成年人不同重心高度爬行的运动学特征分析
2017-12-01孟欢欢周喆啸田忠明赵焕彬
孟欢欢,周喆啸,白 锐,田忠明,赵焕彬
成年人不同重心高度爬行的运动学特征分析
孟欢欢1,周喆啸2,白 锐1,田忠明1,赵焕彬1
人类爬行时与其他灵长类和非灵长类四肢动物有共同的四肢协调运动学特征,并已被证明这是由于他们拥有相似的中枢模式发生器(Central Pattern Generators,CPGs)。目的:以8名健康男性成年人不同的重心高度(分别约0.65 m、0.55 m及0.45 m)进行手足爬行为例,在前人研究的基础上对比四肢爬行与直立行走和四足动物移动时的运动学特征。方法:采用8个红外捕捉镜头的三维动态捕捉设备同步采集8名健康成年受试者的四肢和躯干运动学数据。所有受试者皆参与了为期9周的爬行学习与测试(每3周学习及测试1种爬行方法),所有测试皆在受试者熟练掌握爬行动作后进行,3种爬行动作测试完后进行直立行走的数据采集。选取的运动学指标主要包括同侧相位滞后(Ipsilateral Phase Lag,IPL)和关节活动范围。结果与结论:相比其他研究,本研究爬行速度较慢;与直立行走的典型运动学特征相比,受试者表现出较大的运动学协调模式范围(对角线模式与侧向序列模式)以及各主要关节更大的活动范围,且随着爬行时重心高度降低,肩、肘关节活动范围逐渐减小,髋、膝、踝关节活动范围逐渐增大,表明在一定范围内,重心越高支撑面越小,稳定性越差,需要更多的肢体支撑保持稳定,重心越低,越需要肩、肘关节的稳定性及髋、膝、踝关节的灵活性;反之,则需要肩、肘关节的灵活性和髋、膝、踝关节的稳定性。
爬行;重心;运动学特征;关节活动范围
虽然人类是双肢移动的步行方式,但也可以用四肢 行走。人类幼儿在无辅助行走开始前常常爬行,他们大多使用手膝爬行方法,然而,偶尔也会观察到幼儿用手脚爬行[17]。此外,一些研究[19]认为,患有先天性中枢神经系统(Central Nervous System,CNS)疾病导致平衡状态受损的幼儿优先使用手足爬行练习,例如,乌纳塔恩综合征(Uner Tan Syndrome)患者无法直立行走,只能用四肢爬行。
爬行是一种运动模式,成人也可以爬行,但目前对于这种行为的调查和研究较少。大多数的研究只分析了手和膝[1]与手和足[14]的运动学的、短暂的、稳定性的特点,他们发现四肢协调的一个大的变化,这表明人类爬行具有潜在的灵活控制机制。而爬行作为一种反序运动受到越来越多人的关注,但人们对于爬行的机制并不十分清楚。因此,探索人类爬行时的生物力学机制将有助于人们对爬行的理解。根据以往的研究和观察,及相关前期实验的结果,本研究在对大众爬行健身的调查中发现,人类爬行时,不但速度超过临界值后会改变爬行时的运动模式,不同重心高度爬行也表现出不同运动模式的现象。
基于此,本研究目的在于探究成年人在不同重心高度爬行时,步态特征、关节活动范围等运动学特征的变化及与直立行走的差异。研究假设:人类爬行时,改变重心高度,将导致运动模式的改变,重心越高,稳定性越差,需要更多肢体支撑,肘关节和肩关节活动范围越大,髋、膝、踝关节活动范围越小,以及下肢推进力的逐渐减小;但重心高度并不一定是导致上肢缓冲作用大小的主要因素。
