藏柏种群年龄结构数量动态分析
2017-12-19郑维列郭其强汪书丽张艳福
池 翔 ,郑维列 ,郭其强 ,汪书丽 ,张艳福
(1.西藏农牧学院,西藏 林芝 860000;2.西藏高原生态研究所,西藏 林芝 860000)
藏柏种群年龄结构数量动态分析
池 翔1,2,郑维列1,2,郭其强1,2,汪书丽1,2,张艳福1,2
(1.西藏农牧学院,西藏 林芝 860000;2.西藏高原生态研究所,西藏 林芝 860000)
为保护天然藏柏资源,通过设置典型样地,研究了西藏波密县易贡乡藏柏种群的年龄结构、静态生命表及存活曲线,运用时间序列模型预测了种群数量动态。结果表明:藏柏种群年龄结构呈“金字塔”型,幼龄个体储备丰富,存在断代现象,种群不稳定;存活曲线趋于Deevey Ⅲ型,幼龄期死亡率一直处于较高水平,存在两个死亡率高峰,分别出现在3、9龄级;未来经历3、5和7个龄级时间后,种群总体呈现出幼龄期减少,中龄期稳定,老龄期衰退的趋势。年龄结构和存活曲线都显示出藏柏种群有一定自我更新和维持的能力,但由于人为干扰、种间竞争和自疏作用等因素阻碍着种群的更新,使得种群趋于衰退,未来可通过砍灌、疏伐及清理林下活地被物等抚育措施提高藏柏幼龄个体存活率和中龄个体的存活量,减少人为破坏,以维持天然藏柏种群的长期稳定存在。
藏柏;年龄结构;静态生命表;数量动态;时间序列预测
种群年龄结构是植物生物学特性与环境因素长期相互作用的结果[1]。分析种群年龄结构不仅能反映物种的生存现状,还能反映植物与环境间的适合度[2]、种群受干扰状态,还能预测种群未来发展趋势[3]。通过对植物种群年龄结构特征分析还有助于揭示种群在群落中的地位和作用,这对种群的人工促进更新和复壮,以及生态系统恢复都具有重要意义[4]。
藏柏Cupressus torulosa为柏科,柏木属常绿乔木,树干通直,为优良用材、绿化观赏树种和造林树种,喜湿润、深厚、高钙质的土壤,也能耐贫瘠的山地[5-7],其原产为西藏东南部的墨脱、林芝、易贡等地区及印度北部[8],具有喜光、耐高寒等特性,对于高山生态系统的稳定与恢复具有重要意义。由于藏柏分布区偏僻且种群数量较少,针对藏柏的研究表明:育种过程中种子易萌发[9],水肥管理简单[10-11];造林方面藏柏被引种到贵州黔西县、凤岗县[5-8]和四川巴中县[6]后均生长良好,但需要做好赤枯病和银荆炭疽病防治[12];藏柏喜潮湿生境[13-15],且树干耐火性强[7];利用上,藏柏提取物可有效缓解小蠹侵害云南松Pinus yunnanensis枝梢[16]。由于藏柏分布区偏僻且种群数量较少,对藏柏原生境区的种群生存概况仍未见报道。以西藏波密县易贡乡藏柏种群为研究对象,开展藏柏种群年龄结构及数量动态的研究,对有效保护和合理管理天然藏柏资源,了解藏柏种群的生态特征及其对环境的适应具有重要意义[17]。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于西藏波密县易贡乡境内(95°1′20″~ 95°1′83″E,30°7′4″~ 30°8′19″N),海拔2 130~2 596 m,属山地暖温带湿润季风气侯,具有日温差较大,年温差较小、干雨季分明、气候垂直和水平差异显著等特点[18],土壤为砂土,呈微酸性,年均气温12 ℃,≥10 ℃年积温3 115 ℃,极端低温-11 ℃,极端高温为30.2 ℃,年均降水量972 mm,年均相对湿度71 % ,全年日照时长1 789 h,森林群落的建群种为藏柏C. torulosa。
