吉非替尼诱导p27核转位并激活半胱天冬酶8促进NSCLC细胞凋亡的机制
2017-12-22王新卫刘新年
杨 畅 冯 燕 王新卫 刘新年
(湖北省第三人民医院呼吸内科,湖北 武汉 430033)
吉非替尼诱导p27核转位并激活半胱天冬酶8促进NSCLC细胞凋亡的机制
杨 畅 冯 燕1王新卫 刘新年
(湖北省第三人民医院呼吸内科,湖北 武汉 430033)
目的探讨吉非替尼诱导的p27蛋白表达改变和核转位及半胱天冬酶(Caspase)8的激活对表皮生长因子受体(EGFR)突变的非小细胞肺癌(NSCLC)细胞周期阻滞和细胞凋亡的影响及作用机制。方法选取存在19号外显子缺失突变的对吉非替尼敏感的NSCLC细胞系HCC827,采用3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴盐(MTT)法和流式细胞术分别检测吉非替尼在不同浓度和时间下对细胞活性的影响。利用Western印迹检测p27蛋白和Caspase及其他相关蛋白的相对含量。通过免疫荧光法观察p27蛋白的亚细胞定位。采用蛋白免疫共沉淀验证p27蛋白和Caspase8之间的相互作用。结果吉非替尼浓度-时间依赖性促进了HCC827细胞凋亡。当吉非替尼干预18 h后,细胞周期蛋白依赖性激酶(CDK)抑制因子p27蛋白表达开始出现显著升高(P<0.05),且p27蛋白在此过程中伴随从细胞核到细胞质的核转位现象,而促凋亡蛋白B细胞淋巴瘤(Bcl)-2相关x蛋白(Bax)和Bad及抗凋亡蛋白Bcl-2和Bcl/白血病-x基因长片段(Bcl-xl)在吉非替尼干预后表达并未发生显著改变。此外,细胞质中p27蛋白与Caspase8的降解中间体p43/p41结合从而抑制其自身细胞质向细胞核易位。结论推断吉非替尼通过调节p27蛋白的亚细胞定位及Caspase8之间的相互作用,从而达到促EGFR突变的NSCLC细胞凋亡的效果。
吉非替尼;表皮生长因子受体;p27蛋白;半胱天冬酶8;凋亡;非小细胞肺癌
吉非替尼是一种可逆的、高选择性的酪氨酸激酶抑制剂,能与三磷酸腺苷竞争性结合表皮生长因子受体(EGFR)酪氨酸激酶活性区,从而抑制EGFR自身磷酸化并阻断其下游信号级联放大〔1,2〕。非小细胞肺癌(NSCLC)细胞系HCC827广泛存在19号外显子缺失和21号外显子点突变,已知这种激活突变能增强细胞对EGFR酪氨酸激酶抑制剂的敏感性〔3〕。吉非替尼对EGFR突变的NSCLC细胞可产生显著的促凋亡效果〔4〕,抑制NSCLC细胞的EGFR激酶活性能够促进细胞凋亡并将细胞周期抑制在G1期〔5〕。细胞周期进展受到细胞周期蛋白和细胞周期蛋白依赖性蛋白激酶(CDKs)的调控,而且细胞从G1期进入S期受到负性细胞周期调控因子——CDK抑制因子家族的调控。p27蛋白是主要靶向细胞周期蛋白E/CDK2复合体的CDK抑制因子。抑制CDK2活性可以将周期蛋白E/CDK2驱动的G1期向S期进展阻滞。此外,应用EGFR抑制剂干预后,肿瘤细胞中CDK抑制因子p27蛋白的表达水平显著升高〔6〕。细胞质p27蛋白在凋亡过程中与半胱天冬酶(Caspase)8裂解中间体p43/p41结合,可能是p27蛋白促细胞凋亡的关键〔7〕。本研究旨在探讨吉非替尼诱导的p27蛋白表达改变和核转位及Caspase8的激活对EGFR突变的NSCLC细胞周期阻滞和细胞凋亡的影响及作用机制。