1 研究对象与方法
1.1 研究对象
8名健康的男性成年人自愿参加本实验,并了解实验目的,年龄24.0±1.0岁,身高177.0±2.5 cm,体重68.5±3.5 kg。所有受试者皆符合以下标准:1)至少有1年以上的体育锻炼经历;2)无肩、肘、腕、踝关节及腰部等受伤史;3)无神经性疾病、脑部等疾病史。
1.2 爬行动作
高重心(图1):①准备姿势:手足着地,两臂伸直,两手与肩同宽置于肩正下方,两腿伸直(或稍屈)。②③④爬行动作:右腿直腿前进至右脚尖即将触碰到手的位置,脚跟先着地,过度到全脚掌着地,与此同时,屈肘抬起右手至右肩关节处后,向前伸臂至手臂伸直,然后自然落地到右肩的正前方,左脚左手与右侧相同,以此左右侧交替行进。⑤结束姿势:结束姿势与准备姿势相同。

图1 高重心爬行动作步骤示意图Figure 1. High Center of Gravity Crawling Action Steps
中重心(图2):①准备姿势:双手双脚着地,五指自然张开,左臂伸直置于头左前下方,右臂伸直置于右肩下方,抬头眼向前看,左右腿均伸直(或稍弯曲)置于左右后下方,脚跟稍离地,脚尖向前稍向外,膝盖与脚尖方向相同,背部保持平直。②③爬行动作:右脚抬起向起始位置的正前方移动约一脚的距离,脚尖着地,背部挺直,抬头前看,左手左脚不动。右腿向斜后方用力蹬地伸直,将能量传至躯干,躯干向左旋转,右肩外展带动肘外展带动手,在身体右外侧做一鞭打动作,同时重心前移,当手到达最远端时,右手自然落至比左手稍靠前(约20 cm)的位置处,如此反复循环。④结束姿势:结束姿势与准备姿势相同。
低重心(图3):①准备姿势:手足(前脚掌)着地,躯干与地面平行,右侧手膝为近距离,右小腿与地面平行,大腿与小腿成90°,左侧手膝为远距离,抬头眼看前方。②爬行动作:移动时,左脚和右手屈膝屈肘向前先后移动约一脚的距离,保持躯干与地面平行,抬头眼看前方,如此交替前进。③结束姿势:与准备姿势相同。
1.3 数据采集
本实验采集运动学数据的设备是美国motion运动技术分析公司生产的motion三维动态捕捉设备。该设备由8台Raptor-4型号红外数字捕捉镜头和8根网线,相同(直径1.4 cm)的红外反射标记点,1台数据采集盒及1台电脑组成。29个(Helen Hayes模型)红外反射标记点分别放置于左右两侧的:肩峰点、肘点、手腕点、髂前上棘点、大腿点、膝外侧点、膝内侧点、小腿点、踝外侧点、踝内侧点、跟骨点、足尖点;以及头顶点、额前点、枕骨点、右后背点和后腰点。Raptor-4型号数字捕捉镜头(2 352×1 728像素)采用的是美国Micron公司的MI-MV40传感器,本实验采样频率为200 Hz。所有标记点的数据采用一个二阶双通巴特沃斯滤波器进行截止到7 Hz的低通滤波。

图2 中重心爬行动作步骤示意图Figure 2. Medium Center of Gravity Crawling Action Steps

图3 低重心爬行动作步骤示意图Figure 3. Low Center of Gravity Crawling Action Steps