1.2 样地设置与调查
2016年7月经野外踏查后,在藏柏种群分布区的西藏波密县易贡乡设置面积为30 m×30 m的藏柏典型样地4块(编号依次为A、B、C、D)。记录各样地的海拔、经纬度、地形、坡位、坡度、坡向、土壤状况、水分状况和人为干扰强度[19]。
乔木调查:对样地内的乔木进行每木调查和定位,记录乔木种名、高度、胸径(Dbh)、冠幅、活枝高和死枝高等基本数据。灌木调查:每块样地内沿对角线设置3块5 m×5 m的灌木样方,记录灌木种名、均高、盖度和多度等基本数据。草本调查:在各灌木样方内设置1块1 m×1 m的草本样方,记录草本种名、均高、盖度和多度等基本数据。
1.3 年龄结构的划分
乔木年龄可通过生长锥钻取树芯,数年轮数目而确定,由于藏柏结构细密,材质坚硬,不易钻木取芯[7],因此采用径级代龄级的方法来确定藏柏种群个体的年龄[20]。虽然种群龄级和径级有所不同,但在同一环境下,同一树种的龄级和径级对环境的反应规律具有一致性[21],因此,把树木径级从小到大的顺序视为时间顺序,第1径级对应第1龄级,第2径级对应第2龄级,依次类推[22]。本研究把胸径≤5 cm或树高≤1.5 m的个体划为第1龄级,对于胸径5 cm以上的个体以10 cm为径阶划分出共计15个龄级,其中第15龄级为φ>135 cm的藏柏个体,统计各龄级株数,以龄级为纵轴,株数/hm2为横轴,绘制藏柏种群年龄结构图。

表1 西藏波密县易贡乡调查样地的环境特征Table 1 Environmental characteristics of sample plots in Yigong township, Bomi county, Tibet
1.4 静态生命表的编制及生存分析
以各龄级数据为基础, 编制藏柏种群静态生命表和存活曲线[23],计算公式[24-27]如下:

式中:x为龄级数;ax为在x龄级内现有个体数;ao为ax的初始值;lx为在x龄级开始时标准化存活个体数(一般转化值为1 000);dx为从x到x+1龄级间隔期内标准化死亡数;qx为从x到x+1龄级间隔期间死亡率;Lx为从x到x+1龄级间隔期间存活的个体数;Tx为从x龄级到超过x龄级的个体总数;ex为进入x龄级个体的生命期望寿命;Kx为消失率。
1.5 时间序列预测模型
采用时间序列分析中的一次平均移动法对藏柏种群动态进行预测分析[4],其模型为:
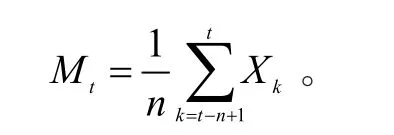
式中:n为需要预测的未来时间年限;t为龄级;Mt为未来n年时t龄级的种群存活数,是近期n个观测值在t时刻的平均值,称为第n周期的移动平均;Xk为当前k龄级种群存活数。应用该模型预测藏柏种群经历3、5和7个龄级时间后的发展趋势。
2 结果与分析
2.1 林分概况
研究区植物群落的垂直结构可明显分为乔木层、灌木层和草本层,林分郁闭度为0.9,乔木丰富度为13,灌木丰富度为8,草本丰富度为47。乔木最大高度为26.2 m(藏柏),最大胸径为167.4 cm(藏柏)。藏柏个数占乔木总个数的55.7%,常见物种即频度不小于75 %的物种,林分概况详见表2。

表2 天然藏柏林林分概况Table 2 Forest overview of natural C. torulosa forest
2.2 种群结构特征
