普通油茶杂交子代幼林经济性状的遗传分析
2017-12-19王开良姚小华滕建华曹永庆
林 萍 ,王开良 ,姚小华 ,滕建华 ,曹永庆
(1. 中国林业科学研究院 亚热带林业研究所,浙江 杭州 311400;2. 浙江省林木育种技术研究重点实验室,浙江 杭州 311400;3.浙江省金华市婺城区东方红林场,浙江 金华 321025)
普通油茶杂交子代幼林经济性状的遗传分析
林 萍1,2,王开良1,2,姚小华1,2,滕建华3,曹永庆1,2
(1. 中国林业科学研究院 亚热带林业研究所,浙江 杭州 311400;2. 浙江省林木育种技术研究重点实验室,浙江 杭州 311400;3.浙江省金华市婺城区东方红林场,浙江 金华 321025)
为了建立油茶杂交育种最佳亲本选配模式,提高育种效率,以浙江省金华市东方红林场6个普通油茶杂交组合子代林为试材,分析了亲本在杂交子代单株产果量、果实大小、出籽率、干籽出仁率、种仁含油率、脂肪酸成分含量等17个经济性状的一般配合力(GCA)和特殊配合力(SCA),估算了各性状的遗传参数。结果表明,除了鲜果干籽率外,其余16个经济性状在6个杂交组合间均存在显著差异;亲本在各性状的一般配合力差异达极显著水平,亲本间特殊配合力在单株产果量、单果质量、种仁含油率、油酸含量等重要育种形状上亦差异显著或极显著;除了含油率外,其余主要经济性状的亲本GCA方差都显著大于亲本在SCA方差,均受基因的加性效应控制为主、非加性效应次之;遗传力较低,广义遗传力在10.10%~46.80%间,狭义遗传力在0%~41.30%之间,受到较强的环境效应影响。综合分析主要育种性状的亲本GCA 、SCA效应,确定优良亲本为53号和81号,优良组合为53×81和178×81,其杂交子代单株产果量分别比平均产果量高78%和35%,单果质量比平均值分别高14%和11%,风干籽出仁率比总平均值分别高8%和5%,种仁含油率提高了5%和9%。参试亲本间遗传距离在0.254 5~1.791 8间,只与油脂中油酸、亚油酸含量在0.1水平上具有相关性,与其他经济性状间无显著相关性。因此,油茶杂交育种中,除了油酸含量外,针对幼林大部分经济性状,未评估过GCA的亲本材料,先评估其GCA比评估亲本间的SCA更重要。
普通油茶;半双列杂交设计;一般配合力;特殊配合力
杂交育种作为最有效的育种技术之一,在林木育种上被普遍应用[1-9],亲本的科学选配是杂交育种成功的先决条件[10]。开展杂交亲本一般配合力和亲本间的特殊配合力测定和估算,能够更高效的配置杂交组合,提高杂交育种效率。相关工作已经在诸多用材树种及经济作物中开展,但经济林树种杂交育种中亲本的配合力估算和评价鲜见报道。
油茶Camellia oleifera作为我国南方最主要的木本食用油料树种,栽培历史悠久,分布区域广[11-14],栽培面积大。目前我国油茶产业所用苗木以选择育种育成的二代无性系良种为主,继续以传统选择育种方式培育高产优质新品种的潜力较小,杂交育种已成为新一代高产、高抗的油茶新品种培育的重要手段和途径。早在上世纪40年代,已有研究者开始了山茶属种间杂交研究,我国研究者亦在油茶杂交育种工作中开展了少量遗传分析的有益探索[15-17],但目前油茶杂交育种还主要局限于简单的具有互补性状的亲本杂交,亲本对重要经济性状的一般配合力(GCA)、特殊配合力(SCA)、遗传力等重要参数分析还未开展。本研究选择油茶二代无性系中具有不同优良性状的4个无性系,采用半双列杂交设计6个杂交组合。通过对杂交子代测定林的17个经济性状测定分析,估算油茶亲本的遗传参数,为油茶杂交育种亲本选配提供参考依据。
1 材料与方法
1.1 供试材料
