盐度对不同规格可口革囊星虫耗氧率和排氨率的影响
2017-12-18曾梦清游出超刘旭佳刘海娟
曾梦清,蒋 艳,游出超,刘旭佳,刘海娟
( 广西海洋研究所,广西海洋生物技术重点实验室,广西 北海 536000 )
盐度对不同规格可口革囊星虫耗氧率和排氨率的影响
曾梦清,蒋 艳,游出超,刘旭佳,刘海娟
( 广西海洋研究所,广西海洋生物技术重点实验室,广西 北海 536000 )
水温18.5~20 ℃下,采取实验生态学方法,测定了10、15、20、25、30和35盐度下3 种规格[干质量(0.220±0.000)、(0.489±0.002) g和(0.899±0.002) g]可口革囊星虫的耗氧率和排氨率。试验结果表明,盐度、规格均显著影响可口革囊星虫的耗氧率和排氨率(P<0.05)。在盐度10~25时,3种规格可口革囊星虫的耗氧率和排氨率均随盐度增加而增大,盐度25时达到峰值后,随着盐度增大而减小。耗氧率和排氨率与盐度呈一元二次函数关系:y= ax2+bx+c。在相同盐度下,可口革囊星虫的耗氧率和排氨率随体质量的增大而显著减小(P<0.05),且耗氧率和排氨率与干质量符合y=amb模型。盐度10~35时,可口革囊星虫的氧氮比值为5.947~25.246,表明可口革囊星虫主要由蛋白质和脂肪提供能量。
可口革囊星虫;盐度;体质量;耗氧率;排氨率
呼吸和排泄是动物最基本的新陈代谢活动。研究新陈代谢可以了解动物自身的生理状态、代谢规律、营养状况以及对外界环境的适应能力[1]。耗氧率和排氨率是养殖生物呼吸代谢的最直观表达参数,受外部环境因子如光照、饵料、温度、盐度、pH 等和内在因子如体质量、发育期等的双重影响。盐度可改变动物的渗透压及物质和能量代谢,包括耗氧率和排氨率,影响水生动物的生存、生长、发育以及繁殖[2]。因此,研究盐度对海洋生物能量代谢的影响具有重要的理论和实际意义。
可口革囊星虫(Phascolosomaesculenta)是我国特有的动物[3],俗称“泥丁”、“泥蒜”,广泛分布于浙江、福建、广东、广西、海南和台湾等省区沿海。目前,部分学者在星虫养殖技术[4-6]、繁殖生物学[7]、胚胎及幼体发育[8]、受精过程[9]、生殖周期[10]、人工繁育[11]等方面已进行了一些研究,但有关生理生态学方面的研究尚未见报道。笔者于2015年研究了盐度对不同规格可口革囊星虫耗氧率和排氨率的影响,探索星虫的生理代谢规律,丰富生理生态学内容,为可口革囊星虫稚虫中间培育及人工增养殖发展提供参考。
1 材料与方法
1.1 材料
试验用可口革囊星虫采集于广西北海市闸口镇红树林,称量湿质量(精确至0.01 g)后,分为小规格组、中规格组和大规格组,在室内泥池暂养24 h(表1), 池内海泥厚2~5 cm,暂养期间不投饵料。试验开始时,徒手挖出可口革囊星虫冲洗干净,试验结束后清除肠道和肾管,置于烘箱内80 ℃烘干至恒定质量并称量。
试验海水为经沉淀、沙滤的自然海水,水温为18.5~20 ℃,盐度梯度为10、15、20、25、30和35,pH为7.8~8.2。
1.2 方法
盐度驯化期间持续少量添加淡水或高盐海水,保持每2 h降低或升高1个单位,到达指定盐度后暂养12 h后进行试验。
试验中,小规格组以250 mL三角烧瓶作为呼吸瓶,每瓶放5条可口革囊星虫;中规格组、大规格组均以1000 mL三角烧瓶作为呼吸瓶,分别放5条和2条可口革囊星虫。每一规格每一盐度设 5 个平行,外加一个空白对照,对照瓶内不放星虫,其他条件与平行样保持一致,装满对应梯度的海水后用塑料薄膜封口。试验采用静水方法,22 h后打开薄膜抽取水样。溶解氧和氨氮分别采用YSI Pro2030溶氧仪和法国Alliance-Futura/ Proxima连续流动分析仪进行测定。