1 材料和方法
1.1细胞培养 HCC827细胞购自美国模式菌种收集中心(ATCC)细胞库。细胞培养于含有10%胎牛血清(浙江天杭生物科技有限公司)和1%链霉素青霉素(北京索莱宝科技有限公司)的杜尔伯科极限必需培养基(DMEM)高糖培养基(美国HyClone公司),置于37℃,5% CO2培养箱(美国Thermo公司371型)培养。
1.2试剂 吉非替尼原料药,3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴盐(MTT)购于美国Sigma公司;膜联蛋白(V-FITC)细胞凋亡检测试剂盒(南京赛泓瑞生物科技有限公司);高灵敏度化学发光(eECL)检测试剂盒(北京康为世纪生物科技有限公司);抗Caspase8、9小鼠单克隆抗体,抗Caspase3兔多克隆抗体,抗p-EGFR和抗磷酸化蛋白激酶B(p-Akt)兔多克隆抗体均购于英国Abcam公司;抗p27和抗p21蛋白兔多克隆抗体,抗β-Actin兔多克隆抗体,抗(GAPDH)及抗核纤层蛋白(Lamin)B兔多克隆抗体购于美国ABGENT公司。抗B细胞淋巴瘤(Bcl)-2、抗Bcl/白血病-x基因长片段(Bcl-xl)、抗Bcl相关x蛋白(Bax)及抗Bad兔多克隆抗体购自武汉三鹰生物技术有限公司;异硫氰酸荧光素(FITC)共轭的抗兔和抗小鼠抗体购于美国BD公司;蛋白免疫共沉淀(蛋白A树脂)试剂盒(上海哈灵生物科技有限公司);亚细胞结构NE-PER 胞质和核蛋白抽提试剂盒(美国pierce公司)。
1.3细胞活力试验 采用MTT法检测细胞增殖和活力。取处于对数生长期的HCC827细胞,用血球计数板计数,用DMEM完全培养基将其稀释成1×104个/ml铺于96孔板,每孔100 μl。加入不同浓度(0.00、0.01、0.10、1.00、5.00、10.00 μmol/L)的吉非替尼处理72 h后,将MTT溶液加入使终浓度达到0.5 mg/ml,37℃培养4 h。弃去培养基后,每孔加入50 μl二甲基亚砜。利用590 nm波长酶标仪(美国Bio-rad公司imark型)检测每孔光密度值。不同浓度加药组与对照组吸光度的比值作为各组细胞相对活力,代表细胞死亡情况。
1.4流式细胞术 分别收集1 μmol/L吉非替尼处理不同时间(0、3、6、12、18、24 h)的HCC827细胞,细胞计数板计数,每组约1×106个细胞。洗涤,与Annexin V-FITC结合,冰浴30 min,随后加入1 μg/ml碘化丙啶(PI),置暗处10 min后采用流式细胞仪(美国BD公司FACSCalibur型)上机检测。Annexin V-FITC对凋亡细胞翻转到细胞膜外侧的磷脂酰丝氨酸具有较高亲和力,DNA染料PI进入凋亡细胞结合其DNA。根据二者所发荧光,仪器会自动分析出各组细胞的凋亡率。