从图1可以看出,藏柏种群的径级分布不连续,结构呈现出顶部窄基部宽的“金字塔”型,在第6龄级、第7龄级和第11龄级出现断代,藏柏种群个体主要集中分布于1~3龄级,占藏柏个体总数的74.58 %。这种结构表明,藏柏种群幼龄个体储备丰富,但幼龄个体向中龄个体过渡的过程中全部死亡,种群存在断代现象,导致更新过程被阻断,而且相对于老龄个体,中龄个体所占比重较小,种群趋于衰退。

图1 藏柏种群的龄级结构Fig.1 Age class structure of C. torulosa populations
2.3 静态生命表分析
从表3可以看出,藏柏种群幼龄期死亡率一直处于较高水平,其中第1龄级死亡率为负值,说明幼苗库不足,种群难以持续发展[19]。种群死亡率存在两个高峰:第一个高峰出现在第3龄级,死亡率高达72.7%;另一个高峰出现在第9龄级,死亡率为66.7%。自第10龄级开始,藏柏死亡率较低且保持平稳。种群生命期望寿命波动性较大,存在4个高峰,分别出现在第1、第5、第8和第10龄级处,其中第1和第5龄级生命期望寿命最大,分别为11.3 a和11.5 a,第1龄级个体生命期望寿命较高,说明幼苗耐荫性较强,第5龄级个体生命期望寿命较高,说明藏柏幼龄个体以高死亡率为代价通过严酷环境筛的过滤和筛选[4,28],存活的幼龄个体具有较强的生存能力,8~10龄级生命期望寿命出现波动,自第10龄级开始,生命期望寿命随年龄增加而减少,说明这些龄级的植株平均生存能力较强,可以活到理论生命值。

表3 藏柏种群静态生命表Table 3 Static life table of C. torulosa populations
2.4 存活曲线分析
从图2可以看出,藏柏种群的存活曲线趋于Deevey III型,早期死亡率较高,一旦存活到某一年龄,死亡率就较低且保持稳定[27]。第1龄级存活量远小于第2龄级存活量,2~4龄级各龄级死亡数较高,除去第9龄级出现的峰值,4~15龄级曲线相对平稳。

图2 藏柏种群存活曲线Fig.2 Survival curve of C. torulosa populations
2.5 发展趋势预测
从图3可以看出,藏柏种群各龄级株数峰值在预测序列中依次向后推移,总体呈现出幼龄期减少,中龄期稳定,老龄期衰退的趋势[17],其中4~8龄级各龄级株数在预测序列中呈上升趋势,第9、第10龄级株数在预测序列中呈下降趋势,11~14龄级各龄级株数在预测序列中保持稳定,第15龄级株数在预测序列中呈下降趋势。由此推断,藏柏种群虽具有较强的繁殖能力,但幼龄期的高死亡率导致种群在未来发展中幼龄个体不断减少,中龄个体数量不因幼龄个体更新受阻而得不到补充,老龄个体随年龄增长不断衰老、死亡,种群未来必将趋于衰退[17]。
3 结论与讨论
分析天然藏柏种群结构、静态生命表、存活曲线和数量动态得出一致结果,藏柏种群幼龄个体储备量丰富,有一定自我更新和维持的能力,但幼龄阶段极高的死亡率导致种群更新过程被阻断,中龄个体数量不仅因为人为干扰及自疏作用等因素影响难以增长,还因幼龄个体更新受阻而得不到补充,未来随着中、老龄个体的不断衰老、死亡,种群趋于衰退。
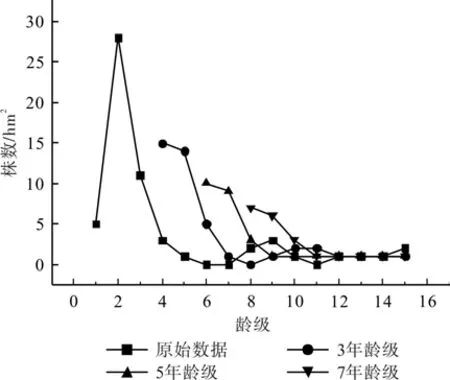
图3 藏柏种群数量动态时间序列预测Fig.3 Time sequence prediction of number dynamics of C. torulosa populations