2009年11月上旬在浙江金华婺城区东方红林场国家油茶种质资源收集库内选择油茶二代无性系具有不同优良性状的长林53号、81号、178号、185号等为亲本(见表1),按照半双列交配设计控制杂交,每组合控制授粉200朵花。2010年10月底果实成熟,6个杂交组合共收获杂交果实173个,杂交种子546粒。杂交种子于2011年3月播种育苗,2013年1月起2年生苗造林。造林按照完全随机区组设计,10株小区,重复3次。

表1 4个无性系亲本的特征Table 1 Traits of four C. oleifera clones
1.2 试验方法
1.2.1 经济性状调查与测定
2015年10月待5%以上果实开裂时,每单株实测果实产果量。每单株随机取20个果实,测定单果质量、果实纵径、横径、单果籽数、单果籽质量,取平均值作为单株数据。鲜果干籽率、干籽出仁率、种仁含油率等测定采用姚小华等[18-19]的方法进行。油脂脂肪酸成分测定采用周长富等[20]的方法进行。
1.2.2 杂交亲本间遗传距离
2015年春季,采集亲本无性系的春稍嫩叶,采用CTAB法提取基因组DNA。采用SRAP法对样本DNA进行PCR扩增,所用SRAP引物序列及扩增程序、数据处理分析参考林萍等[21]方法。
1.3 统计分析方法
采用DPS统计软件对幼林经济性状数据进行分析,对不同组合间性状差异采用单因素方差分析模型xij=μ+αi+βj+εij估算,其中xij为个体的表型值,μ为总体平均值,αi表示处理效应,βj为区组效应,εij代表随机误差。若杂交组合间差异显著,按固定模型(模型Ⅰ)估算一般配合力及亲本间的特殊配合力方差,亲本一般配合力及亲本间的特殊配合力采用公式:

分别估算[22]。 式(1)、式(2)中:p为亲本数,按随机模型(模型Ⅱ)估算遗传方差分量,广义遗传力用估算,狭义遗传力用分别代表子代群体加性遗传方差、亲本的一般配合力方差、子代群体非加性遗传方差、亲本间的特殊配合力方差、子代群体遗传方差、子代群体表型方差及环境方差[22]。亲本间的遗传距离与子代性状的相关性分析采用Pearson积矩相关系数估算[22-23]。
2 结果与分析
2.1 子代单株产果量、果实和油脂脂肪酸成分含量等经济性状的差异分析
对各杂交组合的主要经济性状进行单因素方差分析,结果(见表2)表明,除了鲜果干籽率外,单株产果量、单果质量、果实纵径、横径、单果鲜籽质量、种子出仁率、种仁含油率、各脂肪酸成分含量等所有测定性状在杂交组合间差异均达到0.01极显著水平,组合内区组间差异不显著。参试子代的单株产果量、平均单果质量、鲜果干籽率、种仁含油率等产量相关性状的平均值分别为0.67 kg/株、19.46 g、29.31%和40.73%,单株产果量最高单株是最低单株的400倍、种仁含油率最高单株是最低单株的2.87倍;茶油的主要成分油酸含量最高单株(88.07%)比最低单株(79.46%)高8.61%。说明通过无性系间的控制授粉杂交,可为选择育种创造变异丰富的种质资源。
进一步对差异显著性状进行多重比较,结果(见表3)表明,单株产果量最高的组合为53×81,比所有参试个体总体平均值高78%,其次是178×81,单株产果量是总体平均值的1.35倍;单果质量较大的组合依次是53×81、53×185、178×53,三者间差异不显著,比所有参试个体单果质量平均值分别高14%、11%和7%;果实纵径最大的组合是178×53,比总体平均值高6%;单果籽质量最大的组合依次为53×81、53×185和178×53,这三者间差异不显著,分别比总平均值高23%、15%和14%;气干籽出仁率最高的组合为53×81,达到58.13%,比总平均值高8%;种仁含油率最高的两个组合依次为178