(C)、干燥2 h低温贮藏15 d后花粉(D)的萌发率进行测定。萌发率测定采用离体萌发法[5]。把花粉管萌发长度超过花粉粒直径的花粉视为已萌发花粉,每次处理统计3个视野,每个视野花粉不少于100粒,取其平均值作为该处理的花粉萌发率[5],并用SPSS.19软件进行数据统计分析。
花粉萌发率=(萌发的花粉粒数目/观察总花粉粒数目)×100%
(1)
1.2.2 柱头形态观察与柱头可授性测定
本实验采用观察法和联苯胺—过氧化氢法测定柱头可授性[5]。采集开花后0、2、6、20、24、48、72 h的柱头,在凹槽载玻片的凹槽处滴入联苯胺—过氧化氢反应液,将柱头浸入其中。在光学显微镜下观察,如果反应液变蓝并伴有大量气泡产生,则柱头有较强的可授性;反应液变蓝,有少量气泡产生,表明柱头有可授性;反应液无明显变色反应且无气泡产生,则柱头可授性差。
1.2.3 杂交授粉及取样固定
在开花期选取母本发育良好的花蕾,在松蕾时进行去雄处理,即将内、外花被片和雄蕊全部去除(不伤及柱头),用毛笔蘸取收集好的花粉授于柱头上,每种杂交组合授粉54~81个柱头,授粉后及时套袋并挂牌记录。授粉后0.5、1、2、6、12、24、36、48、72 h各取下6~9个雌蕊,浸没于装有FAA(甲醛∶冰醋酸∶50%乙醇=1∶1∶8)固定液的5 mL离心管中,固定24 h以上,进行荧光镜检。
1.2.4 荧光镜检
将雌蕊用刀片轻轻从花托上剖开,用镊子取出雌蕊柱头及部分花柱,投入8 mol·L-1NaOH软化脱色2 h,然后用0.1%苯胺蓝溶液浸泡10 h染色,甘油压片,在荧光显微镜(Nikon Eclipse 80i)下观察柱头上的花粉附着与花粉萌发情况。每张切片取3个视野观察统计花粉管萌发率。
有花粉管的柱头比率=(花粉萌发柱头数/所观察花粉柱头数)×100%
(2)
花粉萌发率=(萌发形成花粉管的花粉数目/观察总花粉粒数目)×100%
(3)
1.2.5 杂交结实性测定
景宁木兰×望春玉兰、红运二乔×景宁木兰、丹馨玉兰×景宁木兰各组合分别授粉50朵;因紫玉兰花朵数较少,紫玉兰×景宁木兰组合授粉32朵。60 d后统计坐果率,90 d后统计结实率。每个组合3次重复。数据用Excel记录。
坐果率=(果实数量/授粉花朵数)×100%
(4)
结实率=(含有有胚种子的果实数/授粉花朵数)×100%
(5)
2 结果与分析
2.1 景宁木兰与望春玉兰花粉活力比较
图1显示,景宁木兰新鲜未干燥花粉萌发率为88.63%(图1:S-FP),干燥2 h后花粉萌发率为89.05%(图1:S-FP-2h),未干燥-80℃贮藏15 d后花粉萌发率为77.54%(图1:S-FP-15D),干燥2 h-80℃贮藏15 d后花粉萌发率为79.78%(图1:S-FP-2h-15D)。由此可见,花粉萌发率主要受储藏时间影响,储藏时间越长,萌发率越低。新鲜未干燥、新鲜干燥2 h后花粉活力明显高于-80℃冷藏15 d花粉的活力(P<0.01)。相同储藏时间条件下,干燥后花粉萌发率略高于未干燥花粉,且这种效果随储藏时间的延长表现更明显。经干燥2 h处理的新鲜花粉活力高于未干燥的新鲜花粉活力,但二者差异不显著;-80℃冷藏15 d后干燥处理花粉活力显著高于未干燥冷藏处理花粉活力(P<0.01)(图1:GR)。图1:B-FP显示,望春玉兰新鲜花粉活力81.74%,由此可见,景宁木兰与望春玉兰花粉活力均能满足授粉的要求。
2.2 柱头可授性检测
