北方须鳅的耗氧节律、耗氧率和窒息点的研究
2018-06-04王晓光王兴兵寇凌霄石俊艳肖祖国魏宏祥骆小年
王晓光,王兴兵,寇凌霄,石俊艳,肖祖国,魏宏祥,刘 勇,李 军,骆小年
( 1.辽宁省淡水水产科学研究院,辽宁省水生动物病害防治重点实验室,辽宁 辽阳 111000;2.大连海洋大学 水产与生命学院,辽宁 大连 116023 )
北方须鳅(Barbatulabarbatulanuda)属硬骨鱼纲、鲤形目、鳅科、条鳅亚科,在我国东北地区、新疆北部以及俄罗斯、日本和朝鲜均有分布[1]。北方须鳅栖息于清冷水体中沙石底质处,不集群,通常晴朗天离开卵石作近距离觅食游动,主要食物为底栖无脊椎动物。该鱼肉质细嫩味美,深受百姓喜食,具有一定的经济价值。近年来,由于水利工程建设、环境污染以及过度捕捞等,北方须鳅栖息地缩减,生活环境破坏严重,种群数量锐减,资源量明显下降,建立保护区,开展北方须鳅生物学和人工增养殖技术研究具有重要的意义。
鱼类的耗氧率直接或间接地反映出鱼类新陈代谢规律、生理和生存状况,是有氧代谢强度的重要指标之一[2],窒息点则是鱼类忍受低溶解氧的极限指标。水温是鱼类生存的重要环境因子之一,影响鱼的新陈代谢强度、活动和耗氧率及窒息点。了解及掌握北方须鳅的耗氧率和窒息点,对其生理学研究具有重要的理论意义,对其增养殖和保护有应用价值[3]。目前,关于北方须鳅地理种群的形态[4]、线粒体基因组序列及鳅科鱼类系统发育分析[5]等研究已有报道,但尚未见北方须鳅耗氧率和临界窒息点等相关研究。笔者研究了温度对不同体质量北方须鳅耗氧率和临界窒息点的影响及耗氧节律,旨在了解北方须鳅的新陈代谢规律、生理和生活状况,为其生理学研究和健康增养殖、活体运输以及生态保护等提供参考。
1 材料与方法
1.1 材料
北方须鳅地笼网捕获于辽宁省辽阳县下达河,在生态室暂养7 d。暂养期间,北方须鳅放养在60 cm×40 cm×50 cm底部布有鹅卵石的水族箱中,以逐渐适应小水体的试验环境,不投喂。暂养后,选择体质健康的北方须鳅,称量体质量,测定全长和体长。按体质量将北方须鳅分为(1.88±0.33) g、 (4.24±0.61) g和 (7.27±0.76) g 3种规格(表1)。3种规格北方须鳅的体质量、全长和体长形态性均呈极显著性差异(P<0.01)。试验用水为曝气5 d以上的自来水,pH 7.2±0.2,溶解氧(8.0±0.2) mg/L。

表1 北方须鳅的生物学数据
注:标有不同大写字母的平均值间差异极显著(P<0.01).
1.2 方法
1.2.1 北方须鳅24 h耗氧率的变化
采用流水呼吸室法[6]测定北方须鳅24 h耗氧率,(7.27±0.76) g和(4.24±0.61) g试验鱼呼吸室的体积为5 L,分别放鱼40 尾和50 尾,(1.88±0.33) g试验鱼的呼吸室体积为2.5 L,放鱼50 尾,设置平行组。试验鱼放入呼吸室后,调节水流量,使出水溶解氧>6.0 mg/L,水温(15±0.5) ℃;鱼在呼吸室内适应1.0~1.5 h,再调节水流量,使出水溶解氧约为4.0 mg/L,待鱼处于自然平稳状态时,开始试验,连续测量24 h,每2 h测定一次水温、单位时间水流量和进、出呼吸室水的溶解氧质量浓度[7]。耗氧率(mg/g·h)按下式计算:
