长白猪系间杂交初生重性状配合力分析
2021-11-15李世豪杨又兵王丁丁王丹晖马彦博
李世豪 杨又兵 娄 然 王丁丁 王丹晖 马彦博
河南科技大学动物科技学院/洛阳市动物遗传育种重点实验室,河南洛阳 471023
仔猪初生重是影响猪场经济效益的重要因素,其大小对仔猪成活率、断奶重和生长速度等方面有非常重要的影响。林俊英[1]对632 头大白仔猪进行跟踪试验,发现仔猪初生重与仔猪成活率呈正相关关系;陈方琴[2]的研究结果表明,仔猪的初生重与21日龄体重成正比,21日龄体重随初生重的增加而增加;赵云翔等[3]对3 个不同品系杜洛克的初生重与生长速度进行分析发现,杜洛克公猪出生后的生长速度随其初生重的增加而加快,还有许多研究[4-6]与上述结果相似,较高的仔猪初生重有助于提高养殖户的经济效益。通过杂交育种可以有效地提高仔猪初生重[7]。配合力是杂交中体现亲本各性状配合能力的指标,是选择亲本的依据,通过配合力分析可以在较早世代初步鉴定出组合和亲本的优劣,大幅度缩小处理材料和范围的时间,提高育种效率。
长白猪育成已有百年时间,各国利用它与本地猪种杂交,已经形成了十多个品系[8],不同品系间的繁殖性状存在着一定的差异[9]。长白猪与其他猪种间的杂交试验比较多,但各个品系间的杂交利用相对较少。黄瑞华等[10]通过将英系长白猪与丹系长白猪杂交,对屠宰性能进行比较分析发现2 个品系杂交一代的一些胴体品质显著优于亲本,可见长白猪不同品系间杂交亦可获得杂种优势。本研究通过对丹系、法系和加系3 个品系长白猪采用双列杂交方法进行系间杂交,对初生重性状进行配合力分析和遗传力估算,评价亲本的应用价值,了解性状的遗传方式,以期为长白猪的改良及品系间杂交利用提供参考依据。
1 材料与方法
1.1 试验动物
本研究的试验猪只均来自江西省某育种场,育种场内饲养了丹系、法系、加系3个品系长白猪。试验猪只在0日龄进行初生选留,选择符合品种特征,生长发育良好无畸形猪只进行剪牙、断尾、补铁、打耳缺等操作;28日龄进行断奶选留,剔除生长发育不良及出生时未发现畸形的已打耳缺猪只;70日龄进行保育选留,继续剔除在保育舍生长发育不良的猪只;150日龄在猪只100 kg 左右进行结测;165日龄开始诱情,225日龄进行初配。不同阶段猪只分别根据其需要由该育种场提供相应的配合饲料。
1.2 杂交方法
2019年9-11月在该育种场按照Griffing 完全双列杂交方法第1 种设计杂交试验。如表1所示,杂交后代有6 个正反交组和3 个纯繁组,共9 个组合,每个组合配种母猪10头。