1.4 实验步骤
所有受试者皆参加了本实验的4个步骤:第1~3周的高重心爬行学习及测试;第4~6周的中重心爬行学习及测试;第7~9周的低重心爬行学习及测试;以及低重心爬行测试后的直立行走测试。在受试者熟练掌握爬行动作后再进行测试,受试者每掌握一种爬行动作,进行一次测试,直至完成所有测试动作实验结束。
以上提到的每个步骤皆包括以下实验条件:1)测试前72 h无剧烈运动;2)测试开始前进行约10 min的热身和5 min的动态拉伸;3)为防止因标记点粘贴位置不同造成的数据差异,所有标记点的粘贴都由特定的两个实验人员完成;4)在测试过程中,采集受试者以自然速度爬行时的10个连续的步幅。在完成3种爬行动作的数据采集72 h后,再进行直立行走的数据采集,也包括以上提到实验条件的前3个条件以及采集了10个连续的步幅。
1.5 数据分析
运动学数据使用系统自带的Helen Hayes模型进行处理,该模型由头、躯干、上臂、前臂、大腿、小腿和脚组成。该模型主要用来计算身体右侧上肢的肩、肘和下肢的髋、膝、踝关节(图4A)和肩髋夹角的角度(图4B)。本研究中,使用系统自带的Helen Hayes刚体模型来计算重心(图4C)。上下肢的步态周期时间由右手腕点和脚面点分别接触地面(在Z轴上的最低点)确定。上下肢的摆动时间分别是由右侧手腕点离开地面到右手腕接触地面和脚面点离开地面到脚面点接触地面决定。
采用Patrick等[18]人描述的方法确定上下肢之间的同侧相位滞后(Ipsilateral Phase Lag,IPL)。图5示出了用于描述肢体协调的方法,显示了每个肢体的支撑(实线)和摆动(空格)阶段的时序和持续时间。每个肢体开始支撑和摆动的时间表示为右腿周期的函数(即图5中的T)。例如,在图中,在右腿开始支撑与右臂支撑期开始之间的时间间隔为“t1”。将此延迟表示为右腿周期中持续时间的百分比,称为同侧相位滞后(公式1)。简单地说,用一个步态周期的百分比表示右上肢接触地面的相对时间,步态周期百分比是由右脚连续不断的接触地面决定的:
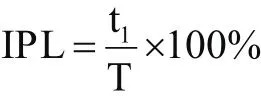
t1是右脚和右手之间着地的间隔时间,T是周期时长(图5)。根据这一方法,同侧步态(同侧上肢/下肢接触地面)是没有取值的,对侧步态模式(对侧上肢/下肢接触类似的情况下)确定的值为50%,中间值(约25%)相当于没有肢体配对。

图4 肩、肘、髋、膝、踝5关节角度及肩髋夹角和重心的定义示意图Figure 4. Schematic Diagram of Joint Angle of Shoulder,Elbow,Hip,Knee,Ankle Joint,Intersection Angle of Shoulder and Hip,and Center of Gravity
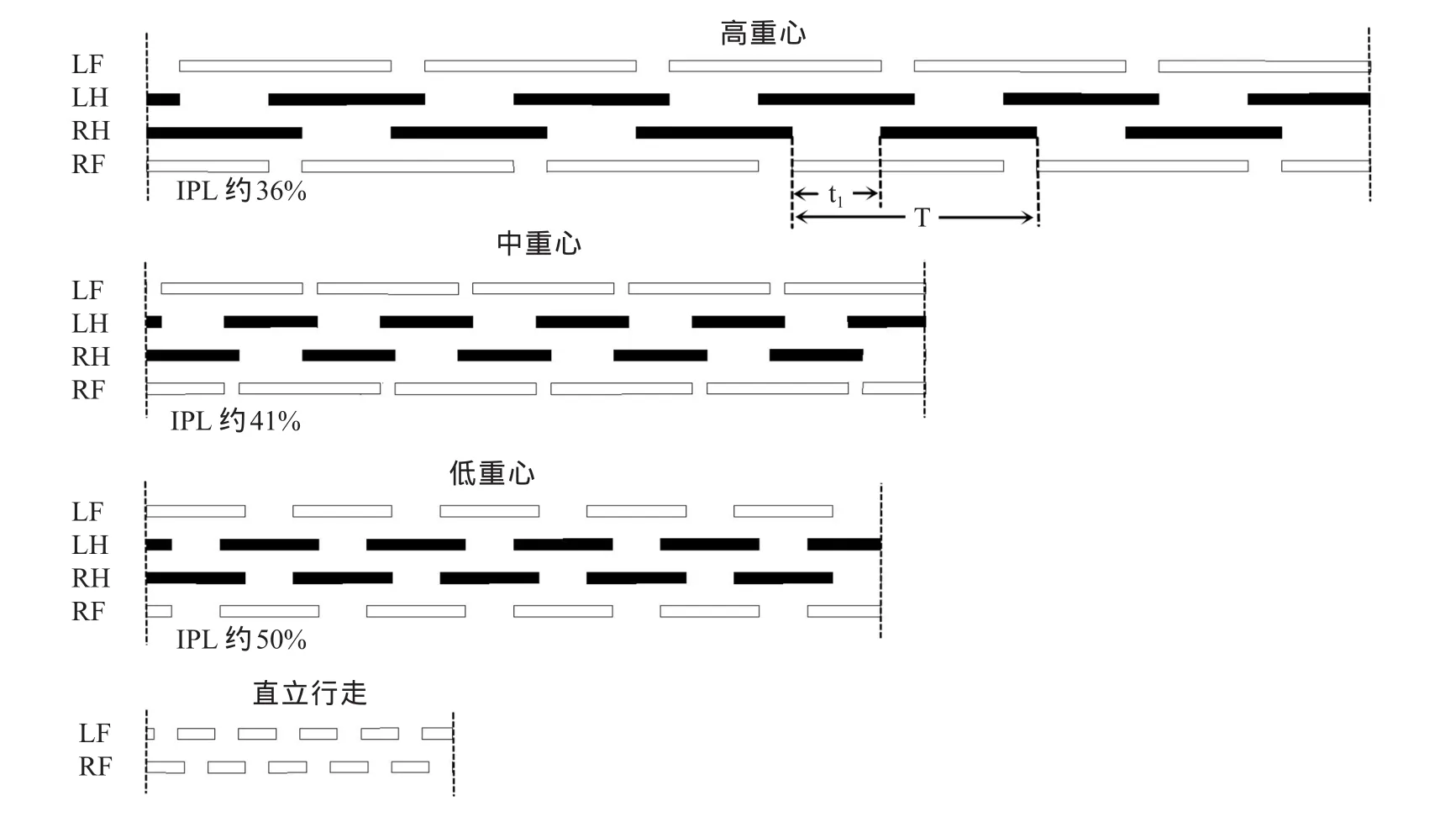
图5 3种爬行动作与直立行走的一般协调模式Figure 5. General Coordinating Modes of Three Kinds of Crawling and Upright Walking