×81和53×81,分别为44.33%和42.59%,两者间差异不显著;油脂中棕榈酸含量较高的组合分别是178×185和178×53,二者的含量均超过了7%;硬脂酸含量显著高于其他组合的是185×81,达到了2.09%;油酸含量最高的组合为185×81,油酸含量达到了85.07%,且所有的杂交组合的油酸含量均高于目前生产上良种的79%~82%的油酸含量;亚油酸含量最高的组合为178×185和178×53,二者的亚油酸含量均超过了7%,比总平均值依次高20%和10%;亚麻酸和20碳烯酸含量最高的组合分别为53×185和178×185,因该2种成分含量较低,其含量绝对值与其他组合间差异不大。综上所述,杂交组合53×81在单株产果量、单果质量、单果籽质量、气干籽出仁率、种仁含油率及油脂中油酸含量等油茶育种的关键指标上均表现出显著的优势,其次178×81组合在单株产果量、干籽出仁率、种仁含油率等指标上与53×81无显著差异,因此亦具有优势。

表2 油茶杂交子代经济性状方差分析结果†Table 2 Variance analysis of the economic traits of the F1 descendant of C. oleifera
2.2 杂交子代果实及油脂相关经济性状的GCA、SCA方差分析
利用固定模型对16个差异显著的经济性状进行配合力方差分析,结果(见表4)表明,除了20碳烯酸含量外,其余15个性状的亲本GCA在组合间均达到差异极显著水平;亲本间的SCA在果实横径、果实纵横径比、种仁含油率、油酸含量、亚油酸含量和20碳烯酸含量等6个性状上具有极显著差异,在单株产果量、单果质量、单果籽数、棕榈酸含量、硬脂酸含量等5个性状上在0.05水平上差异显著,在剩余5个性状上差异不显著。因此,除了20碳烯酸外,以其余15个性状为育种目标的杂交育种工作中,杂交亲本的GCA评估是非常有必要的;在估算了亲本的GCA后,关注产果量、种仁含油率、油酸含量、亚油酸含量等关键性状时,亲本间的SCA也需要重点评估。

表3 油茶杂交组合子代各经济性状的多重比较结果†Table 3 Comparing the differences of economic traits among cross combinations of C. oleifera
进一步对差异显著的亲本GCA进行多重比较(见表5)表明,4个亲本中,81号在单株产果量的GCA为0.36,显著高于其它亲本;53号亲本在单果质量的GCA为3.07,果实籽质量的GCA为2.09,均与其他3个亲本存在极显著差异,这与53号亲本果实大、出籽率高的特点一致; 81号和53号在种仁含油率性状上的GCA分别为3.28和0.97,具有较高的遗传力;178号在油酸、亚油酸含量性状的GCA分别为0.82和0.89,均显著高于其他亲本,这与178号亲本高油酸的特点一致;185号在风干籽出仁率上的GCA为-3.28,极显著低于其余亲本,在其余性状上的GCA也普遍较低,为表现较差的杂交亲本。
亲本间SCA差异显著性状的相应SCA值见表6。针对幼林单株产果量性状,178×185和53×81的SCA最高,均为0.13,但由于178号和185号亲本在单株产果量上的GCA均较低,因此178×185组合的单株产果量未表现出明显优势,而81号在单株产果量的GCA也显著高于其余亲本,因此53×81的单株产果量在所有测试组合中是最高的。在单果质量和种仁含油率性状上,均以178×81和53×185组合的SCA最高,分别为1.12和1.49,因此,虽然178号和185号在单果质量和种仁含油率性状上的GCA较低,这两个组合的单果质量仍然是较高的;另一方面,53×81组合的SCA在所有组合中最低,分别为-0.84和-2.40,但由于53号和81号在此两个性状上的GCA均较高,因此该组合的单果质量和种仁含油率仍然较高。在油酸含量性状上,亦以178×81和53×185组合的SCA最高,SCA值均为0.48。因本研究所用杂交群体仍处于幼林阶段,此处分析所得各亲本在单株产果量性状上的GCA和SCA只是针对幼林的初步结论,这些高配合力亲本在油茶盛产期的产果量性状上是否仍能表现出较高的配合力还有待于持续研究。