采用水瓶法测定方格星虫水体中代谢瓶和对照瓶的溶解氧(DOt、DO0,μg/L)和氨氮(At、A0,μg/L)质量浓度。耗氧率和排氨率的计算公式为:

表1 试验可口革囊星虫的生物学数据
耗氧率/μg·(g·h)-1=(DO0-DOt)×V/(m·t)
排氨率/μg·(g·h)-1=(At-A0)×V/(m·t)
式中,DOt、DO0为方格星虫水体中代谢瓶和对照瓶的溶解氧(μg/L),At、A0为方格星虫水体中代谢瓶和对照瓶的氨氮质量浓度(μg/L),V为试验瓶的体积(L),m为体壁干质量(g),t为代谢时间(h)。
氧氮原子数比的计算公式如下:
氧氮比=(耗氧率/16)/(排氨率/14)
试验结果采用SPSS 17.0和EXCEL软件进行回归和方差分析,采用LSD方法进行比较检验,以P<0.05作为差异显著的标准。
2 结果与分析
2.1 盐度和规格对可口革囊星虫耗氧率的影响
盐度显著影响不同规格可口革囊星虫的耗氧率(P<0.05)(图1)。在各个盐度组中小规格组的可口革囊星虫的单位体质量耗氧率与其他盐度组存在显著差异(P<0.05);中规格组的可口革囊星虫在盐度15~30耗氧率无显著差异 (P>0.05),其他处理间均存在显著差异(P<0.05);在盐度10~35,大规格组的可口革囊星虫的耗氧率无显著差异(P>0.05),盐度15~30的耗氧率无显著差异 (P>0.05),其他盐度之间均存在显著差异(P<0.05)。
盐度10~25,3种规格可口革囊星虫的单位体质量耗氧率均随着盐度的上升而增大,在盐度25时达到最大值,之后随着盐度上升逐渐减小(图1)。可口革囊星虫耗氧率与盐度之间的拟合最符合一元二次方程:y=ax2+bx+c(a、b、c为常数,x为盐度,y为耗氧率)(表2)。
可口革囊星虫的耗氧率随体质量的增加而减小(表3)。相同盐度下,3种规格的耗氧率由高至低依次为:小规格>中规格>大规格,耗氧率和干质量之间的关系符合函数y=amb(a、b为常数,m为干质量,y为耗氧率)。

图1 盐度和规格对可口革囊星虫耗氧率的影响
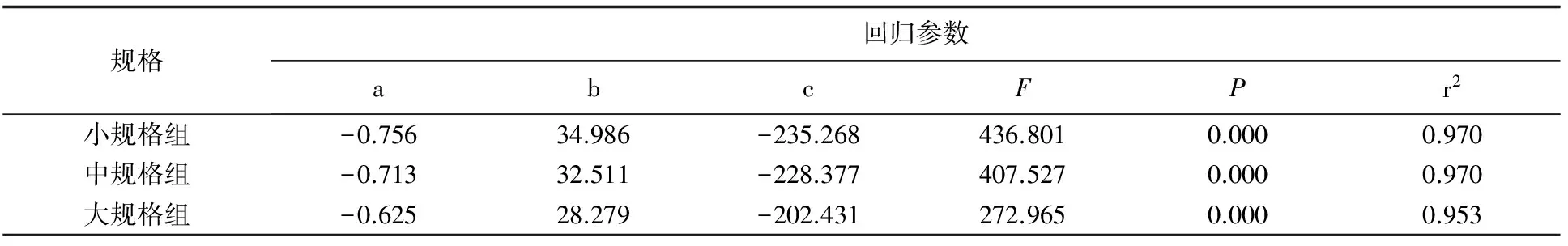
规格回归参数abcFPr2小规格组-0.75634.986-235.268436.8010.0000.970中规格组-0.71332.511-228.377407.5270.0000.970大规格组-0.62528.279-202.431272.9650.0000.953

表3 不同盐度下可口革囊星虫干质量和耗氧率的回归结果
2.2 盐度对不同规格可口革囊星虫排氨率的影响
盐度显著影响不同规格可口革囊星虫的排氨率(P<0.05)(图2)。各个盐度组,小规格组的可口革囊星虫的排氨率与盐度存在显著差异(P<0.05);盐度为10~35、15~30、20~25,中规格组的可口革囊星虫的排氨率无显著差异 (P>0.05);盐度为10~35,大规格组可口革囊星虫的排氨率无显著差异 (P>0.05),其他盐度处理之间均存在显著差异(P<0.05)。
盐度10~25,3种规格可口革囊星虫的单位体质量排氨率随盐度的上升而增大,在盐度25时达到最大值,之后随着盐度上升逐渐减小(图2)。可口革囊星虫排氨率与盐度之间拟合最符合一元二次方程:y=ax2+bx+c(a、b、c为常数,x为盐度,y为排氨率)(表4)。
在6个试验盐度下,不同规格的可口革囊星虫的排氨率随着体质量的增加而减小(表5)。相同盐度下,3种规格可口革囊星虫的排氨率为:小规格组>中规格组>大规格组,排氨率和干质量之间的关系符合函数y=amb(a、b为常数,m为干质量,y为排氨率)。

图2 盐度和规格对可口革囊星虫排氨率的影响