观察不同开花时期的柱头形态,发现从花苞开始膨大到花蕾完全打开,柱头形态均为向外向下弯曲,有明显的粘性分泌物,柱头表面均呈不同程度透明状,其中花蕾膨大到将要完全展开阶段最明显,推测该阶段柱头具有较高的可授性。到花瓣完全张开后1~2 d,柱头表面开始出现褐化现象。联苯胺—过氧化氢法测定结果显示(图2),景宁木兰、紫玉兰、红运二乔、丹馨玉兰在花蕾期至开放前期状态时,柱头有明显的粘性分泌物,并向外弯曲,反应液检测气泡量较大。
2.3 景宁木兰自交亲和性
荧光镜检结果显示(表1),景宁木兰开花初期,可观察到有花粉附着于柱头上(图3:A);12 h时可观察到花粉管生长,具有花粉萌发的柱头比率为80.0%(图3:B);到了24 h时可观察到花粉管已生长至花柱中,有花粉萌发的柱头比率为83.3%,花粉萌发率为100%(图3:C)。
2.4 景宁木兰(♀)×望春玉兰(♂)杂交的亲和性
望春玉兰新鲜花粉的活力为81.7%,极显著低于景宁木兰新鲜花粉活力(P<0.01),但极显著高于冷藏后的景宁木兰花粉活力(P<0.01),能满足授粉要求(图1)。荧光镜检结果显示(表1),授粉6 h后(图3:D)景宁木兰柱头上可观察到少量花粉萌发,且有花粉萌发的柱头比率为5%,而花粉萌发率为100%。12 h后有花粉萌发的柱头比率达到37%(图3:E),24 h后升至50%。且此时花粉萌发率达到88.33%,花粉管进一步伸长,但部分花粉管呈现扭曲(图3:F)。36 h时可观察到花粉管生长数量增加减少,但花粉管末端出现膨大、扭曲,周围出现胼胝质沉积(图3:G),此时有花粉萌发的柱头比率为63.3%。48 h时可观察到部分花粉管达到花柱,其余花粉管末端变形,甚至出现断裂,生长受到抑制(图3:H),有花粉管萌发的柱头比率为48.6%。
2.5 紫玉兰(♀)×景宁木兰(♂)杂交的亲和性
荧光镜检结果显示(表1),4 h时可观察到紫玉兰柱头上有少量景宁木兰花粉(图3:I),12 h后附着的花粉粒数量增加,但未见花粉萌发(图3:J)。24 h时,大量花粉萌发,花粉管向花柱基部方向伸长,其中有花粉萌发的柱头比率为11.7%,花粉萌发率为92.57%(图3:K),大部分花粉管生长形态正常,并能顺利进入花柱。授粉后36~72 h,花粉大量萌发,花粉管快速生长,到72 h时有花粉萌发的柱头比率达33.3%,并可观察到花粉管花伸向柱基部方向(图3:L-N)。

图1 不同处理下景宁木兰、望春玉兰花粉萌发率 S-FP.景宁木兰新鲜未干燥花粉萌发情况;S-FP-2h.景宁木兰新鲜花粉干燥2 h后花粉萌发情况;S-FP-15D.景宁木兰未干燥低温贮藏15 d后花粉萌发情况;S-FP-2h-15D.景宁木兰干燥2 h低温贮藏15 d后花粉萌发情况;B-FP.望春玉兰新鲜花粉的萌发情况;GR. 花粉萌发率,差异极显著P<0.01Fig.1 Pollen germination rate of M.sinostellata and M.biondii in vitro under different conditions S-FP. The germination of fresh pollen of M.sinostellata; S-FP-2h. The germination of fresh pollen which was drought for 2 h of M.sinostellata; S-FP-15D. The germination of fresh pollen was stored in -80℃ after 15 d of M.sinostellata; S-FP-15D. The germination of drought pollen was stored in -80℃ after 15 d of M.sinostellata; B-FP. The germination of fresh pollen of M.biondii; GR.Pollen germination rate(significant different was P<0.01)