本研究对采集的10个周期数据,每个周期的数据均单独作为一组数据进行计算,描述性统计使用均值±标准差(±SD)表示。爬行重心高度的不同步态和运动模式参数的影响,以及上下肢之间的差异使用单因素方差分析法(ANOVA)评估。对于比较统计,当分布通过正态分布同质性方差的检验时应用参数检验(Tukey检验);否则,使用非参数检验(Dunnett’s检验)。两两之间的比较采用配对样本 t 检验,IPL的统计学分析使用Pearson相关系数。数据均经SPSS 20.0统计软件进行统计学处理,所有统计检验的显著性水平设置为P<0.05。
2 研究结果
2.1 爬行重心高度及速度
本实验所测结果显示,低重心爬行、中重心爬行及高重心爬行在运动过程中的重心高度变化范围分别为0.43~0.48 m、0.49~0.60 m及0.61~0.68 m(图6B)。3种爬行重心变化量之间皆发现了较大差异(P<0.01),但与重心高度的变化没有线性关系(图6A),然而,重心最低的爬行变化量最小。其一般爬行速度分别约为0.28 m/s、0.17 m/s、0.15 m/s,在本研究中的低重心爬行和速度与大多数研究[1,14,18]所描述的爬行方法类似;而随着爬行时重心高度的升高,爬行速度逐渐降低,这可能是由于随着重心的升高,人类关节活动范围降低而导致的。

图6 3种爬行及直立行走重心变化量( ASD) 及1个代表性受试者重心高度变化( B) 对比图Figure 6. Comparison of the Change Quantity of Gravity Center(A,±SD) and Heights of Gravity Center from a Cepresentative Participant( B)of Three Kinds of Crawling and Standing Walking
2.2 基本步态参数
图5展示了不同重心高度下,人类爬行步态参数的基本变化。人类爬行一般是4点、3点或2点交替支撑体重爬行的步态模式。在本研究中步态特征显示,高重心与中重心的四肢分别交替进入摆动期(图5),即,左腿先短暂的摆动后进入支撑期,即刻开始左手的摆动,经过较长的摆动期(远高于左腿的摆动,P<0.05)后再进入支撑期,依次进入右腿摆动期,直至右手进入支撑期开始下一个循环。因此,高重心和中重心爬行属于3点交替支撑体重方法(其中1个肢体处于摆动期时,另外3个肢体皆处于支撑),这种方法与MacLellan等[14]人描述的侧向步态类似。但与之前的研究[14,18]相反的是,本研究中高重心与低重心手的摆动远长于腿的摆动,造成手的支撑远小于腿的支撑(如图5中的高重心与中重心)。在低重心爬行中,对角线的手和脚出现了同时摆动及同时支撑的现象(图5),也就是说,手和脚的支撑与摆动周期比相同。这也与之前的研究(手的摆动远小于腿的摆动)有差异。因此,低重心爬行属于两点交替支撑体重方法。就本研究来看,随着重心高度降低,支撑身体重量的肢体数量减少,速度呈增加的趋势。