规格回归参数abcFPr2小规格组-0.0381.666-7.77949.7650.0000.793中规格组-0.0321.425-7.64953.1970.0000.816大规格组-0.0371.690-12.13241.3750.0000.754

表5 不同盐度下可口革囊星虫干质量与排氨率的回归结果
2.3 不同规格和盐度下可口革囊星虫的氧氮比
小规格组、中规格组、大规格组可口革囊星虫的氧氮比值分别为5.947~14.861、6.661~14.071和7.629~25.246(表6)。盐度15~30,3种规格的可口革囊星虫的氧氮比均呈现先升后降,在盐度25时达到最小值后又升高的规律。盐度10时,3种规格的氧氮比值为5.947~7.629,随着体质量的增加而增大,盐度35时,3种规格的氧氮比值为8.082~12.312,随着体质量的增加而减小,显示了不同规格的可口革囊星虫在不同盐度下的氧氮比。

表6 不同盐度和规格下可口革囊星虫的氧氮比
3 讨 论
3.1 可口革囊星虫与其他物种耗氧率的比较
水生动物代谢过程中的耗氧率、排氨率不仅受环境因子影响(如温度、盐度、pH等),而且与养殖生物本身的行为和生理特点等因素密切相关[12]。部分水产动物的耗氧率研究表明[13],水生动物的耗氧率差别较大,变化范围为 0.02 mg/(g·h)[犬齿牙鲆(Paralichthysdentatus)][14]至9.09 mg/(g·h)[栉孔扇贝(Chlamysfarreri)][15],并按耗氧率大小将水生生物分为低型[耗氧率最大值<1 mg/(g·h)]、中型[(耗氧率最大值为1~3 mg/(g·h)]和高型[耗氧率最大值>3 mg/(g·h)],其主要原因是维持水生生物生命活动的多种组织相对含量不同,推测了水生生物运动量大小影响耗氧率。本试验中,在不同盐度下可口革囊星虫的耗氧率为0.017~0.18 mg/(g·h),属于耗氧率低型。据测定,光裸星虫(Sipunculusnudus)的最大单位体质量耗氧率为0.479 mg/(g·h)[16],同属于耗氧率低型。光裸星虫钻沙速度很快,活动很敏捷,可口革囊星虫行动缓慢,活动范围小,不同的生活习性与这两个物种的最大单位体质量耗氧率有对应关系。笔者观察到,3种规格的可口革囊星虫在高盐和低盐时长期保持静止状态,不活动,吻部不伸出,而在盐度25时活动量最大,几乎所有个体的吻部均伸出互相缠绕作一团。这种生活习性与耗氧率和排氨率的变化规律相对应。推测可口革囊星虫的耗氧率除主要受其生物特异性影响外,运动量也是影响因素之一。可口革囊星虫这种耐低氧的生物特性,与它栖息的底质密切相关,为高密度人工养殖提供了必要条件。
3.2 盐度对可口革囊星虫耗氧率和排氨率的影响
盐度影响动物体的渗透压和新陈代谢[17]。盐度对水生生物耗氧率和排氨率的影响因物种特异性而有所不同。例如,鳗鲡(Anguillajaponica)仔鱼[18]随盐度升高,耗氧率逐渐降低;中华绒螯蟹(Eriocheirsinensis)亲蟹[19]在盐度0~24内,耗氧率和排氨率均随着盐度升高而降低;在一定的盐度范围内,日本黄姑鱼(Nibeajaponica)[20]、东风螺(Babyloniaareolata)[21]的耗氧率和排氨率随盐度的升高而升高,达到峰值后,随盐度的继续升高而降低。