表4 油茶半双列杂交(无自交)配合力方差分析-固定模型(F值)†Table 4 Variance analysis of genetic effects of combining ability in half-diallel cross of C. oleifera-model (F value)

表5 亲本GCA多重比较Table 5 Comparison of the differences in general combining ability of parents of C. oleifera

表6 差异显著的各亲本组合间SCA效应值Table 6 Comparison of the differences in general combining ability of parents of C. oleifera
2.3 果实及油脂等经济性状的遗传力评估
采用随机模型估算各性状的遗传参数(见表7)表明,本研究中所有性状的遗传力都相对较低,广义遗传力在46.80%~10.10%间,广义遗传力最大的性状为果实纵横径比值,最小的为棕榈酸含量,因此,油茶杂交子代幼林的经济性状多受到较强的环境效应影响。狭义遗传力在0~41.30%之间,其中油酸含量和20碳烯酸含量的加性方差均为0,狭义遗传力为0,广义遗传力主要由非加性方差构成;单株产果量、单果质量、果实纵、横径、单果籽数、单果籽质量、干籽出仁率、硬脂酸含量等性状加性方差显著高于非加性方差,狭义遗传力相对较大;含油率加性方差和非加性方差分别为6.23和6.99,二者相当,狭义遗传力接近广义遗传力的1/2。根据全同胞的遗传方差理论,当近交系数等于0时,基因的加性效应是形成一般配合力的主因,而特殊配合力一般是由于杂交双亲的显性、超显性和上位偏差等非加性效应形成[24-25]。因此,在油茶杂交育种中,当以油脂中油酸含量为育种目标时,以考量亲本间的SCA为主,亲本的GCA对子代的油酸含量影响不大;在以子代单株产果量、单果质量、果实纵、横径、单果籽数、单果籽质量、干籽出仁率、硬脂酸含量等性状为育种目标时,考量亲本的GCA比亲本间的SCA更重要;以种仁含油率为育种目标时,亲本的GCA和亲本间的SCA均需要着重考量。

表7 油茶各经济性状的主要遗传参数估算值Table 7 The genetic parameters for economic traits of C. oleifera
2.4 亲本间遗传距离与子代性状的相关性分析
采用8对SRAP引物对油茶杂交亲本进行PCR扩增,共检测到234个多态性位点。利用软件Popgene1.32计算亲本间的遗传距离变化范围为0.254 5~1.791 8,平均遗传距离为0.797 8。采用DPS软件分析亲本遗传距离与子代果实及油脂等经济性状的相关性发现,亲本遗传距离只与油脂中亚油酸、油酸含量在0.1显著水平上具有相关性,与其它经济性状间无显著相关性(表8)。
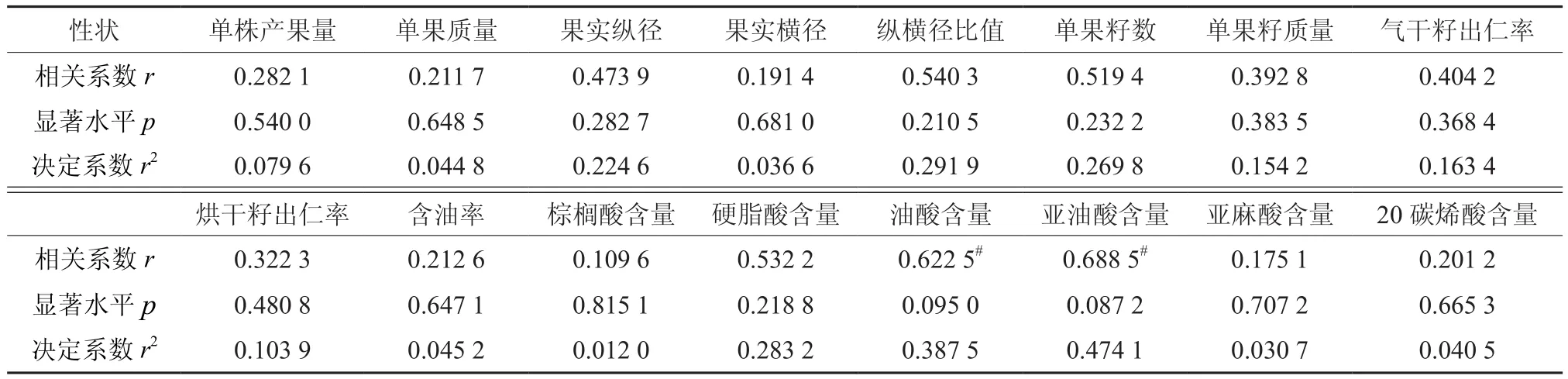
表8 亲本间遗传距离与子代性状的相关性†Table 8 Correlation analysis of parents genetic distance and descendant traits