图2 4个种质的母本柱头可授性 MS-1.景宁木兰花朵;MS-2.景宁木兰柱头可授性;ML-1.紫玉兰花朵,ML-2.紫玉兰柱头可授性;MR-1.红运二乔花朵;MR-2.红运二乔柱头可授性;MF-1.丹馨玉兰花朵;MF-2.丹馨玉兰柱头可授性Fig.2 Stigma receptivity of four species MS-1. Flower of M.sinostellata; MS-2. Stigma receptivity of M.sinostellata; ML-1. Flower of M.liliiflora; ML-2. Stigma receptivity of M.liliiflora; MR-1. Flower of M.soulangeana ‘Red lucky’; MR-2. Stigma receptivity of M.soulangeana ‘Red lucky’; MF-1. Flower of M.soulangeana ‘Danxin’; MF-2. Stigma receptivity of M.soulangeana ‘Fragrant cloud’
2.6 红运二乔(♀)×景宁木兰(♂)杂交的亲和性
荧光镜检结果显示(表1),授粉后12 h内,花粉开始粘附于柱头上(图4:A),随着时间延长,粘附的花粉量逐渐增加,但花粉并未萌发;授粉24 h后,观察到花粉管生长,有花粉萌发的柱头比率达18.5%,此时花粉萌发率为84.17%(图4:B);36 h后有花粉萌发的柱头比率达到25.8%(图4:C);授粉后48~72 h,花粉管进一步生长(有花粉萌发的柱头比率分别为29.4%和17.5%),但部分花粉管生长方向杂乱,花粉管末端出现膨大并扭曲(图4:D-F)。
2.7 丹馨玉兰(♀)×景宁木兰(♂)杂交的亲和性
荧光镜检结果显示(表1),丹馨玉兰做母本与景宁木兰杂交时,同样具有杂交亲和性。授粉后12 h多数柱头上未观察到有花粉粘附(图4:G),少量有花粉粘附的柱头上花粉也未萌发(图4:H);到授粉后24 h,花粉萌发率为81.76%,有花粉萌发的柱头比率为8.8%,并观察到花粉管生长,但只有部分花粉管伸向花柱方向,另一部分卷曲在柱头周围,出现异常生长(图4:I);在授粉36、48 h后,有花粉萌发的柱头比率依次为18.7%、13.1%,花粉管生长明显受到抑制(图4:J-K),到授粉72 h时,花粉管末端呈现尖刺状,未见其延伸到花柱基部方向(图4:L)。

表1 5个组合有花粉萌发的柱头比率与花粉萌发率
注:景宁木兰自交,景宁木兰×望春玉兰在6 h之前未见花粉萌发情况;红运玉兰×景宁木兰,丹馨玉兰×景宁木兰,紫玉兰×景宁木兰在12 h之前未见花粉萌发。
Note:No pollen germination was found inM.sinostellataself-hybridization,M.sinostellata×M.biondiibefore 6 h; No pollen germination was found inM.liliflora×M.sinostellata,M.soulangeana‘Red lucky’×M.sinostellata,M.soulangeana‘Fragrant cloud’×M.sinostellatabefore 12 h.