在盐度5和40时,可口革囊星虫(规格为2.6~3.2 g)的一周成活率分别为90.6%和92.8%;而在10~35内,一周存活率可达97.7%~99%[22]。结合本试验可知,在盐度10~35范围内对可口革囊星虫的存活率无显著影响,但显著影响呼吸和排泄。本试验中,盐度和耗氧率、盐度和排氨率的关系均为一元二次方程,这与日本黄姑鱼、东风螺相似。3种规格可口革囊星虫的耗氧率和排氨率的峰值均在盐度25时,这表明盐度25是可口革囊星虫的最佳代谢盐度。笔者认为其原因可能是,在盐度25时可口革囊星虫只需消耗一小部分能量即能保持体内外渗透压的平衡,实际观察到此盐度下可口革囊星虫的活动量最大,所以耗氧率和排氨率均最大;高于或低于适应的海水盐度范围时,其减少活动量、降低耗氧率和排氨率来适应环境的渗透压变化。数据显示,3种规格的可口革囊星虫都能调节自身的生理状态适应盐度的变化,尤其是小规格组的耗氧率和排氨率在各个盐度梯度都有显著差别,说明小规格可口革囊星虫对盐度变化有更加强大的调节能力。
3.3 氧氮比
氧氮比是指生物体对蛋白质、脂肪和碳水化合物等能源物质分解代谢所消耗的氧原子和排出的氮原子之间的比率,指示了机体代谢过程中的能量来源,受环境压力(温度、盐度、pH等)影响。如果完全由蛋白质提供能量,氧氮比约为7[23], 如果是蛋白质和脂肪氧化供能,氧氮比约为24 ,如果主要由脂肪或碳水化合物供能,氧氮比将变无穷大[24]。本试验中,小规格组、中规格组、大规格组可口革囊星虫的氧氮比为5.947~25.246。在本试验的盐度范围内,可口革囊星虫主要由蛋白质和脂肪提供能量,蛋白质比例较大。在较适宜的盐度条件下(15~30),3种规格可口革囊星虫的氧氮比随盐度的增加而降低,在最适盐度25时,氧氮比最低,然后随盐度增加而增大,这与方格星虫的氧氮比变化规律相似(待发表)。据此推测,在盐度15~25范围内,可口革囊星虫体内的代谢底物——蛋白质的比例可能随着盐度的增加而增大,脂肪的比例减小;在盐度25~30内,代谢底物蛋白质减少而脂肪增多。本试验中,盐度15时大规格组的氧氮比明显偏高,可能是大规格组受盐度影响较大,数据出现较大偏差。
4 结 论
可口革囊星虫的最适生长盐度为25,3种规格(特别是小规格)对盐度有很强的适应调节能力。在人工养殖条件下,盐度25时,可加大投喂量,促进可口革囊星虫的生长;盐度10~35范围内均不会影响小规格可口革囊星虫的生存。本试验设定的最小盐度梯度为10,但在雨季,海水实际盐度有可能低于10。关于可口革囊星虫的盐度下限以及代谢变化,有待进一步研究。
[1] Lee H, Lee B P, Messersmith P B, et al. A reversible wet/dry adhesive inspired by mussels and geckos[J].Nature,2007,448(7151):338-341.
[2] 李超伦,王克.植食性浮游桡足类摄食生态学研究进展 [J].生态学报,2002,22(4):593-596.
[3] 周红,李风鲁,王玮,等.中国动物志[M].北京:科学出版社,2007:53-77.
[4] 丁理法,徐敏娴,陈飞,等.不同施肥和投饵方式对可口革囊星虫养成结果的影响试验[J].科学养鱼,2015(8):69-70.
[5] 丁理法,竺俊全,叶荣华,等.可口革囊星虫人工养殖试验[J].水产养殖,2004,25(5):24-28.
[6] 刘瑞义.可口革囊星虫养殖技术要点[J].科学养鱼,2006(4):34-35.
[7] 张虹,曾达,艳华.可口革囊星虫繁殖生物学研究概况[J].大众科技, 2015,17(4):114-115.
[8] 吴洪喜,应雪萍,陈琛,等.可口革囊星虫胚胎与幼体的发育[J].动物学报,2006,52(4):765-773.
[9] 竺俊全,王武,应雪萍,等.可口革囊星虫受精过程及早期卵裂的细胞学变化[J]. 动物学报,2008,54(2):290-298.
[10] 陈慧,林国文,陈武,等.可口革囊星虫生殖周期的观察[J].海洋渔业,2009,31(2):139-145.
[11] 吴洪喜,陈琛,曾国权,等.可口革囊星虫人工繁育试验[J].海洋科学,2010,34(3):21-25.