亲本间遗传距离与子代亚油酸含量成负相关,与油酸含量成正相关,相关系数分别为-0.688 5和0.622 5;线性回归方程分别为y=7.125 9-1.008 368x和y=83.504 34+0.810 8x(图1)。
3 结论与讨论

图1 亲本遗传距离与子代亚油酸含量、油酸含量散点Fig.1 Scatter diagram on the genetic distance of parents and descendant oil composition
通常,利用GCA高的亲本杂交可望获得好的遗传增益,而SCA高则有可能在杂交后代中获得更高的杂交增益[26]。根据普通油茶半双列杂交子代幼林的各经济性状的配合力分析结果,综合考虑产果量、果实大小、干籽出仁率、种仁含油率、脂肪酸成分等关键经济指标,筛选出优良杂交亲本为53号和81号,178号在油酸含量的GCA显著高于其他亲本,但由于油酸含量的狭义遗传力为0,所以178号不能将高油酸含量的优良性状遗传给子代;综合考量各亲本在主要性状上的GCA、亲本间的SCA及遗传力等多个指标,确定53×81和178×81为优良杂交组合。本研究中,多数的经济性状均以81号和53号亲本的GCA最高,185号亲本基本在所有的性状上的GCA均是最低的,这与白桦Betula platyphylla[26]、杉木Cunninghamia lanceolata[5]、 桉 树Eucalyrtus globulus[27]、 油 松Pinus tabulaeformis[28]等树种中针对不同性状亲本间的一般配合力差异较大的现象不同。
测定和估算杂交亲本的GCA和SCA,可在杂交育种工作时根据育种目的有效的筛选最佳亲本和杂交组合,从而提高杂交育种效率。同时还可根据亲本配合力的估算结果,分析了解各性状的基因作用方式,制定最佳的遗传改良策略[29]。多个用材树种亲本配合力的研究发现,GCA和SCA的相对重要性受测试材料、性状、年龄和地点等因素的影响,通常GCA的选择比SCA更重要[30-31]。本研究对油茶半双列子代幼林的经济性状测定结果表明,17个性状中子代单株产果量、单果质量、果实纵径、果实横径、单果籽数、单果籽质量、干籽出仁率、种仁含油率、硬脂酸含量等多数性状,评估亲本的GCA值比亲本间的SCA值更重要;但油茶油脂中的油酸含量这一性状不同,其主要由非加性效应控制,评估亲本的SCA值比GCA值更重要。本研究中的油茶子代仍处于幼林阶段,亲本在各性状上的配合力是否随着测试样本树龄的增长而改变,还有待于进一步分析。本研究中各性状的全同胞家系狭义遗传力均较低,说明各性状受到较弱的遗传控制,受环境影响较大,因此通过筛选优良家系作为优良品种选育的策略是不可取的,在全同胞家系内选择优良单株从而培育成优良品种的效率相对更高,这与油茶亲本在生长性状上的配合力分析结果一致[32]。
在众多的用材树种[33-38]及经济农作物[39-41]中的研究证实,亲本间的分子遗传距离与杂交子代生长性状或经济性状间存在不同程度的相关性。但也有研究认为,亲本间分子遗传距离与子代杂种优势间的关系是变化的,这种变化依赖于研究中所用的遗传材料、亲本间遗传距离的范围以及杂种优势遗传基础的复杂性等[42]。本研究的分析结果表明,亲本间的遗传距离只与油酸含量、亚油酸含量间有显著相关,且相关系数亦较高,分别为0.622 5和-0.688 5,与其余性状无显著相关性。这与马尾松[38]、水稻[41]等只有亲本间遗传距离在一定范围内时才与其重要经济性状具有显著相关性,扩大遗传距离范围,这种相关性便不存在的研究结果不同。本研究中油茶亲本间的遗传距离分布范围较广,在0.254 5~1.791 8之间,油茶亲本的遗传距离是否在较小的范围内与其余经济性状间有相关性还有待于进一步研究。