1.5Western印迹 利用放射免疫沉淀法(RIPA)将细胞裂解进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE),随后将裂解蛋白电转于硝酸纤维素膜。将此膜用含5%脱脂奶粉和0.1%吐温20的磷酸盐缓冲液(T-PBS)室温封闭1 h,然后将一抗用T-PBS混合液以1∶1 000稀释比例4℃孵育过夜。待将硝酸纤维素膜用T-PBS冲洗3遍后用含辣根过氧化物酶标的1∶2 000稀释倍数的相应二抗室温孵育1 h。再次将硝酸纤维素膜洗涤数次后,滴加eECL化学发光试剂并进行暗室曝光。条带的灰度值用 Image J软件检测分析。
1.6免疫荧光分析 在培养板中将已爬好细胞的玻片用PBS浸洗后,将其置于含2%多聚甲醛的PBS 37℃下固定20 min。采用含0.2% Triton X-100的PBS室温通透30 min后,将细胞用PBS冲洗3次后于37℃进行一抗(抗p27,抗Caspase8)孵育1 h,一抗用PBS混合液(3%牛血清白蛋白,0.05%吐温20,0.08%叠氮化钠)稀释至1∶100的比例。细胞冲洗3次,每次5 min。细胞用1∶400稀释的共轭有FITC或德克萨斯红(Texas Red)的二抗于37℃下孵育30 min。将玻片置于含0.1 μg/ml Hoechst H33342荧光染料的PBS中浸泡。冲洗2遍后,细胞用共聚焦显微镜(德国NanoFocus公司μsurf explorer型)进行直接观察。
1.7蛋白免疫共沉淀 利用细胞质提取液进行免疫沉淀反应(IP)。待细胞生长融合至90%左右时将细胞培养瓶置于冰上,加入细胞质裂解液孵育1 h。4℃,1 000 r/min离心10 min,分离抽取上清液。取1 mg细胞裂解产物加入一抗于4℃过夜孵育。然后加入蛋白A-琼脂糖4℃振摇24 h。用洗涤缓冲液将沉淀洗涤5次,2倍SDS样品缓冲液将沉淀重悬后上样进行SDS-PAGE免疫印迹分析。
1.8统计学方法 应用SPSS18.0软件进行t检验、单因素方差分析、HSD-q检验。
2 结 果
2.1吉非替尼对HCC827细胞凋亡的影响 吉非替尼浓度依赖性对HCC827细胞的活力产生了显著抑制,其中0.00 μmol/L相对细胞活性为(100.0±5.4)%,0.01 μmol/L为(87.2±5.3)%,0.10 μmol/L为(49.0±4.9)%,1.00 μmol/L为(34.1±5.8)%,5.00 μmol/L为(32.7±5.9)%,10.00 μmol/L为(14.9±5.0)%(F=115.908,P=0.000)。流式细胞仪检测发现吉非替尼时间依赖性促进了HCC827细胞凋亡,其中0 h细胞凋亡率为1.0%,3 h为6.4%,6 h为12.7%,12 h为20.6%,18 h为33.9%,24 h为65.8%。
2.2吉非替尼通过激活Caspase8和促进p27蛋白表达诱导细胞凋亡 吉非替尼处理HCC827细胞24 h后检测到有活性的裂解Caspase8(p18)和Caspase3的18片段,与PARP降解的时间进程相似。紧接着有活性的Caspase8和Caspase8的中间降解产物p43及p41在吉非替尼处理18 h后同样被明显的检测出来。与此相反,能够启动细胞凋亡通路的Caspase9在这个过程中并未发现显著活化现象。促凋亡蛋白Bax和Bad及抗凋亡蛋白Bal-2和Bcl-xl的表达在吉非替尼干预后同样未发现有显著改变。当HCC827细胞分别接受吉非替尼处理0、3、6、18和24 h后,有效抑制了EGFR和Akt的磷酸化状态,同时在吉非替尼干预18 h后p27蛋白的表达出现显著上调(F=123.019,P=0.000)。其中0 h时p27蛋白灰度值为5.8±2.3,3 h为2.2±2.1,6 h为4.5±2.4,18 h为32.3±2.6,24 h为35.1±2.6。但是在同样情况下并未发现p21蛋白的表达增加,见图1。