Or= (A1-A2)×S/m
式中,Or为耗氧率;A1为进水溶解氧质量浓度(mg/L);A2为出水溶解氧质量浓度(mg/L);S为单位时间的流量(L/h);m为试验鱼体总质量(g)。
试验期间避免阳光直射,保持室内安静,为减少灯光照射对鱼体的干扰,18:00至次日6:00呼吸室用不透光的黑塑料包裹。
1.2.2 水温对耗氧率和窒息点影响的测定
采用静水呼吸室法测定水温对耗氧率和窒息点影响。水温设0、2、5、10、15、20、25、27、30 ℃ 9个梯度,误差在±0.5 ℃以内。(7.27±0.76) g和(4.24±0.61) g北方须鳅每组10 尾,(1.88±0.33) g北方须鳅每组20 尾,设定平行组。试验前采用逐级升温或降温的方法,0 ℃试验时间为1.5 h,2、5、10、15、20 ℃为2 h,25、27、30 ℃为1 h。待北方须鳅处于自然平稳状态后,试验开始,封闭进、出水。试验结束后计算试验前后呼吸室耗氧量和耗氧率。
上述不同温度静水试验结束后,再次封闭进、出水,密封呼吸室,待50%试验鱼失去平衡、昏迷或濒临死亡时试验结束,此时呼吸室内溶解氧质量浓度为该温度下的临界窒息点。
耗氧率(mg/g·h)的计算公式为:
Or=(B1-B2)×V/(m×t)
式中,Or为耗氧率;B1和B2为试验前、后溶解氧质量浓度(mg/L);V为呼吸室水体积(L);t为试验时间(h);m为试验鱼总体质量(g)。
1.2.3 数据的统计和分析
试验数据用平均值±标准差表示,用 Excel 和 SPSS 16.0软件进行统计分析,用方差分析来检验昼夜和体质量对北方须鳅耗氧率的影响显著性,显著水平为0.05,极显著水平为0.01;用Pearson 相关分析,分析耗氧率与水温相关性;用 Excel软件确定体质量与北方须鳅耗氧率呈幂函数关系。
2 结果与分析
2.1 北方须鳅24 h耗氧率变化
(1.88±0.33) g北方须鳅24 h耗氧率波动较大:6:00—18:00,耗氧率逐渐升高,18:00达最高值[(0.166±0.028) mg/(g·h)];而后耗氧率逐渐下降,次日06:00耗氧率达最低值[(0.080±0.012) mg/(g·h)]。(1.88±0.33) g北方须鳅在白天(8:00—16:00)耗氧率为(0.131±0.023) mg/(g·h),夜间(18:00—6:00)为[(0.124±0.029) mg/(g·h)],白天耗氧率略高于夜间 (P>0.05)。而在12:00—20:00出现耗氧率高峰区域,平均值为 [(0.151±0.016) mg/(g·h)),极显著高于22:00—次日10:00[(0.110±0.016) mg/(g·h)](P<0.01)。