[1]CARSON S D. Control-pollinated seed orchards of best general combiners-a new strategy for radiata pine improvement[J].Agronomy Society of New Zealand, 1986(5): 144-148.
[2]SLUDER E R. Results at age 15 years from a half-diallel cross among 10 loblolly pines selected for resistance to fusiform rust(Cronartiumquercuumf. sp. Fusiforme)[J]. Silvae Genetica, 1993,42(4-5): 223-230.
[3]叶培忠, 陈岳武,刘大林, 等.配合力分析在杉木数量遗传研究中的应用[J]. 南京林产工业学院学报, 1981, 5(3): 1-21.
[4]王赵民, 陈益泰. 杉木主要生长性状配合力分析及杂种优势的利用[J]. 林业科学研究, 1988, 1(6): 614-623.
[5]李 力, 施季森, 陈孝丑, 等. 杉木两水平双列杂交亲本配合力分析[J]. 南京林业大学学报, 2000, 24(5):8-13.
[6]张含国, 袁桂华,李希才, 等. 落叶松生长和材性杂种优势的研究[J]. 东北林业大学学报, 1998,16(3):25-28.
[7]Randall W K. Advance of breeding in Aigeirossection[J].Proceedings Symposium on Eastern Cottonwood and Related Species, 1976:140-150.
[8]Viart M. Importance of Populusdeltoides to poplar silviculture in France[J]. Proceedings Symposium on Eastern Cottonwood and Related Species, 1976:38-44.
[9]Sekawin M. Poplar breeding in northern Italy, IncludingP.deltoids[J]. Proceedings Symposium on Eastern Cottonwood and Related Species, 1976:170-175.
[10]张爱民. 植物育种亲本选配的理论和方法[M]. 北京: 农业出版社, 1994.
[11]庄瑞林. 中国油茶[M]. 2版. 北京: 中国林业出版社, 2008.
[12]曹永庆, 任华东, 姚小华. 油茶低产林截干更新技术研究[J].经济林研究, 2015, 33(1):111-114, 118.
[13]曹永庆, 姚小华, 任华东, 等. 油茶果实矿质元素含量和油脂积累的相关性[J].中南林业科技大学学报,2013,33(10):38-41.
[14]宋祥兰, 王兰英, 邝先松, 等. “赣州油”系列油茶容器育苗基质配方实验初探[J]. 中南林业科技大学学报, 2014,34(1):23-26.
[15]孙鸿有, 林其盛, 沈建忠. 油茶交配亲和力的初步研究[J]. 浙江林学院学报, 1984, 1(1):105-110.