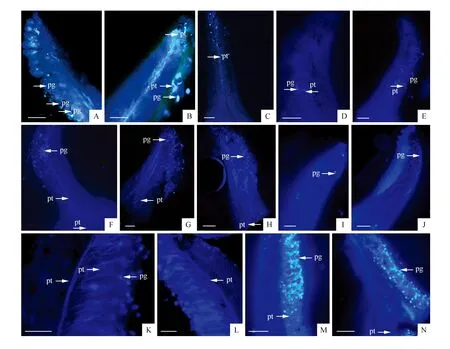
图3 花粉萌发与花粉管生长 pg.花粉粒;pt.花粉管 A-C依次为0、12、24 h景宁木兰自交时花粉管萌发情况;D-H依次为6、12、24、36、48 h望春玉兰花粉粒在景宁木兰柱头上花粉萌发与花粉管生长情况;I-N依次为6、12、24、36、48、72 h景宁木兰花粉粒在紫玉兰柱头上花粉萌发与花粉管生长情况。Fig.3 The germination of pollen and the growth of pollen tube pg.Pollen grains;pt.Pollen tube A-C.The pollen germination and pollen tube growth of M.sinostellata selfing at 0, 12, 24 h; D-H.The pollen germination and pollen tube growth of M.sinostellata×M.biondii at 6, 12, 24, 36, 48 h; I-N. The pollen germination and pollen tube growth of M.liliflora×M.sinostellata at 6, 12, 24, 36, 48, 72 h.

图4 花粉萌发与花粉管生长情况说明 pg.花粉粒;pt.花粉管 A-F依次为6、12、24、36、48、72 h景宁木兰花粉粒在红运二乔柱头上花粉萌发与花粉管生长情况;G-L依次为6、12、24、36、48、72 h景宁木兰花粉粒在丹馨玉兰柱头上花粉萌发与花粉管生长情况。Fig.4 The germination of pollen and the growth of pollen tube pg.Pollen grains;pt.Pollen tube A-F.The pollen germination and pollen tube growth of M.×soulangeana ‘Red lucky’×M.sinostellata at 6, 12, 24, 36, 48, 72 h; G-L. The pollen germination and pollen tube growth of M.‘Danxin’×M.sinostellata at 6, 12, 24, 36, 48, 72 h.
Table2Comparisonoffertilitybetweenfourcrossingcombinations

组合Crossingcombination授粉数Pollinationnumber坐果率Capsulesnumber(%)结实率Capsulesetting(%)种子数量(粒)Seedsnumber景宁木兰自交(CK)M.sinostellataselfing5020%2%5景宁木兰×望春玉兰M.sinostellata×M.biondii5010%0%0紫玉兰×景宁木兰M.liliflora×M.sinostellata3264%25%13红运玉兰×景宁木兰M.×soulangeana‘Redlucky’×M.sinostellata506%4%3丹馨玉兰×景宁木兰M.‘Fragrantcloud’×M.sinostellata506%0%0
2.8 杂交结实性比较
杂交结实性统计结果显示(表2),60 d后紫玉兰×景宁木兰组合坐果率最高,为64%,高于对照(CK)的20%,其余依次为景宁木兰×望春玉兰10%、红运玉兰×景宁木兰6%、丹馨玉兰×景宁木兰6%,且均低于对照CK。90 d后观察,4个组合结实率均很低,景宁木兰×望春玉兰与丹馨玉兰×景宁木兰组合未见种子,紫玉兰×景宁木兰组合收获13粒种子,红运玉兰×景宁木兰组合收获3粒种子,对照(CK)组收获5粒种子。
3 讨论