(4.24±0.61) g北方须鳅24 h耗氧率波动较小: 凌晨06:00达最低值[(0.073±0.006) mg/(g·h)],随后逐渐升高,在12:00达最高值[(0.141±0.021) mg/(g·h)];达到最高值后,耗氧率逐渐降低;在16:00和20:00分别出现两个峰值,其值分别为[(0.123±0.011) mg/(g·h)]和[(0.124±0.014) mg/(g·h)]。(4.24±0.61) g北方须鳅白天 (8:00—16:00) 的耗氧率[(0.107±0.024) mg/(g·h)]略高于夜间(18:00—6:00) [(0.098±0.015) mg/(g·h)] (P>0.05);而在12:00—20:00耗氧率[(0.114±0.023) mg/(g·h)]明显高于22:00—次日10:00[(0.093±0.010) mg/(g·h)],差异显著 (P<0.05)。
(7.27±0.76) g北方须鳅24 h耗氧率曲线平直,波动性较小: 10:00耗氧率[(0.080±0.010) mg/(g·h)]最低,12:00耗氧率[(0.139±0.017) mg/(g·h)]最高;白天(8:00—16:00)的耗氧率[(0.102±0.023) mg/(g·h)]略高于夜间(18:00—6:00) [(0.094±0.008) mg/(g·h)] (P>0.05);12:00—20:00耗氧率[(0.109±0.017) mg/(g·h)]与22:00—次日10:00耗氧率[(0.088±0.007) mg/(g·h)]差异显著 (P<0.05)(图1)。
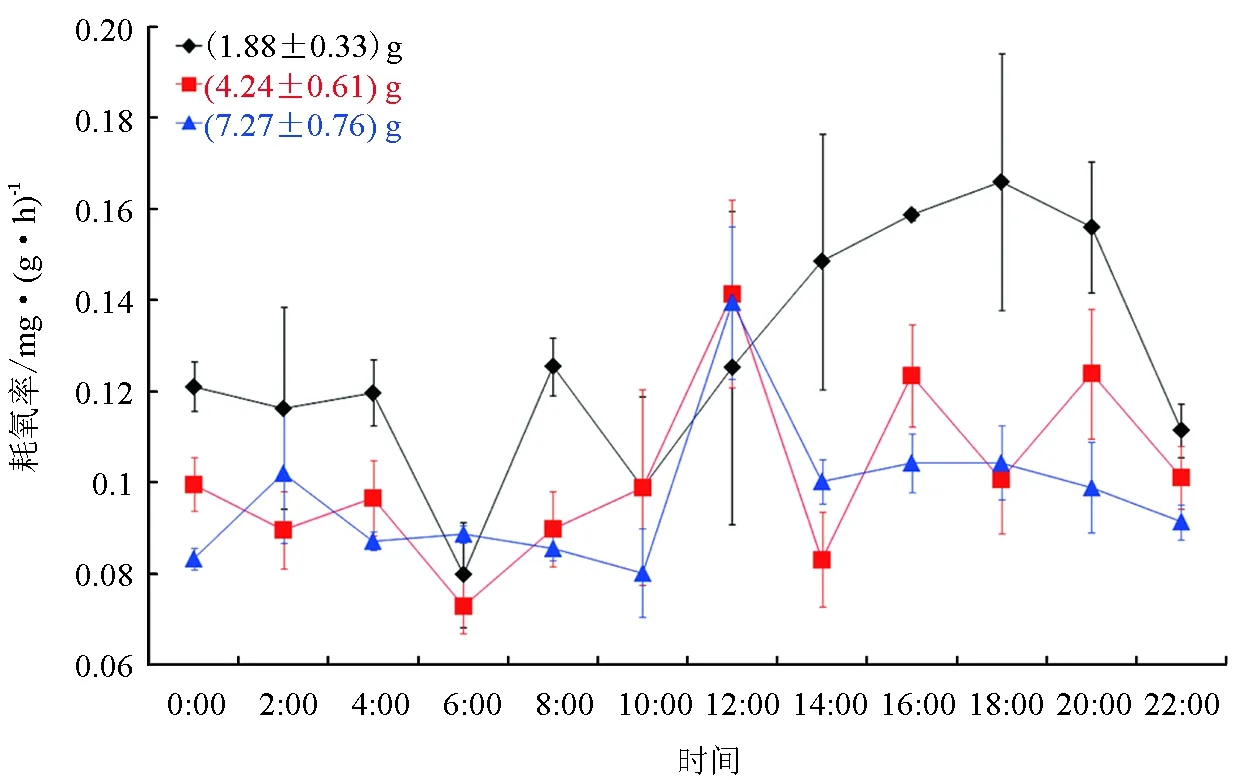
图1 不同规格北方须鳅24 h耗氧率
2.2 水温对北方须鳅耗氧率影响
3种规格北方须鳅的耗氧率均随着水温(2~27 ℃)的升高而逐渐增加,且增加趋势基本一致,表明北方须鳅的耗氧率与水温关系密切,呈正相关(P<0.01)(图2)。
2 ℃时,3种规格北方须鳅的耗氧率最小,分别为(0.045±0.009) mg/(g·h)、(0.049±0.008) mg/(g·h)、(0.046±0.004) mg/(g·h),耗氧率相近;水温2~15 ℃时,(1.88±0.33) g北方须鳅的耗氧率变化较小(斜率为0.0054),耗氧率为0.045~0.114 mg/(g·h);而在15~27 ℃时,耗氧率变化大(斜率为0.0222),高于2~15 ℃间耗氧率的变化,耗氧率为0.114~0.385 mg/(g·h)。
水温2~10 ℃时,(4.24±0.61) g北方须鳅的耗氧率变化较小(斜率为0.0026),为0.049~0.070 mg/(g·h);10~25 ℃时,耗氧率为0.070~0.209 mg/(g·h),耗氧率变化量较大(斜率为0.0093),高于2~10 ℃水温时的变化;而在25~27 ℃时,耗氧率陡然增加,由0.209 mg/(g·h)增至0.401 mg/(g·h),增加近一倍,变化量最大。