2.3吉非替尼促进p27蛋白由胞核到胞质的转位 见图2,图3。随着处理时间的增加,p27蛋白含量在细胞质显著升高(0 h:52.5±4.7,3 h:53.5±6.3,6 h:48.3±6.1,18 h:80.5±6.8,24 h:105.2±8.6;P<0.05),但在细胞核中却随之降低(0 h:59.5±8.0,3 h:74.4±9.2,6 h:73.7±10.5,18 h:65.1±8.8,24 h:46.2±7.5)。随着吉非替尼处理时间的增加,p27蛋白在细胞核和细胞质中亚细胞定位与Western印迹分析结果均一一对应。

图1 吉非替尼通过激活Caspase8和提高p27蛋白表达水平诱导HCC827细胞凋亡
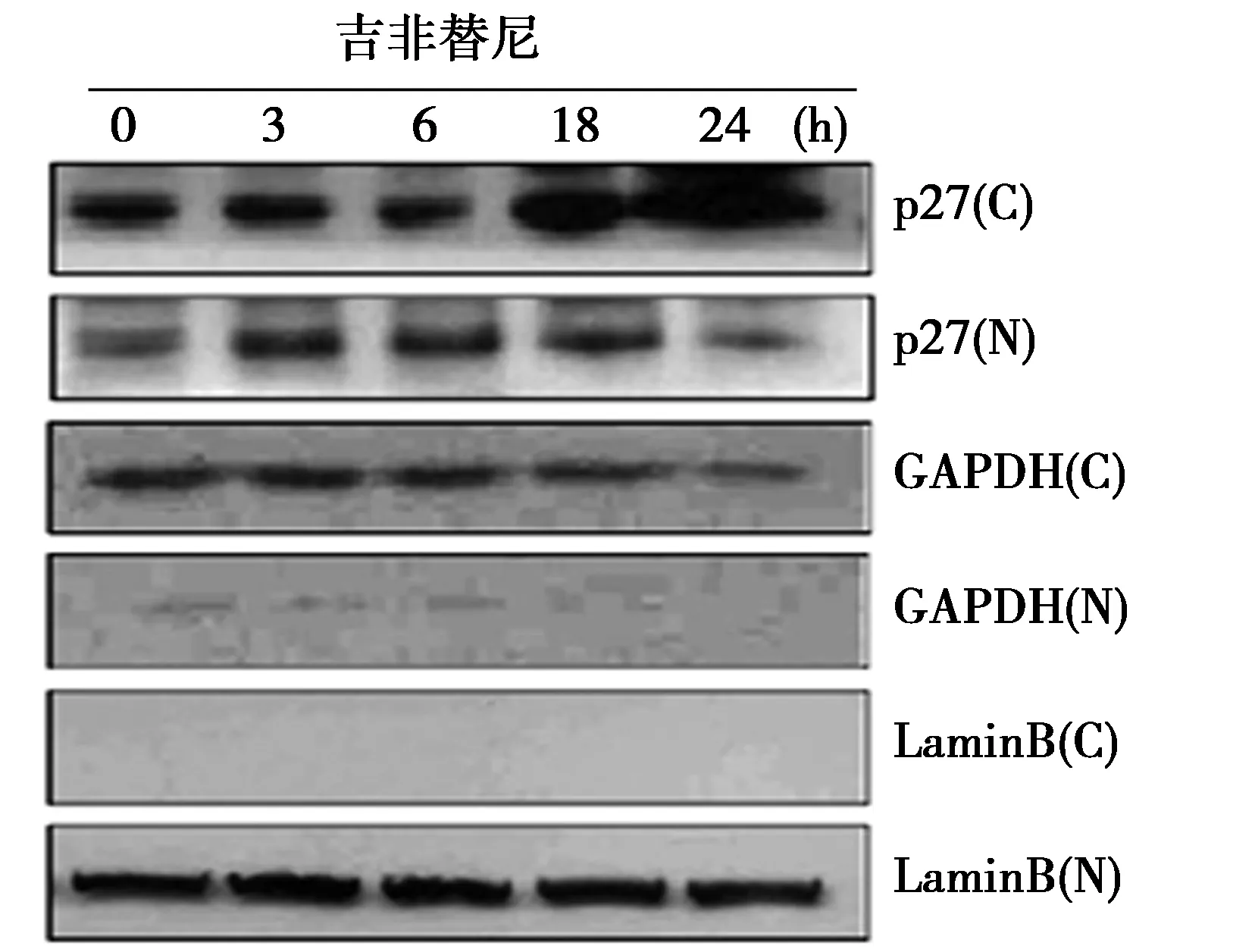
图2 Western印迹检测细胞质(C)和细胞核(N)裂解物中p27蛋白相对含量
2.4吉非替尼对细胞质中p27与Caspase8中间体p43/p41结合的影响 吉非替尼促进了细胞质中p27蛋白的表达和Caspase8的活化。吉非替尼处理24 h后,用抗p27蛋白抗体和抗Caspase8抗体对胞质蛋白进行免疫共沉淀反应。吉非替尼处理后检测到Caspase8的裂解中间体p43/p41。利用抗p27蛋白抗体进行免疫沉淀并未检测到总长度为57 kD的Caspase8。吉非替尼处理18 h和24 h后细胞质中p27蛋白和Caspase8的信号增强且结合在一起,见图4。

图3 吉非替尼处理不同时间共聚焦显微镜下p27蛋白表达(比例尺=20 μm)

a、b:抗p27蛋白抗体(a)、抗Caspase8抗体(b)对细胞质溶解产物孵育的IP结果;c:免疫细胞化学染色(比例尺= 20μm)图4 吉非替尼诱导细胞质中p27与Caspase8降解中间体p43/p41结合
3 讨 论
EGFR家族广泛表达的跨膜酪氨酸激酶参与多种肿瘤的发生发展。根据受体和配体的结合特点,同源或异源二聚体的受体类型及细胞质中酪氨酸残基的自身磷酸化产生的信号级联反应导致细胞增殖、血管生成、细胞迁移和凋亡抑制〔8〕。靶向EGFR已经成为一种切实可行的针对晚期NSCLC患者的治疗策略。吉非替尼作为口服小分子EGFR酪氨酸激酶抑制剂,目前已广泛应用于NSCLC的临床治疗。部分NSCLC患者存在EGFR酪氨酸激酶结构域突变,且这种突变在亚洲不吸烟的患腺癌的女性中尤为常见〔9〕。EGFR突变预示着其对酪氨酸激酶抑制剂具有极高的反应率〔10〕。尽管临床效果值得肯定,但吉非替尼诱导细胞凋亡和细胞周期抑制的机制迄今仍不明确。