表1 完全双列杂交试验设计
1.3 性能测定方法
试验所测定的数据于2019年12月至2020年3月测定完成,每个组合选取10头仔猪进行初生重测定,重复3次,共测定270头仔猪的初生重。
1.4 统计分析
所得数据用Excel 2016 软件按照Griffing 介绍的配合力分析方法第1种进行配合力分析和遗传力估算[11-15]。
2 结果与分析
2.1 组间差异性检验
完全双列杂交的9个组合初生重方差分析表见表2,经F检验表明,在固定模型和随机模型2 种模型的假定上,初生重组合间差异均极显著,需要进一步进行配合力分析。
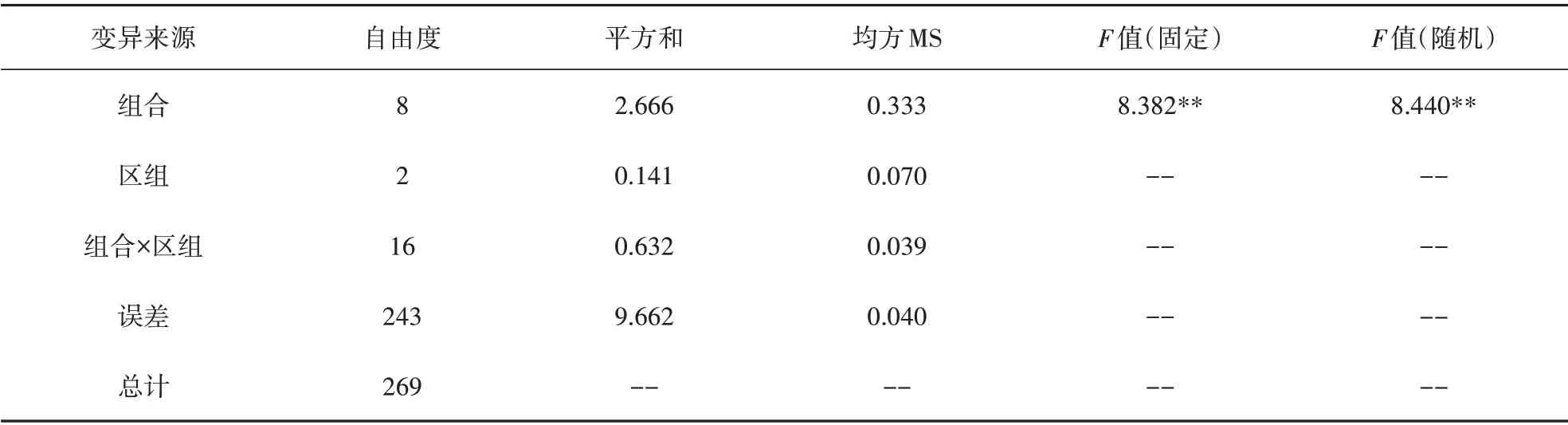
表2 初生重组间方差分析
2.2 配合力方差分析
为进行配合力方差分析,列初生重组合均数二向表(表3)。