水温2~15 ℃时,(7.27±0.76) g北方须鳅的耗氧率变化较小(斜率为0.0055),为0.046~0.115 mg/(g·h);15~25 ℃时,耗氧率为0.115~0.245 mg/(g·h),耗氧率变化量较大(斜率为0.013),高于2~15 ℃时的耗氧率变化;25~27 ℃,由0.245 mg/(g·h)增至0.365 mg/(g·h),变化量大。
试验中还测定了3种规格北方须鳅在水温0 ℃和30 ℃时的耗氧率。在0 ℃时,(1.88±0.33) g、(4.24±0.61) g和(7.27±0.76) g北方须鳅的耗氧率分别为(0.089±0.023) mg/(g·h)、(0.082±0.027) mg/(g·h)和(0.046±0.021) mg/(g·h),其中(1.88±0.33) g和(4.24±0.61) g北方须鳅耗氧率高于在2 ℃和5 ℃耗氧率,与其在10 ℃时耗氧率相近。同时观察到北方须鳅呼吸频率加快,远远高于其在水温2~27 ℃中的呼吸频率;而(7.27±0.76) g北方须鳅在0 ℃水温中的耗氧率与其在2 ℃耗氧率相近,呼吸频率正常。在30 ℃水温中,3种规格北方须鳅的呼吸加快,剧烈游动,并伴有窜出水面的现象,同时部分鱼死亡。
2.3 体质量对北方须鳅耗氧率的影响
不同体质量对北方须鳅耗氧率的影响见图3。由图3可见,(1.88±0.33) g、(4.24±0.61) g和(7.27±0.76) g北方须鳅24 h耗氧率分别为(0.127±0.026) mg/(g·h)、(0.102±0.019) mg/(g·h)和(0.097±0.016) mg/(g·h),其中(1.88±0.33) g北方须鳅耗氧率最大,其次为(4.24±0.61) g规格,(7.27±0.76) g规格耗氧率最小。方差分析表明,(1.88±0.33) g北方须鳅耗氧率分别与(7.27±0.76) g和(4.24±0.61) g规格耗氧率差异极显著(P<0.01),而(4.24±0.61) g北方须鳅24 h耗氧率与(7.27±0.76) g北方须鳅24 h耗氧率差异不显著(P>0.05)。北方须鳅24 h耗氧率随体质量的增加而下降,符合y=0.1426x-0.2055(r2=0.9414)关系。
2.4 温度对北方须鳅窒息点的影响
3种规格试验鱼的临界窒息点随温度升高,呈“V”字型先降后升的波动规律,波动规律基本一致(图4)。在2~10 ℃时,随着水温的上升,北方须鳅临界窒息点下降,3种规格试验鱼均在10 ℃时出现最低临界窒息点,(1.88±0.33) g、(4.24±0.61) g和(7.27±0.76) g北方须鳅最低临界窒息点分别为(0.352±0.047) mg/L、(0.298±0.047) mg/L和(0.430±0.050) mg/L。出现最低临界窒息点后,3种规格北方须鳅窒息点随水温的升高(15~27 ℃)逐渐上移。