p27蛋白在多种细胞的增殖、分化和死亡中发挥关键作用〔11〕。研究报道,p27蛋白对肿瘤细胞具有促凋亡作用,通过重组腺病毒对p27过表达引起了肿瘤相关细胞系的凋亡〔12〕。此外,已经发现在肺癌细胞系中p27蛋白过表达引起的细胞死亡可能受到pRb蛋白的调控〔13〕。而且,有研究发现,对比于高表达p27蛋白,胃癌中p27蛋白的低表达展现了一个较低水平的细胞凋亡和较差的不良预后〔14〕。p27蛋白的亚细胞定位对于其功能的发挥至关重要。前人已在多种人类肿瘤中检测到p27蛋白的细胞质定位〔15〕。本研究发现NSCLC细胞系中p27蛋白的细胞质定位可能是由于吉非替尼诱导的p27蛋白的动态核转出所引发。
本文同样发现吉非替尼诱导的细胞凋亡与Caspase8的激活相关。Caspase8的两条主要凋亡路径已被确定〔16〕。受体-配体作为外在凋亡路径需要配体结合到细胞表面受体,引起受体寡聚化,激活起始凋亡蛋白酶Caspase8。第二条为从细胞内部激活凋亡蛋白酶的线粒体途径,这个内在路径受到Bcl-2蛋白家族调控并通过Caspase9启动信号级联。本研究还发现数种Bcl-2家族蛋白,包括促凋亡蛋白Bax、Bad及抗凋亡蛋白Bcl-2、Bcl-xl在吉非替尼干预后表达并未发生显著改变。由于Caspase9在吉非替尼干预后同样未检测到明显激活现象,因此猜测吉非替尼在HCC827细胞中并未通过内部凋亡路径诱导细胞凋亡。有研究报道,在肿瘤细胞中Caspase8调控的细胞凋亡与p27蛋白有关〔17〕。在骨肉瘤中,蛋白酶体抑制剂MG132诱导的细胞凋亡过程中同时检测到Caspase8的激活增强及p27蛋白表达增加〔18〕。根据本研究结果猜测,吉非替尼干预NSCLC细胞后,细胞质中p27蛋白的表达升高和Caspase8的激活增强可能协同导致细胞凋亡。同样还发现吉非替尼的干预导致细胞质中p27蛋白与Caspase8的裂解中间体p43/p41结合。长度为57 kD的Caspase8已知是被含有前肽结构域和p18亚基的中间体p43/p41催化激活。随后,中间体p43/p41从功能前区释放粘连有p18的活性部位。此外,通过抑制p27蛋白的细胞质易位发现吉非替尼诱导的细胞死亡率下降,推断p27蛋白的核转位与Caspase8的结合在吉非替尼诱导的细胞凋亡进程中极为关键。
本文结果表明,大部分HCC827细胞在吉非替尼处理24 h后凋亡,但与免疫荧光检测结果不相符。究其原因发现,在免疫荧光法中,细胞必须被固定、透明化处理并在抗体孵育前对细胞进行冲洗。通过这一系列步骤,死亡的悬浮细胞被洗掉,而只有贴壁的活细胞被荧光检测。然而,无论是PARP或Caspase的裂解,还是p27蛋白的表达升高及其细胞质易位,在吉非替尼处理18 h后才被Western印迹清晰地检测到。推测这种结果差异很可能是Western操作中抗体对这些蛋白的敏感性差异所致。
1Khelwatty SA,Essapen S,Seddon AM,etal.Acquired resistance to anti-EGFR mAb ICR62 in cancer cells is accompanied by an increased EGFR expression,HER-2/HER-3 signalling and sensitivity to pan HER blockers〔J〕.Br J Cancer,2015;113(7):1010-9.