从藏柏种群年龄结构来看,藏柏种群结构呈金字塔型,年龄分布不连续,表明种群稳定性不高,这与滇中地区翠柏Calocedrus macrolepis种群[29]、朗县巨柏C. gigantea种群[30]、雷公山秃杉Taiwania fl ousiana种群[31]的研究结果相似。种群结构显示藏柏种群第1龄级个体数量远少于第2龄级个体数量,这与湖南珙桐Davidia involucrata种群的研究结果相似[32],该研究认为珙桐第1龄级个体数量稀少是由于动物取食、种子萌发困难等原因造成的,而第2龄级个体数量较多是因为珙桐的萌孽能力较强。藏柏样地调查中发现人为干扰强度较大的A、B样地均有牛、猪等家畜活动,而且静态生命表分析表明第1龄级个体生命期望寿命较高,但实际存活量却较少,由此推断藏柏种群幼龄个体生长过程受到了较强的外界干扰。藏柏第1龄级数量较少可能与动物取食有关,第2龄级个体数量较多可能与藏柏生长特性有关,温跃明等[6]研究指出,藏柏苗木高和径生长有两次高峰期,因此以径级代龄级进行种群结构分析存在一定偏差。
种群生命表分析结果表明,藏柏幼龄期死亡率一直处于较高水平,这与雷公山秃杉T.fl ousiana种群的研究结果相似[31],与毛乌素沙地臭柏Sabina vulgaris种群[1]、湘鄂皖连香树Cercidiphyllum japonicum种群[4]、塔里木河上游胡杨Populus euphratica种群[22]、灵空山辽东栎Quercus wutaishansea种群[24]、色季拉山冷杉Abies fabri种群[26]的研究结果不同,即与第一个死亡率高峰出现在第一龄级,随后种群死亡率在第二个高峰期出现之前一直处于较低水平这个变化趋势不同。样地调查中发现,在灌木密集分布、灌木盖度远高于灌木平均盖度的C、D样地,藏柏幼龄个体只占种群幼龄个体总数的22.73%,由此推断幼龄阶段的高死亡率可能是由于激烈的种间竞争造成的,相对于灌木、草本植株,藏柏幼苗生长缓慢,竞争力弱小,随着时间的推移与其他物种争夺阳光、水分及营养物质能力的差距越来越大而造成藏柏幼龄期死亡率一直处于较高水平。在第9、10龄级出现了第二个死亡高峰,死亡高峰出现后伴随着第11龄级个体的缺失,第二个死亡高峰的出现可能与藏柏个体开始进入主林层,对空间、光线和养分空前激烈的竞争而导致了强烈的自疏过程有关[26,33-34],在调查中发现人为干扰强度较大的A、B样地中没有中龄个体的分布,由此推断第二个死亡高峰的出现可能也与人为干扰有关。
天然藏柏种群能够较好的完成林下自然更新,但由于人为砍伐、放牧、藏柏幼龄个体种间竞争力较差及自疏作用等因素导致种群难以延续。鉴于目前现状,可采取就地保护措施,减少人为干扰,通过砍灌、清理林下活地被物等抚育措施为藏柏幼龄个体创造有利的生境,在种群发育到一定时期,通过进行人工稀疏(例如疏伐)来提高中龄个体的存活量,使藏柏种群能够稳定的更新和生长[21,29,33-34]。天然藏柏分布区攀爬困难,因此本研究中样地的选取存在一定局限性,如果要更全面了解天然藏柏种群结构特征,还需引入其他手段对分布于山脊处、陡峭涯壁处的藏柏种群进行调查,在今后的研究中,我们将对西藏柏木属群落物种组成特征进行比较分析,以探讨该树种在小尺度范围内不连续分布的成因。
[1]李文良,张小平,郝朝运,等.湘鄂皖连香树种群的年龄结构和点格局分析[J].生态学报,2009,29(6):3221-3230.