观察发现,随着水温逐渐升高,北方须鳅呼吸频率逐渐加快。5 ℃时(4.24±0.61) g北方须鳅的呼吸频率为(84±4)次/min,20 ℃时呼吸频率增至(168±3)次/min,增加近一倍,窒息时间随温度升高而缩短。3种规格北方须鳅昏迷或濒临死亡的过程基本一致。当溶解氧充足时(封闭时溶解氧>3.0 mg/L),北方须鳅在呼吸室内底部静卧,处于自然状态,随时间增加,密闭呼吸室内溶解氧减少,北方须鳅呼吸频率由快变慢,上下蹿动,烦躁不安,随溶解氧的进一步减少,鱼体游动失去平衡,濒临死亡,同时体色变浅,花纹更加明显,腹部苍白。
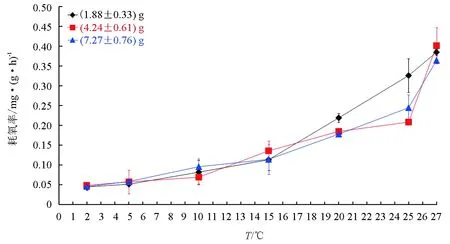
图2 2~27 ℃时北方须鳅的耗氧率
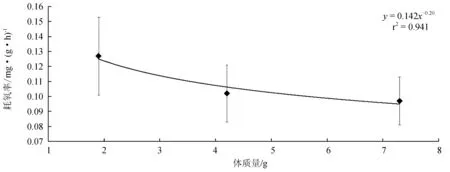
图3 不同体质量北方须鳅的24 h耗氧率
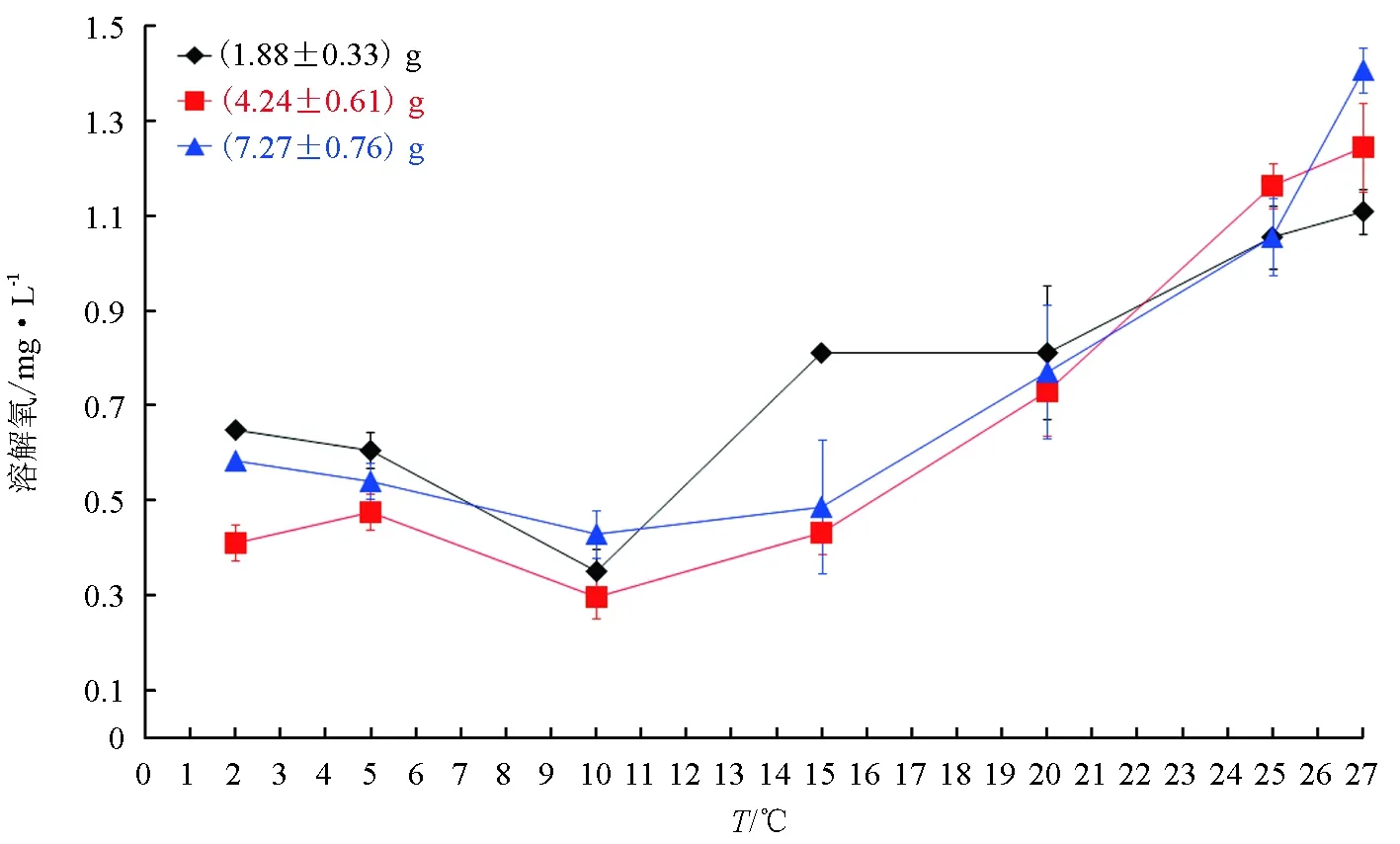
图4 北方须鳅在水温2~27 ℃下的窒息点
3 讨 论
3.1 昼夜变化对北方须鳅耗氧率的影响
耗氧率变化可直接或间接地反映鱼类的代谢规律、生理以及鱼类活动周期[8],如鱼类在进食或活动旺盛时期,耗氧量大。研究表明,鱼类耗氧率的昼夜节律可分为:(1)白天耗氧率高于夜间耗氧率,如麦穗鱼(Pseudorasboraparva)[9]和浅色黄姑鱼(Nibeacoibor)[10]等;(2)夜间耗氧率高于白天耗氧率,如鸭绿沙塘鳢(Odontobutisyaluensis)[11]和雷氏七鳃鳗(Lampetrareissneri)[12]等;(3)昼夜耗氧率无差异,白天和夜晚均有耗氧高峰期与低峰期,如鲇鱼(Silurusasotus)[13];(4)某一时间段内出现耗氧率高峰区域,如鲻鱼(Mugilcephalus)幼鱼[14]。本试验结果表明,北方须鳅昼夜24 h耗氧率与鲻鱼幼鱼具有一定的相似性,在12:00—20:00的耗氧率较高(P<0.05),说明北方须鳅在12:00—20:00呼吸代谢旺盛,活动频繁,可能与捕食有关。自然条件下,中午至傍晚溪水温度较高,溪水高温时间段与北方须鳅呼吸代谢旺盛时间段基本相同(12:00—20:00),此时水体中昆虫幼虫、甲壳类动物和寡毛类丰富,北方须鳅捕食。
北方须鳅白天耗氧率均高于夜间,与长薄鳅(Leptobotiaelongata)[18]、叶尔羌高原鳅[Triplophysa(Hedinichthys)yarkandensis][19]、似鲇高原鳅(T.siluorides)[20]、红尾副鳅(Paracobitisvariegatus)[21]、大鳞副泥鳅(Paramisgurnusdabryanus)[22]和泥鳅 (Misgurnusanguillicaudatus)[23]耗氧率的昼夜规律不同,但北方须鳅最高耗氧率和最低耗氧率的出现时间,与其他鳅都在同一时间段。