木兰科植物为雌雄异熟,在传统意义上被认为是为了避免自交的繁殖机制,但也有研究者认为这种现象可能属于较原始种类植物在性状上表现出的保守性[6]。其交配方式多为异交,部分自交亲和[7],如木兰科特有种(品种)(M.praecocissima‘borealis’)[8]、濒危物种星花木兰(M.Stellata)[9]、大果木莲(Manglietiagrandis)[10]、醉香含笑(Micheliamacclurei)[11]等。对以上种类进行人工自交,结果发现自交结实率均低于异交与常规自然授粉,且种子繁殖系数极低[8]。本研究结果中,景宁木兰也具有以上特征。在研究景宁木兰花粉活力与柱头可授性时发现,新鲜花粉、低温储藏、干燥处理之后的景宁木兰花粉活力与柱头可授性均能满足自交的要求,且花粉管正常萌发,并能进入花柱,但结实率低。由此可见,景宁木兰应存在受精后障碍。
远缘杂交的亲和性是反映物种之间的亲缘关系以及育种可能性的重要依据[12],亲缘关系较远物种间存在严重的生殖障碍而导致杂交往往难以成功[13~15]。在杂交过程中,母本的育性、父本花粉的活力是杂交获得高结实率的关键因素[16]。而在花粉具备活力的情况下,花粉粒与柱头、花粉管与花柱之间相互识别对杂交成功也有重要影响[17],相互间不能识别就会导致生殖障碍[18~19]。本研究中花粉活力与柱头可授性均满足了授粉的条件,各个柱头对所授花粉均能识别,但花粉萌发与花粉管生长情况存在差异。在授粉初期,仅景宁木兰×望春玉兰组合中花粉管开始萌发,其他组合均未见萌发;到24 h观察到其余组合花粉管萌发,且除丹馨玉兰×景宁木兰组合外,其余组合的多数花粉管都能生长到花柱位置。通常情况下,杂交亲和表现为花粉在柱头上最初萌发时间早,花粉管生长速度较快,到达花柱底部的花粉管比率较高;而花粉管形态出现不同程度的扭曲、末端膨大、末端刺状等,这些现象是导致杂交不亲和的主要原因[20]。本试验中,随着时间的延长,各个组合花粉管生长形态发生变化:望春玉兰花粉管生长过程中末端出现膨大、扭曲,且当花粉管进入花柱时,出现大量的胼胝质沉积,影响了花粉管的进一步生长,表现出较低的亲和性;丹馨玉兰柱头内,景宁木兰花粉管末端呈现刺状,未能进入花柱中,亲和性最低;紫玉兰×景宁木兰与红运玉兰×景宁木兰组合虽然花粉在柱头上萌发时间较晚,但是大部分花粉管生长形态正常,并能顺利到达花柱,与此同时花粉管萌发率高于其他组合。坐果率、结实率统计结果进一步表明,景宁木兰与紫玉兰亲和性最好,其次为红运二乔、望春玉兰,与丹馨玉兰的亲和力最低。以上结果也进一步验证了景宁木兰与紫玉兰亲缘关系较近[21],其与红运二乔表现出较好的亲和性,是因为该物种为紫玉兰与白玉兰的杂交后代,从遗传角度看,该物种特征可能更接近紫玉兰。
综上所述,景宁木兰自交亲和性很低。该物种的野生生境存在破碎化,种群极小,近交衰退也可能是导致其自身繁育障碍的主要原因。由此可见,景宁木兰与其他玉兰物种杂交不亲和,也可能是自身繁育系统原因导致杂交障碍。因此,有必要开展景宁木兰授粉后受精、胚胎发育等的系统研究,为景宁木兰的保护与育种开发提供重要理论基础。
1.刘饶,徐端妙,潘智慧,等.景宁木兰濒危原因初探[J].河北农业科学,2009,13(6):31-32.
Liu R,Xu D M,Pan Z H,et al.Analysis on the reasons of endangered ofMagnoliasinostellata[J].Journal of Hebei Agricultural Science,2009,13(6):31-32.
2.Wang Y L,Ejder E,Yang J F,et al.Magnoliasinostellataand relatives(Magnoliaceae)[J].Phytotaxa,2013,154(1):47-58.
3.龚洵,潘跃芝,杨志云.木兰科植物的杂交亲和性[J].云南植物研究,2001,23(3):339-344.
Gong X,Pan Y Z,Yang Z Y.The cross-compatibility of Magnoliaceae[J].Acta Botanica Yunnanica,2001,23(3):339-344.
4.王亚玲,李勇,张寿洲,等.木兰科植物的人工杂交[J].武汉植物学研究,2003,21(6):508-514.