表3 初生重组合均数二向表kg
对配合力进行方差分析,分别计算一般配合力、特殊配合力和反交效应的平方和。

随机模型中一般配合力的误差M*=(1-A)M′e+AMs,而,M*的自由度f=,其中k=
通过计算得到配合力方差分析结果列于表4。在固定模型和随机模型中,初生重一般配合力及特殊配合力差异均达到极显著水平,可以确定这些效应间存在真实差异,能够进行配合力效应值的估算和遗传力的估算。
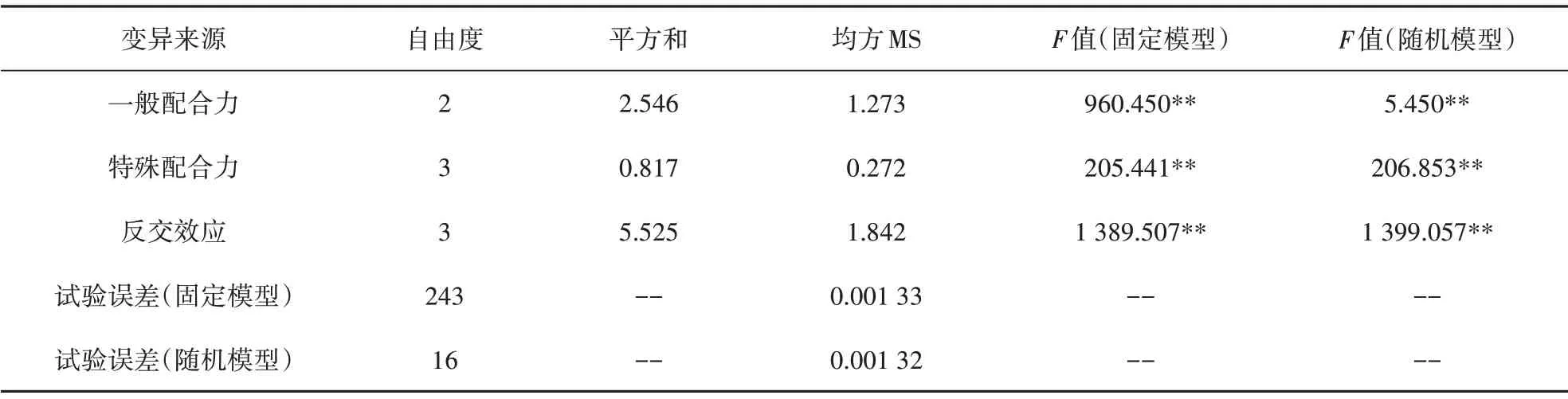
表4 初生重配合力方差分析
2.3 配合力效应值分析
配合力的好坏需要根据育种目标来确定,在育种中希望通过杂交提高初生重来提高经济效益,所以其配合力为正值时杂交效果更好。一般配合力(gca)用于表示杂交组合中某一亲本系的平均表现,是加性效应的体现,一般配合力越大,亲本对性状传递能力越强。
一般配合力效应值估算:
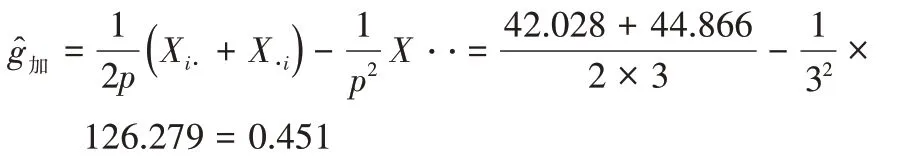
其余类推……
为了确定一般配合力效应是否真实存在,需要对其进行显著性检验。同时为了掌握配合力各效应值之间的显著性差异,还需要进一步比较各效应值之间的效应差数。
一般配合力效应值的标准差及效应差数标准差为:
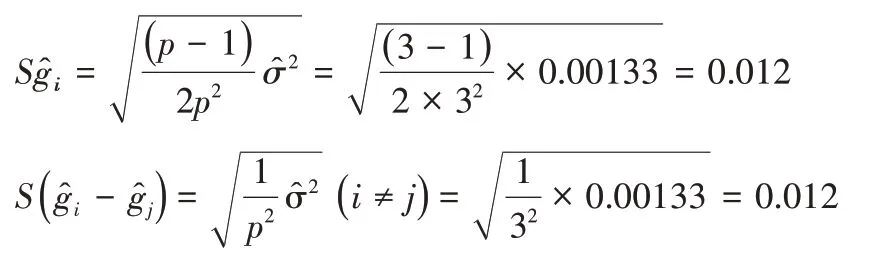
当误差项自由度为243 时,t0.05=1.965,t0.01=2.586,所以L.S.D临界值如下,一般配合力效应值及效应差数大于|(L.S.D)0.05|就有95%的把握肯定其真实存在。

对一般配合力效应值估计、检验后得到表5,各亲本初生重的一般配合力效应值范围为-0.469~0.451,加系和法系的一般配合力效应值为正值,丹系为负值,加系的一般配合力最好,极显著高于法系和加系。说明加系在提高初生重的育种中效果极显著,法系不显著,而丹系负向效果极显著,不利于初生重的提高。所以育种中利用加系作为亲本可以有效提高初生重。

表5 初生重一般配合力估计、检验及其差值检验
特殊配合力(sca)用于表示一定组合的表现比之由其所包含的亲本系的平均表现预期要更好或更差,是非加性效应的体现,特殊配合力越大,2 个亲本杂交时杂种优势越强。特殊配合力虽和杂种优势率很像,但它们实际上是不同育种学家在不同时期分别提出的2套统计学概念[16]。
特殊配合力效应值估算如下:

其余类推……
特殊配合力效应值的标准差及效应差数的标准差为:

当误差项自由度为243 时,t0.05=1.965,t0.01=2.586,所以L.S.D 临界值如下,特殊配合力效应值及效应差数大于|(L.S.D)0.05|就有95%的把握肯定其真实存在。
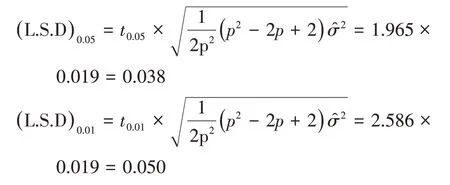
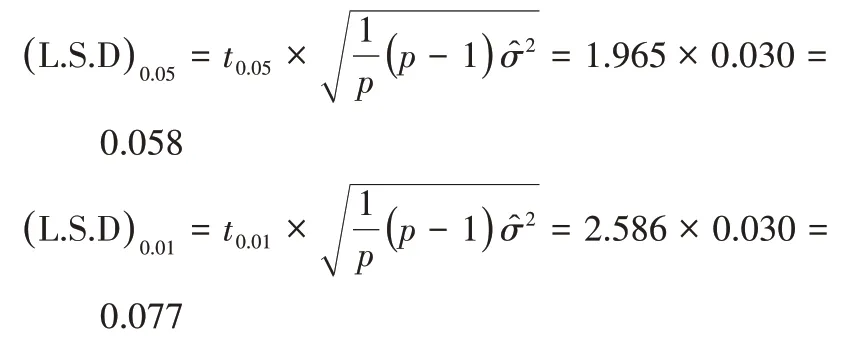
对特殊配合力效应值估计、检验后得到表6,各组合初生重的特殊配合力效应值范围为-0.292~0.309,法加组合和丹加组合特殊配合力效应值为正值,丹法组合为负值,法加组合特殊配合力最好,极显著高于丹法组合。说明法加组合和丹加组合在提高初生重的育种中效果极显著,而丹法组合负向效果极显著,不利于初生重的提高。组合的优劣不能仅靠特殊配合力确定,应该同时考虑一般配合力,所以还需要计算组合的配合力总效应才能确定。

表6 初生重特殊配合力估计、检验及其差值检验
反交效应用于表示相同组合父母本交换时后代的差异程度,反交效应越大,正反交组合间性状差异越大。配合力总效应结合了双亲的一般配合力和组合的特殊配合力,同时考虑了加性效应、非加性效应和上位效应,较仅靠特殊配合力判断组合优劣来说更加准确。
反交效应值估计:
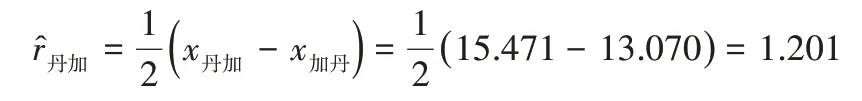
其余类推……
反交效应值的标准差为:
当误差项自由度为243 时,t0.05=1.965,t0.01=2.586,所以L.S.D 临界值如下,反交效应值大于|(L.S.D)0.05|就有95%的把握肯定其真实存在。
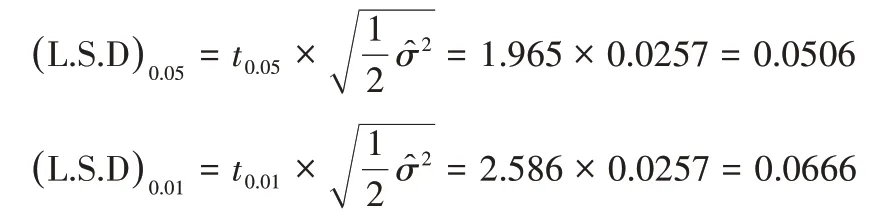
组合的配合力总效应计算如下:
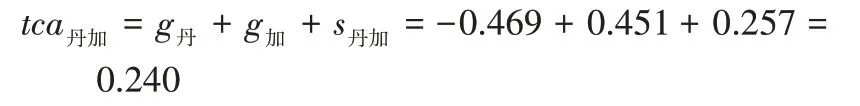
对反交效应值估计及配合力总效应计算后得到表7,各组合的初生重反交效应均极显著,丹加组合反交效应最大。法加组合的配合力总效应最大,所以育种中利用法加组合可以有效提高后代初生重,由表3可知加系做母本时杂交效果更好。