本试验中,(1.88±0.33) g和(4.24±0.61) g北方须鳅均在06:00耗氧率最低,这与似鲇高原鳅[20](8:00耗氧低峰值)、大鳞副泥鳅[22][(1.88±0.33) g规格在3:00出现耗氧低峰值, (7.27±0.76) g在7:00出现耗氧低峰值]和泥鳅[23](在6:00出现耗氧低峰值) 的最低耗氧率时间相近;(7.27±0.76) g北方须鳅在10:00耗氧率最低,与红尾副鳅[21](在12:30出现耗氧低峰值) 出现最低耗氧率的时间相近。(1.88±0.33) g北方须鳅在18:00耗氧率最高,与似鲇高原鳅[20](20:00 耗氧率最高)和泥鳅[23](耗氧率最高点均出现在18:00) 出现最高耗氧率的时间相近。与其他鳅科鱼类相比,北方须鳅出现最高和最低耗氧率的时间相近,说明鳅科鱼类生活习性相近,但北方须鳅有独特的生活习性,这可能与其生活的地理环境有关。
3.2 温度对北方须鳅耗氧的影响
水温与鱼类耗氧率之间密切相关[11-12,24]。在适温范围内随水温升高,鱼体内各种酶活性增强,基础代谢旺盛,耗氧量上升,影响鱼体的新陈代谢[25]。本研究表明,水温为2~27 ℃,3种规格的北方须鳅耗氧率均随着水温的升高逐渐增加,耗氧率与水温关系密切(P<0.01),与长薄鳅[18]、大鳞副泥鳅[22]和泥鳅[26]耗氧率与水温关系一致。
耿龙武等[27]将温度对鱼类耗氧率的影响分为低温升高期、适温平稳期和温度改变期,与本试验结果中温度对北方须鳅耗氧率的影响相似。当温度低于5 ℃时,北方须鳅耗氧率低,耗氧率变化幅度小,随温度的降低耗氧率下降,其活动能力下降。温度的降低使维持鱼体内代谢能力下降,各种细胞酶活性减弱,基础代谢周期相对延长[28];水温为10~15 ℃时,(1.88±0.33) g和(7.27±0.76) g北方须鳅的耗氧率增加量较小,而(4.24±0.61) g北方须鳅在5~10 ℃时耗氧率增加量较小。水温超过15 ℃,(1.88±0.33) g、(4.24±0.61) g和(7.27±0.76) g北方须鳅的耗氧率相对增加量均较大。水温为0 ℃,(1.88±0.33) g和(4.24±0.61) g北方须鳅耗氧率异常增高,呼吸频率加快,推测可能由于水温过低,北方须鳅处于一种应激状态。而(7.27±0.76) g北方须鳅抗应激能力强,其耗氧与水温2 ℃的耗氧相近,呼吸频率正常。水温30 ℃,3种规格的北方须鳅均表现出不适现象,部分死亡,这可能是温度过高所致,在北方须鳅养殖过程中应注意高温对其危害。
邹桂伟等[18,29]认为,某一温度范围内,随温度升高,鱼类耗氧率和标准代谢率增加幅度相对较小,表明鱼类新陈代谢的异化作用上升较少,该温度范围为鱼类生长的适宜温度。水温为10~15 ℃,北方须鳅在呼吸室内游动正常,随温度升高而耗氧率变幅相对较小,推断北方须鳅适宜在此温度区域内生长和生活,是该鱼生长适宜温度范围。
3.3 体质量对北方须鳅耗氧率的影响