[12] 刘鹏,宋协法,葛长字,等.温度/体重和昼夜节律对大菱鲆幼鱼代谢的影响[J] .河北渔业,2009(5):1-27.
[13] 邱成功,徐善良,林少珍,等. 不同温度条件下褐菖鲉幼鱼的耗氧率和排氨率[J] .应用海洋学学报,2014,33(1):84-89.
[14] 王波,李继强,曹志海,等.犬齿牙鲆幼鱼呼吸耗氧率的研究[J].海洋科学进展,2003,21(3):325-330.
[15] 王俊,姜祖辉,唐启升,等.栉孔扇贝耗氧率和排氨率的研究[J].应用生态学报,2002,13(9):1157-1160.
[16] 逯云召,王庆恒,杜晓东,等.温度和体质量对不同规格光裸星虫耗氧率和排氨率的影响[J].广东海洋大学学报,2012,32(1):87-91.
[17] Tiffany D T, John M L. The effect of salinity on respiration, excretion, regeneration and production inOphiophragmusfilograneus[J]. Journal of Experimental Marine Biology and Ecology,2002,275(1):1-14.
[18] 王广军,谢骏,潘德博.日本鳗鲡初孵仔鱼耗氧率的初步研究[J].海洋水产研究,2001,22(1):52-55.
[19] 庄平,贾小燕,冯广朋,等.盐度对中华绒螯蟹雌性亲蟹代谢的影响[J].中国水产科学,2012,19(2):217-222.
[20] 柴学军,胡则辉,徐君卓,等.盐度和pH 对日本黄姑鱼幼鱼耗氧率和排氨率的影响[J].浙江海洋学院学报:自然科学版,2009,28(2):146-150.
[21] 刘建勇,绍杰,卓健辉,等.盐度对东风螺耗氧率和排氨率的影响[J].热带海洋学报,2005,24(4):35-40.
[22] 陈慧,陈武,林国文,等.可口革囊星虫对盐度耐受力的影响[J].福建水产,2006,12(4):74-76.
[23] Mayzalld P.Respiration and nitrogen excretion of zooplankton Ⅳ.The influence of starvation on the metabolism and biochemical composition of some species[J].Mar Biol,1976,37(1):47-58.
[24] Ikeda T.Nutrition ecology of marine zooplankton[J].Mem Fac Fish Res,1974,22(1):1-77.
EffectsofSalinityandBodyWeightonOxygenConsumptionandAmmoniaExcretionofSipunculaPhascolosomaesculenta
ZENG Mengqing,JIANG Yan,YOU Chuchao,LIU Xujia,LIU Haijuan
( Guangxi Key Laboratory of Marine Biotechnology, Guangxi Institute of Oceanology, Beihai 536000, China )
The effects of salinity(10, 15, 20, 25, 30 and 35)and body weight [dry body weight of (0.220± 0.000) g, (0.489±0.002) g and (0.899±0.002) g]on oxygen consumption (OR) and ammonia excretion (AR) of sipunculaPhascolosomaesculentawere studied at water temperature of 18.5—20 ℃ by experimental ecology methods. The results showed that salinity had significant effect on OR and AR (P<0.05), enhanced both OR and AR with salinity increment, and the peak at a salinity of 25, then decreased rapidly with salinity increasing. The relationship between OR and salinity was represented by the quadratic equationy=ax2+bx+c, and the relationship between AR and salinity was the similar with relationship between OR and salinity. The oxygen consumption and ammonia excretion rates were shown to be negatively correlated with its body weight in sipuncula. There were significant difference in OR and AR among the three size groups (P<0.05), and their relationship was expressed by power function (y=amb).The O/N values were found to be ranged from 5.947 to 25.246 within a salinity of 10 to 35, indicating that the sipuncula consumed fat or protein as the major energy sources.
Phascolosomaesculenta; salinity; body weight; oxygen consumption rate; ammonia excretion rate
10.16378/j.cnki.1003-1111.2017.05.018
2016-08-19;
2016-10-18.
广西科学研究与技术开发计划项目(桂科攻14121006-2-3);广西培养新世纪学术和技术带头人专项资金资助项目.
曾梦清(1986-),女,助理研究员;研究方向:水生动物繁育技术.E-mail:86mengqing@163.com.通讯作者:蒋艳(1980-),女,副研究员;研究方向:海水新品种繁育技术.E-mail:hdjy98@163.com.
S968.9
A
1003-1111(2017)05-0647-05
DOI:10.16378/j.cnki.1003-1111.2017.05.019