Wang Y L,Li Y,Zhang S Z,et al.The crossing result of Magnoliaceae[J].Journal of Wuhan Botanical Research,2003,21(6):508-514.
5.卢璐,余泽智,刘雪燕,等.景宁木兰花粉萌发与贮藏特性研究[J].植物研究,2014,34(2):182-187.
Lu L,Yu Z Z,Liu X Y,et al.Pollen germination and storage ofMagnoliasinostellata[J].Bulletin of Botanical Research,2014,34(2):182-187.
6.张大勇.植物生活史进化与繁殖生态学[M].北京:科学出版社,2004.
Zhang D Y.Evolution of plant life history and reproductive ecology[M].Beijing:Science Press,2004.
7.Wang R H,Jia H,Wang J Z,et al.Flowering and pollination patterns ofMagnoliadenudatawith emphasis on anatomical changes in ovule and seed development[J].Flora-Morphology,Distribution,Functional Ecology of Plants,2010,205(4):259-265.
8.Ishida K.Beetle pollination ofMagnoliapraecocissimavar.borealis[J].Plant Species Biology,1996,11(2-3):199-206.
9.Hirayama K,Ishida K,Setsuko S,et al.Reduced seed production,inbreeding,and pollen shortage in a small population of a threatened tree,Magnoliastellata[J].Biological Conservation,2007,136(2):315-323.
10.付玉嫔,陈少瑜,吴涛.濒危植物大果木莲与中缅木莲的花部特征及繁育系统比较[J].东北林业大学学报,2010,38(4):6-10.
Fu Y P,Chen S Y,Wu T.Comparison of floral traits and breeding system of endangered plantManglietiagrandisand non-endangered plantManglietiahookeri[J].Journal of Northeast Forestry University,2010,38(4):6-10.
11.Zhao X F,Sun W B.Abnormalities in sexual development and pollinator limitation inMicheliacoriacea(Magnoliaceae),a critically endangered endemic to Southeast Yunnan,China[J].Flora-Morphology,Distribution,Functional Ecology of Plants,2009,204:463-470.
12.韩欣,成仿云,肖佳佳,等.以‘凤丹白’为母本的杂交及其育种潜力分析[J].北京林业大学学报,2014,36(4):121-125.
Han X,Cheng F Y,Xiao J J,et al.Crosses ofPaeoniaostii‘Feng Dan Bai’ as maternal parents and an analysis on the potential in tree peony breeding[J].Journal of Beijing Forestry University,2014,36(4):121-125.
13.Mohapatra A,Rout G R.Study of embryo rescue inFloribundarose[J].Plant Cell,Tissue and Organ Culture,2005,81(1):113-117.
14.Datson P M,Murray B G,Hammett K R W.Pollination systems,hybridization barriers and meiotic chromosome behaviour in Nemesia hybrids[J].Euphytica,2006,151(2):173-185.
15.Cheng F Y.Advances in the breeding of tree peonies and a cultivar system for the cultivar group[J].International Journal of Plant Breeding,2007,1(2):89-104.
16.彭晓莉,廖康,贾杨,等.9个新疆杏品种间杂交亲和性研究[J].果树学报,2015,32(2):192-199.
Peng X L,Liao K,Jia Y,et al.Study on the cross compatibility among 9 apricot cultivars in Xinjiang[J].Journal of Fruit Science,2015,32(2):192-199.
17.孙青春,陈发棣,房伟民,等.甘菊与栽培菊‘金陵黄玉’种间杂交失败的原因[J].园艺学报,2009,36(9):1333-1338.
Sun Q C,Chen F D,Fang W M,et al.Investigation on the factors leading to infertility in the cross betweenDendranthemalavandulifoliumandD.grandiflorum‘Jinling Huangyu’[J].Acta Horticulturae Sinica,2009,36(9):1333-1338.