鱼类体质量与耗氧量呈正相关,与耗氧率呈负相关[30]。鱼体直接维持生命的多种组织(肾脏、脑、鳃等)每分钟耗氧量较高,而非直接维持生命的多种组织(骨骼、肌肉等)耗氧量较低,两类组织在鱼的生长过程中所占比例不同。幼鱼正处于生长发育时期,第一类组织所占比例较高,而随鱼体发育,第二类组织相对增长较快,因而耗氧率随个体体质量增加而下降。邹桂伟等[18]研究表明,长薄鳅耗氧率随体质量增加而逐渐降低,二者成明显的负相关。杜宗君等[21]研究结果表明,红尾副鳅耗氧率随着体质量的增大而降低。本研究表明,北方须鳅24 h耗氧率随体质量的增加而下降,呈负相关,这与鳅科鱼类以及其他鱼类具有一定的相似性。
3.4 温度对窒息点影响
王晓光等[12]研究了雷氏七鳃鳗窒息点与温度的关系,水温在1~20 ℃,3种规格的雷氏七鳃鳗窒息点均为先降后升,北方须鳅的临界窒息点随温度升高也先降后升。3种规格的北方须鳅均在水温10 ℃时出现最低临界窒息点,说明水温10 ℃时北方须鳅生理机能处于最佳状态,体质最强、抵抗力最强[26,31]。水温在10~27 ℃时,北方须鳅的临界窒息点随温度升高而升高,这与叶尔羌高原鳅[31]在水温20~29 ℃的窒息点具有相同的趋势。其他鱼类中,水温20~30 ℃的窒息点也随温度升高而升高,如豹纹鳃棘鲈(Plectropomusleopardus)、驼背鲈(Cromileptesaltivelis)、棕点石斑鱼(Epinephelusfuscoguttatus)、赤点石斑鱼(E.akaara)和珍珠龙胆石斑鱼(E.fuscoguttatus♀×E.lanceolatu♂)[32]。这可能是由于水温升高,鱼体内各种细胞酶活性增强,代谢旺盛所致[24]。
在相近的水温,北方须鳅的窒息点(水温5 ℃)高于叶尔羌高原鳅 (水温6.5~7.5 ℃)[19];在水温15 ℃时窒息点低于似鲇高原鳅(水温13 ℃)[20]和红尾副鳅[水温(14±0.5) ℃][21];在25 ℃时,高于大鳞副泥鳅(水温25 ℃)[22]和长薄鳅的窒息点(水温22~23 ℃)[18]。影响鱼类窒息点的因素很多,除鱼类种类差异外,还包括体质量、温度、二氧化碳分压和pH等因素[21]。在北方须鳅的养殖中应根据鱼体质量以及环境因子变化,增加水体中的溶解氧,保证正常生理活动与生长(表2)。
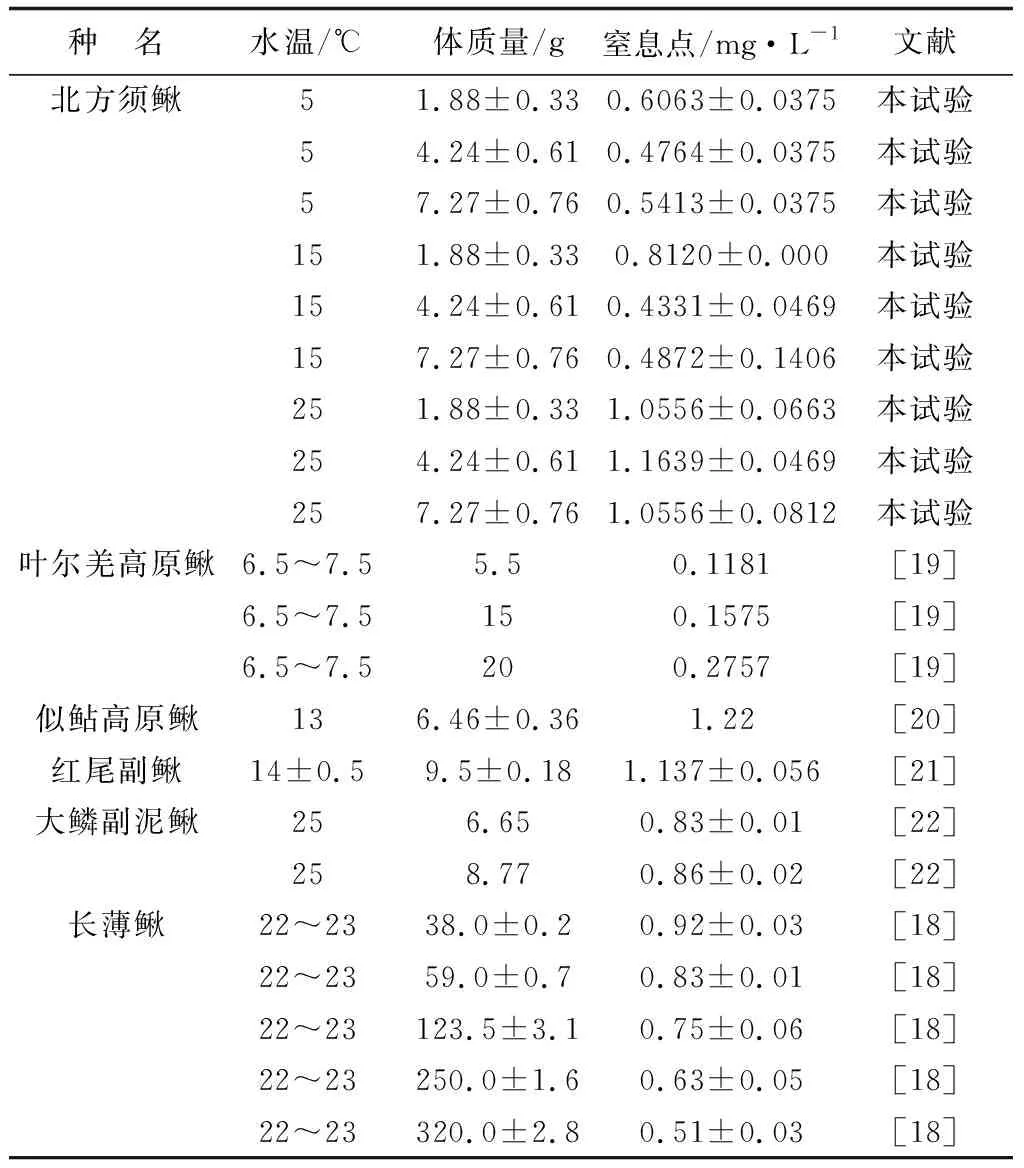
表2 北方须鳅与其他鳅科鱼类窒息点的比较
参考文献:
[1] 解玉浩,李文宽,解涵.东北地区淡水鱼类[M].沈阳:辽宁科学技术出版社, 2007:235-240.
[2] Modlin R F, Froelich A J. Influence of temperature, salinity, and weight on the oxygen consumption of a laboratory population ofAmericamysisbahia (Mysidacea)[J]. Journal of Crustacean Biology, 17(1):21-26.
[3] 殷名称.鱼类生态学[M].北京:中国农业出版社,1995:91-97.