[16]张乃燕, 陈国臣, 王东雪, 等. 岑软2号、3号等油茶无性系花粉配合力测定[J]. 广西林业科学, 2009, 38(1):1-4.
[17]陈 辉, 刘佳洁, 刘国敏, 等. 油茶F1代试验林优良杂交组合及亲本的选择[J]. 福建林学院学报, 2008,28(4):304-309.
[18]姚小华, 王亚萍, 王开良, 等. 地理经纬度对油茶籽中脂肪及脂肪酸组成的影响[J]. 中国油脂, 2011, 36(4):31-34.
[19]谢一青, 李志真, 姚小华, 等. 小果油茶果实性状与含油率及脂肪酸组成相关性分析[J]. 中国油脂, 2013, 38(5):80-83.
[20]周长富,姚小华,林 萍, 等. 油茶种子发育过程组分含量动态研究[J].中国油料作物学报, 2013,35(6):680-685.
[21]林 萍, 姚小华, 王开良, 等. 油茶长林系列优良无性系的SRAP分子鉴别及遗传分析[J], 农业生物技术学报, 2010,18(2): 272-279.
[22]唐启义, 冯明光. 实用统计分析及其DPS数据处理系统[M].北京: 科学出版社, 2002.
[23]Ghosh H, DAS A. Optimal diallel cross designs for estimation of heritability[J]. Journal of Statistical Planning and Inference,2003,116: 185-196.
[24]Mullin T J, PARK Y S. Estimating genetic gains from alternative breeding strategies for clonal forestry[J]. Canadian Journal of Forest Research, 1992, 22(1):14-23.
[25]盛志廉, 陈瑶生. 数量遗传学[M]. 北京: 科学出版社, 1999.
[26]李开隆, 姜 静, 姜 莹, 等. 白桦5×5完全双列杂交种苗性状的遗传效应分析[J]. 北京林业大学学报, 2006, 28(4):82-87.
[27]李淡清, 刘永平, 曾德闲, 等. 蓝桉6×6全双列交配生长性状的遗传效应分析[J]. 遗传学报, 2002, 29(9):835-840.
[28]郭俊荣, 杨培华, 谢 斌, 等. 油松双列杂交实验配合力分析[J]. 西北林学院学报, 1997,12(4):13-18.
[29]刘青华, 金国庆, 储德裕, 等. 基于马尾松测交系子代的生长、干形和木材密度的配合力分析[J]. 南京林业大学学报(自然科学版), 2011, 35(2):8-14.
[30]Sprague G F, TATUM L A. General vs. specific combining ability in single crosses of corn[J]. Journal American Society of Agronomy, 1942, 34(4):923-932.
[31]齐 明. 杉木育种中GCA与SCA的相对重要性[J]. 林业科学研究, 1996, 9(5):498-503.
[32]林 萍, 姚小华, 滕建华, 等. 油茶杂交子代幼林生长性状的遗传分析[J]. 经济林研究, 2016, 34(1):6-11.
[33]李 梅, 甘四明. 杉木杂交亲本分子遗传变异与子代生长相关的研究[J]. 林业科学研究, 2001,14(1):35 -40.
[34]Kopp R F, Smartl B, Maynardc A,et al.Predicting withinfamily variability in juvenile height growth of Salix based upon similarity among parental AFLP fi ngerprints[J]. Theoretical and Applied Genetics, 2002,105(10):106-112.
[35]陈考科, 黄少伟. 桉树的分子标记技术及遗传图谱构建进展[J]. 分子植物育种, 2005, 3(2):255-260.
[36]李善文, 张有慧, 张志毅, 等. 杨属部分种及杂种的AFLP分析[J]. 林业科学, 2007, 43(1):35-41.
[37]李周歧, 王章荣. 用RAPD标记进行鹅掌楸杂种识别和亲本选配[J]. 林业科学, 2002, 38(5):169-174.