2Das BR,Bhaumik S,Ahmad F,etal.Molecular spectrum of somatic EGFR and KRAS gene mutations in non small cell lung carcinoma:determination of frequency,distribution pattern and identification of novel variations in Indian patients〔J〕.Pathol Oncol Res,2015;21(3):675-87.
3刘 莉,陆 远,王 媛,等.EGFR、HER2、CXCR4在非小细胞肺癌中的表达及临床意义〔J〕.现代生物医学进展,2014;14(6):1069-73.
4Kuwako T,Imai H,Masuda T,etal.First-line gefitinib treatment in elderly patients (aged ≥75 years) with non-small cell lung cancer harboring EGFR mutations〔J〕.Cancer Chemother Pharmacol,2015;76(4):761-9.
5Kuiper JL,Hendriks LE,van der Wekken AJ,etal.Treatment and survival of patients with EGFR-mutated non-small cell lung cancer and leptomeningeal metastasis:a retrospective cohort analysis〔J〕.Lung Cancer,2015;89(3):255-61.
6韩新华,田 薇,施公胜,等.胶质瘤组织中EGFR、p27kip1的表达变化及意义〔J〕.山东医药,2010;50(30):6-8.
7王林娜,侯素平,刘洪博,等.结直肠癌中RegⅣ和EGFR的表达及临床意义〔J〕.临床与实验病理学杂志,2015;31(12):1352-5.
8Bertotti A,Papp E,Jones S,etal.The genomic landscape of response to EGFR blockade in colorectal cancer〔J〕.Nature,2015;526(7572):263-7.
9Cadranel J,Ruppert AM,Beau-Faller M,etal.Therapeutic strategy for advanced EGFR mutant non-small-cell lung carcinoma〔J〕.Crit Rev Oncol Hematol,2013;88(3):477-93.
10Tang Y,Fang W,Zhang Y,etal.The association between PD-L1 and EGFR status and the prognostic value of PD-L1 in advanced non-small cell lung cancer patients treated with EGFR-TKIs〔J〕.Oncotarget,2015;6(16):14209-19.
11Lee JY,Ku BM,Lim SH,etal.The BIM deletion polymorphism and its clinical implication in patients with EGFR-mutant non-small-cell lung cancer treated with EGFR tyrosine kinase inhibitors〔J〕.J Thorac Oncol,2015;10(6):903-9.
12Jin X,Jin X,Sohn YW,etal.Blockade of EGFR signaling promotes glioma stem-like cell invasiveness by abolishing ID3-mediated inhibition of p27 (KIP1) and MMP3 expression〔J〕.Cancer Lett,2013;328(2):235-42.
13Fang Y,Wang Y,Wang Y,etal.A new tumour suppression mechanism by p27Kip1:EGFR down-regulation mediated by JNK/c-Jun pathway inhibition〔J〕.Biochem J,2014;463(3):383-92.
14Jameson MJ,Taniguchi LE,van Koevering KK,etal.Activation of the insulin-like growth factor-1 receptor alters p27 regulation by the epidermal growth factor receptor in oral squamous carcinoma cells〔J〕.J Oral Pathol Med,2013;42(4):332-8.
15Osaki LH,Gama P.MAPK signaling pathway regulates p27 phosphorylation at threonin 187 as part of the mechanism triggered by early-weaning to induce cell proliferation in rat gastric mucosa〔J〕.PLoS One,2013;8(6):e66651.
16Antonopoulos C,Russo HM,El Sanadi C,etal.Caspase-8 as an effector and regulator of NLRP3 inflammasome signaling〔J〕.J Biol Chem,2015;290(33):20167-84.
17Moreno-Galindo C,Hermsen M,García-Pedrero JM,etal.P27 and BCL2 expression predicts response to chemotherapy in head and neck squamous cell carcinomas〔J〕.Oral Oncol,2014;50(2):128-34.
18苗丽君,王 静,李珊珊,等.非小细胞肺癌组织中P27表达及定位与磷酸化AKT的关系〔J〕.癌症,2006;25(10):1216-20.
R73
A
1005-9202(2017)23-5771-04;
10.3969/j.issn.1005-9202.2017.23.011
1 武汉市第一医院病理科
冯 燕(1977-),女,主治医师,主要从事病理学研究。
杨 畅(1978-),男,主治医师,主要从事呼吸系统疾病研究。
〔2017-05-26修回〕
(编辑 袁左鸣/滕欣航)