[4] 孙鹏,马波,赵文阁. 北方须鳅4个地理种群的形态差异与判别分析[J].水产学杂志,2014,27(2):14-19.
[5] 赵习.北方须鳅线粒体基因组序列测定及鳅科鱼类系统发育分析[D].武汉:华中农业大学,2014.
[6] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会.GB/T 18654.8—2008,养殖鱼类种质检验,第8部分:耗氧率与临界窒息点的测定[S].北京:中国标准出版社,2008.
[7] 国家环境保护局.GB/T 7489—1987,水质溶解氧的测定碘量法[S].北京:中国标准出版社,1987.
[8] Clausen R G. Oxygen consumption in freshwater fishes[J].Ecology,1936,17(2):216-226.
[9] 杨坤,祝东梅,王卫民.麦穗鱼耗氧率和窒息点的测定[J].水产科学,2013,32(5):256-260.
[10] 李加儿,刘士瑞,区又君,等.浅色黄姑鱼幼鱼耗氧率、排氨率及窒息点的初步研究[J].海洋学报:中文版, 2008, 30(5):165-170.
[11] 李军,骆小年,徐忠源,等.鸭绿沙塘鳢耗氧率、氨氮排泄率与临界窒息点研究[J].上海海洋大学学报, 2010, 19(5):685-691.
[12] 王晓光,于伟君,李军,等.雷氏七鳃鳗耗氧率和窒息点的研究[J].大连海洋大学学报,2011,26(2):119-125.
[13] (英)勃朗M E编著,费鸿年译. 鱼类生理学[M].北京:科学出版社,1962:82-105.
[14] 李加儿,曹守花,区又君,等.鲻幼鱼耗氧率、排氨率和窒息点的研究[J].四川动物, 2014,33(3):434-439.
[15] Muir B S, Niimi A J. Oxygen consumption of the euryhaline fish aholehole (Kuhliasandvicensis) with reference to salinity, swimming and food consumption[J]. J Fish Res Board Can, 1972(29):67-77.
[16] Jobling M, Davies P S. Effects of feeding on metabolic rate, and the specific dynamic action in plaice,PleuronectesplatessaL.[J].J Fish Biol, 1980(16):629-638.
[17] Lucas M C, Priede I G. Utilization of metabolic scope in relation to feeding and activity by individual and grouped zebrafish,Brachydaniorerio(Hamilton-Buchanan)[J]. J Fish Biol, 1992(41):175-190.
[18] 邹桂伟,罗相忠,胡德高,等. 长薄鳅耗氧率与窒息点的研究[J].湖泊科学, 1998,10(1):49-54.
[19] 向伟,范镇明. 叶尔羌高原鳅低温下耗氧率和窒息点的研究[J].水产养殖, 2014,35(4):46-48.
[20] 何光彬,杨 淞,陈柯,等. 似鲇高原鳅幼鱼耗氧率和窒息点的初步究[J].水产养殖,2014,35(10):16-20,47.
[21] 杜宗君,邹建平,李正杰,等. 红尾副鳅耗氧率和窒息点的初步研究[J].四川动物,2013,32(5):695-700,706.
[22] 周翠萍,杨淞,陈柯,等.大鳞副泥鳅耗氧率与窒息点[J].河北渔业, 2014(12):13-16.
[23] 田盛秀,许瑾,杨淞,等. 2种温度下泥鳅耗氧率的昼夜变化规律[J].长江大学学报:自然科学版, 2013,10 (17):33-35.
[24] 顾若波,徐钢春,闻海波.花耗氧率和窒息点的初步研究[J].上海水产大学学报, 2006,15(1):118-122.
[25] 王吉桥,赵兴文.鱼类增养殖学[M]. 大连:大连理工大学出版社, 2000:39-131.
[26] 陈王柯,包杰,倪锡远,等.温度及其突然变化对泥鳅耗氧率、排氨率和窒息点的影响[J].沈阳农业大学学报, 2014,45(6):741-745.
[27] 耿龙武,徐伟,李池陶,等.大鳞鲃耗氧率和窒息点的测定[J].上海水产大学学报,2012,21(3):363-367.
[28] 陈苏维,吉红,朱文东,等.温度对泥鳅消化酶活性影响的研究[J].水产科学, 2009,28(9):528-530.
[29] 刘飞,张轩杰,刘筠.湘云鲫耗氧率和溶氧临界窒息点[J].湖南师范大学 自然科学学报, 2000,23(3):72-75.
[30] 徐连伟,董宏伟,邹作宇,等.匙吻鲟幼鱼耗氧率与窒息点的研究[J].大连水产学院学报,2009,24(4):359-361.
[31] 雷曼红,陈根元,程勇,等. 叶尔羌高原鳅耗氧率和窒息点的初步研究[J].水产养殖,2007,28(6):1-3.
[32] 陈婉情,吴洪喜,吴亮. 5种海水养殖鱼类幼鱼的耗氧率及窒息点[J].海洋学研究,2015,33(2):76-81.