18.Van Tuyl J M,Marcucci M C,Visser T.Pollen and pollination experiments.Ⅶ.The effect of pollen treatment and application method on incompatibility and incongruity inLilium[J].Euphytica,1982,31(3):613-619.
19.Sarmah B K,Sarla N.Overcoming prefertilization barriers in the crossDiplotaxissiettiana×Brassicajunceausing irradiated mentor pollen[J].Biologia Plantarum,1995,37(3):329-334.
20.耿兴敏,张超仪,尹增芳.部分野生杜鹃杂交授粉后花粉管生长状况分析[J].安徽大学学报:自然科学版,2014,38(4):94-101.
Geng X M,Zhang C Y,Yin Z F.Analysis of pollen tube growth after pollination in some wildRhododendron[J].Journal of Anhui University:Natural Science Edition,2014,38(4):94-101.
21.刘秀丽,胡陶,张启翔.16种玉兰亚属植物的AFLP分析[J].东北林业大学学报,2013,41(8):91-94.
Liu X L,Hu T,Zhang Q X.AFLP analysis for 16 species of SubgenusYulania[J].Journal of Northeast Forestry University,2013,41(8):91-94.
National Natural Science Foundation of China(31400599);the 12th Five-Year-Plan Projects for Floriculture Special Breeding of Zhejiang Province,China(2012C12909-4)
introduction:CHEN Xiang-Xiang(1992—),female,master whose major is application of garden plants.
date:2017-03-29
DistantHybridizationBarriersofMagnoliasinostellatainthePollinationProcess
CHEN Xiang-Xiang1LU Lu1FAN Li-Jie1WANG Qian-Ying1YIN Zeng-Fang2SHEN Ya-Mei1*
(1. School of Landscape Architecture,Zhejiang A&F University,Linan 311300; 2.College of Biology and the Environment,Nanjing Forestry University,Nanjing 210037)
With selfing ofMagnoliasinostellataas a control, distant crosses ofM.sinostellatawithM.biondii,M.liliiflora,M.soulangeana‘Red lucky’ andM.soulangeana‘Danxin’, respectively, were conducted to study the compatibility of selfing and that between species.M.sinostellatahad a relatively high pollen viability and stigma receptivity, and its pollen tube could go into the maternal style normally but had a selfing-based fruiting rate of 2%. The pollen fromM.sinostellatawas fast identified by the stigma ofM.liliifloraand germinated at a higher percentage with a faster growing normal tube than by those ofM.soulangeana‘Red lucky’ andM.soulangeana‘Danxin’.M.liliiflora×M.sinostellatahad a fruiting rate of 25% while the fruiting rate ofM.soulangeana‘Red lucky’×M.sinostellataandM.‘Fragrant cloud’×M.sinostellatawas only 4% and 0, respectively. WithM.sinostellataas a maternal parent, the pollen ofM.biondiihad a higher pollen germination rate than that of other three species, but the pollen tube twisted during growth and the callose was formed inside the style ofM.sinostellata, which resulted in no seed formation. All these indicated that pre-fertilization barriers, i.e. abnormal growth of pollen tube before reaching the bottom of the style, was the main reason for no fruiting in the cross ofM.sinostellatawith either ofM.soulangeana‘Danxin’ andM.biondii. The abnormal state of pollen tube after entering into the ovary might lead to a low fruiting rate from selfing ofM.sinostellataand hybridization ofM.sinostellatawithM.liliiflora, orM.soulangeana‘Red lucky’.
Magnoliasinostellata;artificial crossing;pollen germination;fruiting rate;crossing compatibility
国家自然科学基金资助项目(31400599);浙江省农业新品种选育重大科技专项(2012C12909-4)
陈翔翔(1992—),女,硕士研究生,主要从事园林植物应用研究。
* 通信作者:E-mail:yameishen@zafu.edu.cn
2017-03-29
* Corresponding author:E-mail:yameishen@zafu.edu.cn
S685.11
A
10.7525/j.issn.1673-5102.2017.06.003