表7 初生重反交效应估计及配合力总效应
2.4 亲本特殊配合力方差估算
亲本特殊配合力方差用于表示各亲本在各组合中性状传递能力的整齐度,需要借助每个亲本的配合力效应值求出,凡是亲本特殊配合力方差小的亲本,表示该亲本能够较为整齐地将其性状传递给所有组合的子一代,反之则表示变异程度较大。计算初生重亲本特殊配合力方差,结果见表8,丹系特殊配合力方差相对较小,杂交后代性状整齐度最好,加系和法系次之,后代性状变异程度较大。

表8 初生重亲本特殊配合力方差
2.5 遗传力估算
广义遗传力是性状的遗传方差占表型方差的比值,体现了后代性状的变异主要由遗传因素影响还是环境因素影响。狭义遗传力是性状的加性方差占表型方差的比值,遗传方差中的加性方差可以固定遗传,育种中狭义遗传力比广义遗传力更可靠,更加接近于实际。计算初生重各遗传参数,结果见表9,初生重性状的广义遗传力很高,达到了92.95%,说明性状遗传时主要由遗传因素影响,受环境影响较小。狭义遗传力为61.85%,说明加性方差占到了表型方差的一半以上,遗传较为稳定。

表9 初生重遗传参数估算
3 讨 论
3.1 亲本应用价值评价
一般配合力用于表示杂交组合中某个亲本的平均表现[17],是加性效应的体现,一般配合力越大,亲本对性状传递能力越强。特殊配合力方差是亲本参与的所有杂交组合后代整齐度的体现,特殊配合力方差越大,后代整齐度越低,变异程度越大,可能出现偏离一般配合力效应所估计的较极端的后代。判断亲本的利用价值可分为4 种情况:①一般配合力大、特殊配合力方差大,这类亲本最好;②一般配合力效应高、特殊配合力方差小,这类亲本较好;③一般配合力效应低、特殊配合力方差较大,这类亲本利用价值较小;④一般配合力效应低、特殊配合力方差小,这类亲本无利用价值[18]。本研究中加系的一般配合力最大,同时加系的特殊配合力方差较大,所以加系是3 个品系中提高初生重性状时的优良亲本。
3.2 优势组合评价
特殊配合力用于表示一定组合的表现比之由其所包含的亲本系的平均表现预期要更好或更差,是显性效应和上位效应的体现,特殊配合力越大,2个亲本杂交时杂种优势越强。在对杂交组合进行评价时,首先要看亲本的一般配合力大小,其次需要关注组合的特殊配合力大小,因为杂交效果是受加性效应、显性效应和上位效应共同影响,所以应该用配合力总效应值来对杂交组合的优劣进行评价。本研究中丹加组合总配合力为0.240,丹法组合为-0.743,法加组合为0.779,法加组合的配合力总效应值最大,所以法加组合是3 个组合中提高初生重性状时的优势组合,加系做母本时杂交效果更好。
3.3 性状的遗传特点
广义遗传力在计算时不仅包含了基因的加性效应方差,还包括显性效应等的方差,而实际上显性效应并不能稳定遗传,计算时不应考虑。因此,狭义遗传力在育种中比广义遗传力更接近实际[19]。一般认为狭义遗传力高的性状早期选择有较大的可靠性,适合用于个体或群体表型选择法进行选种,采用纯繁来提高;狭义遗传力低的性状早期选择效果较差,适合用于家系选择或家系内选择,通过杂交利用其杂种优势[20]。本研究中初生重性状的广义遗传力达到了92.95%,狭义遗传力为61.85%,属于高遗传力,应该采用纯种繁育方法,通过个体或群体表型选择法在早期世代对性状进行选择。这与强巴央宗等[21]和王青来等[22]估计的0.28 和0.342 差别较大,可能原因是估算遗传力时采用的方法和环境方差不同导致[23]。
4 结 论
配合力分析表明,加系是3 个品系中提高初生重性状时的优良亲本,法加组合是优势组合,加系做母本时杂交效果更好。初生重性状遗传时主要由遗传因素影响,遗传较为稳定,应该采用纯种繁育方法,通过个体或群体表型选择法在早期世代对性状进行选择。