[2]Fuchs M A,Krannitz P G,Harestad A S.Factors affecting emergence and first-year survival of seedlings of Garry oaks(Quercus garryana) in British Columbia,Canada[J]. Forest Ecology and Management, 2000, 137(1/3): 209-219.
[3]Armesto J J, Casassa I, Dollenz O. Age structure and dynamics of Patagonian beech forests in Torres del Paine National Park,Chile[J]. Vegetatio, 1992, 98(1): 13-22.
[4]韩 路,王家强,王海珍,等.塔里木河上游胡杨种群结构与动态[J].生态学报,2014,34(16):4640-4651.
[5]宋亚新.藏柏引种造林研究[J].毕节学院学报,2011,29(4):116-119.
[6]温跃明.巴中中山引种藏柏试验初报[J].四川林业科技,1995,16(1): 41-43.
[7]李世友,胡小龙,马爱丽,等.中等径级藏柏树干的耐火性研究[J].福建林学院学报,2009,29(1):65-68.
[8]张承芬,汪萍珍.藏柏、墨西哥柏引种造林试验[J].贵州林业科技,2001,29(2):14-17.
[9]翟书华,樊传章,张光飞.紫茎泽兰的水浸提液对干香柏和藏柏种子萌发的影响[J].昆明学院学报,2011,33(3):90-91.
[10]胡丁猛,臧真荣,王翠香,等.不同水肥处理对藏柏容器苗生长的影响[J].安徽农业科学,2012,40(7):4095-4096.
[11]黄 杏,刘崇欣,周应书.浅谈藏柏两段育苗技术[J].农业装备技术,2013,39(4):39-39.
[12]甘家生,杨卫明,陆 斌.藏柏赤枯病和银荆炭疽病的初步观察[J].云南林业科技,1995(2):44-46.
[13]贾 晨,兰发正,熊大国,等.剑阁县藏柏人工林生长特性研究[J].四川林业科技,2016,37(3):15-19.
[14]赵 总,张兆国,唐兴联,等.川滇桤木与藏柏人工林分生长量对比分析[J].林业调查规划,2009,34(4):19-21.
[15]范林元,孙 维,赵航文,等.藏柏36个家系自由授粉子代测定与优选[J].西南林学院学报,2010,30(4):28-31.
[16]赵 宁,泽桑梓,杨 斌.藏柏提取物对纵坑切梢小蠹侵害云南松松梢的影响[J].中国森林病虫,2012,31(5):39-41.
[17]郭其强,卢 杰,罗大庆,等.西藏色季拉山阳坡林线方枝柏种群结构特征研究[J].西北林学院学报,2010,25(2):15-18.
[18]顾祖文.西藏易贡茶叶生产的农业气象条件分析[J].自然资源,1988,1(1):94-96.
[19]张文辉,许晓波,周建云,等.濒危植物秦岭冷杉种群数量动态[J].应用生态学报,2005,16(10):1799-1804.
[20]张桥英,张运春,罗 鹏,等.白马雪山阳坡林线方枝柏种群的生态特征[J].植物生态学报,2007,31(5):857-864.
[21]伊力塔. 山西灵空山辽东栎林群落特征研究[D]. 北京: 北京林业大学, 2008.
[22]张 婕,上官铁梁,段毅豪,等.灵空山辽东栎种群年龄结构与动态[J].应用生态学报,2014,25(11):3125-3130.
[23]张文辉,王延平,康永祥,等.濒危植物太白红杉种群年龄结构及其时间序列预测分析[J].生物多样性,2004,12(3):361-369.
[24]王 卓,黄荣凤,王林和,等.毛乌素沙地天然臭柏种群生命表分析[J].中国沙漠,2009,29(1):118-124.