[38]张 一, 储德裕, 金国庆, 等. 马尾松亲本遗传距离与子代生长性状相关性分析[J], 林业科学研究, 2010, 23(2):215-220.
[39]桑世飞, 王 会, 梅德圣, 等. 利用全基因组SNP芯片分析油菜遗传距离与杂种优势的关系[J]. 中国农业科学, 2015,48(12): 2469-2478.
[40]赵庆勇, 朱 镇, 张亚东, 等. SSR标记遗传距离与粳稻杂种优势的相关性分析[J]. 中国水稻科学, 2009, 23(2):141-147.
[41]罗小金, 贺浩华, 彭小松, 等. 利用SSR标记分析水稻亲本间遗传距离与杂种优势的关系[J]. 植物遗传资源学报, 2006,7(2):209-214.
[42]Zhang Q F, Zhou Z Q, Yang G P,et al.Molecular marker heterosisgosity and hybrid performance in indica and japonica rice[J]. Theoretical and Applied Genetics, 1996, 93:1218-1224.
Genetic analysis of the economic traits ofCamellia oleiferaF1 descendant inhalf-diallel cross design
LIN Ping1,2, WANG Kailiang1,2, YAO Xiaohua1,2, TENG Jianhua3, CAO Yongqing1,2
(1. Research Institute of Subtropical Forestry, Chinese Academy of Forestry, Hangzhou 311400, Zhejiang, China;2. Key Laboratory of Tree Breeding of Zhejiang Province, Hangzhou 311400, Zhejiang, China;3. Dongfanghong Forest Farm of Zhejiang Province, Jinhua 321025, Zhejiang, China)
In order to improve the breeding ef fi ciency ofCamellia oleifera, selecting optimum parents for the cross breeding is needed.The genetic parameters ofCamellia oleiferaparents were estimated by analyzing 17 economic traits of F1 descendant, such as the yield,fruit size, seeds content of fruit, kernel content of dry seed, oil content of kernel, fatty acid kinds and content in seed oil, and so on.The results showed that the genetic difference was extremely signi fi cant in all economic traits among families but dry seeds content of fresh fruit, the GCA effects of parents were signi fi cant different in 16 economic traits and SCA effects had signi fi cant difference in main breeding traits such as yield, fruit weight, oil content of kernel, oleic acid content of oil, and so on. Additive effects of genes played the major role and non-additive effects were next for the 16 traits except oil content of kernel. The heritability was low in all traits, the broadsense heritability was between 10.10%—46.80% and the narrow-sense heritability was 0%—41.31%.According to the statistical analysis results of GCA and SCA, the Changlin53# and 81# will be better parents in cross breeding, and Changlin53#×81# and 178#×81# are superior cross combinations, which were 78% and 35% higher than the average value on yield, 14% and 11% higher than the average value on fruit weight, 8% and 5% higher than the average value on kernel content of dry seed, 5% and 9% higher than the average value on oil content of kernel, respectively. The genetic distances of parents were 0.254 5—1.791 8, which were signi fi cant correlation with oleic acid and linoleic acid content of oil at 0.1 level. In a word, the estimate of the GCA of parents is more important than that of SCA between parents for the main economic traits except oleic acid content of oil inCamellia oleiferacross breeding.
Camellia oleiferaAbel.; half-diallel cross design; general combining ability (GCA); speci fi c combining ability (SCA)
10.14067/j.cnki.1673-923x.2017.12.006
http: //qks.csuft.edu.cn
S794.4
A
1673-923X(2017)12-0031-08
2016-09-13
国家自然科学基金青年基金项目(31400580);中央级公益性科研院所基本科研业务费专项资金项目(RISF2013008)
林 萍,助理研究员,博士
姚小华,研究员,博士,博士生导师;E-mail:yaoxh168@163.com
林 萍,王开良,姚小华,等. 普通油茶杂交子代幼林经济性状的遗传分析[J].中南林业科技大学学报,2017, 37(12): 31-38.
[本文编校:文凤鸣]