[25]Yan G Q, Zhao G F, Hu Z H. Population structure and dynamics of Larix chinensis in Qinling range[J]. Chinese Journal of Applied Ecology, 2001, 12(6): 824-828.
[26]任青山,杨小林,崔国发,等.西藏色季拉山林线冷杉种群结构与动态[J].生态学报,2007,27(7):2669-2677.
[27]肖宜安,何 平,李晓红,等.濒危植物长柄双花木自然种群数量动态[J].植物生态学报,2004,28(2):252-257.
[28]林 芳,黄梓良,叶功富,等.茫荡山小叶青冈天然种群生命表分析[J].中南林业科技大学学报,2016,36(2):8-11.
[29]刘方炎,李 昆,廖声熙,等.濒危植物翠柏的个体生长动态及种群结构与种内竞争[J].林业科学,2010,46(10):23-28.
[30]郑维列,薛会英,罗大庆,等.巨柏种群的生态地理分布与群落学特征[J].林业科学,2007,43(12):8-15.
[31]陈志阳,杨 宁,姚先铭,等.贵州雷公山秃杉种群生活史特征与空间分布格局[J].生态学报,2012,32(7):2158-2165.
[32]刘海洋,金晓玲,沈守云,等.湖南珍稀濒危植物--珙桐种群数量动态[J].生态学报,2012,32(24):7738-7746.
[33]胡尔查,王晓江,张文军,等.乌拉山自然保护区白桦种群的年龄结构和点格局分析[J].生态学报,2013,33(9):2867-2876.
[34]熊 能,金则新,顾婧婧,等.千岛湖次生林优势种种群结构与分布格局[J].生态学杂志,2010,29(5):847-854.
Age structure and quantity dynamics analysis ofCupressus torulosa
CHI Xiang1,2, ZHENG Weilie1,2, GUO Qiqiang1,2, WANG Shuli1,2, ZHANG Yanfu1,2
(1. Xizang Agricultural and Anim al Husbandry College, Linzhi 860000, Tibet, China; 2. Institute of Plateau Ecology of Tibet, Linzhi 860000, Tibet, China)
In order to effectively protect naturalCupressus torulosapopulation, theC. torulosapopulation in Yigong township, Bomi county, Tibet was regarded as the research object and its age structure, static life table and survival curve were analyzed with the typical sample plot method, its number dynamics was predicted by the time series model. The results showed: Its age structure was a“Pyramid” type, seedling reserve was rich, there existed age structure de fi ciency, which brought population instability; its survival curve was similar to the Deevey III type, the early mortality rate was high, there were two mortality peaks, which appeared in 3, 9 age class, and the mortality became low and stable after the age of tenth; after 3, 5 or 7 age class time in the future, the population showed the trend of reduction in the juvenile period, middle age stable stability and recession of aging period. The age structure and survival curve showed that there was a certain ability of self renewal and maintain ofC. torulosapopulation, but because of human disturbance,species competition and self thinning effects lead to the decent of the age class phenomenon, the population tends to decline, we can take some measures,such as cutting shrub, cleaning up the living matter of forest and low intensity thinning to increase the survival rate of the seedlings and the survival of the middle age class individuals, and reduce man-made destruction in order to maintain the long-term stability of naturalC. torulosapopulation.
Cupressus torulosa; age structure;static life table; quantity dynamic; time sequence prediction
10.14067/j.cnki.1673-923x.2017.12.018
http: //qks.csuft.edu.cn
S714.2
A
1673-923X(2017)12-0114-06
2016-09-23
国家自然科学基金项目(31460207);西藏林芝森林生态系统定位研究项目(2012-LYPT-DW-016)
池 翔,硕士研究生
郑维列,教授;E-mail:1131317504@qq.com
池 翔,郑维列,郭其强,等 . 藏柏种群年龄结构数量动态分析[J].中南林业科技大学学报,2017, 37(12): 114-119.
[本文编校:文凤鸣]