本研究通过用步态周期百分比[18]来计算右上肢与地面的接触,以此量化这种协调模式(图5)。以爬行时身体重心高度作为一个函数对上下肢之间的同侧相位滞后(IPL)分析,结果显示,高重心(约36%)与中重心(约41%)爬行之间(P<0.001),中重心与低重心(约50%)爬行之间(P<0.001)皆有显著性差异。总的来说,当前速度下,重心越高受试者越趋向于无肢体配对步态模式(IPL约25%),重心越低越趋向于Patrick等[18]人和MacLellan等[14]人描述的“对角线”模式(IPL约50%)。相关性分析发现,重心高度与IPL值之间具有显著相关性(r=-0.926,P<0.001)。正如预期那样,随着重心高度的降低,周期持续时间有逐渐降低的趋势(高重心与中重心相比P<0.05,中重心与低重心相比P=0.895,图7A),而摆动百分比两两之间皆有显著性差异(P<0.001),但与重心高度无关(图7B、C)。

图7 不同重心高度爬行时周期时长 (A) 和右手 (B) 及右脚 (C) 摆动百分比对比图Figure 7. Compared of Cycle Length(A) and Swing Percentage of Right Hand(B) and Right Foot(C)of Different Hights of Gravity Center
2.3 关节活动范围
图8显示了人类使用3种不同方法爬行时的关节运动学模式。在3种爬行方法之间,肩峰(高重心27.8±3.5 cm,中重心30.9±5.1 cm;P=0.104)和髂前上棘(高重心19.2±4.8 cm,中重心21.4±4.6 cm;P=0.520)的垂直位移量在高重心和中重心之间没有显著差异(图8,A和B的上部),但高重心和中重心皆比低重心(肩峰7.7±1.8 cm,髂前上棘13.1±2.2 cm)有较大的垂直位移量(P<0.001)。值得注意的是,与之前的研究[14]发现不同的是,高重心与中重心肩峰的垂直位移显著高于髂前上棘(P<0.05),而低重心恰恰相反,呈现髂前上棘垂直位移高于肩峰。另外,低重心爬行时的肩峰与髂前上棘的垂直位移皆显著低于(P<0.05)高重心与中重心。

图8 3种方法爬行时的运动学模式图Figure 8. Kinematic Model of Three Methods Crawling
关节运动范围显示,肘、肩、髋关节和踝关节在中重心(活动范围分别为60.2°±16.8°、63.1°±12.7°、89.0°±11.1°、52.3°±13.5°)与低重心(活动范围分别为55.4°±12.4°、65.5°±8.9°、94.7°±10.6°、60.8°±9.2°)之间没有显著差异(P>0.05),而高重心的肘关节(127.1°±3.1°)与肩关节(91.8°±12.0°)显示了较中重心和低重心更大的运动范围,虽然肘关节和肩关节在中重心与低重心之间没有差异(P>0.05),但随着重心的降低,两者皆有逐渐下降的趋势(图8,B的下部左侧)。髋关节和踝关节在3种爬行之间没有显著性差异(P>0.05),但从结果来看,两者有随重心降低而升高的趋势(图8,B的下部右侧)。膝关节(高、中、低分别为24.2°±7.5°、71.3°±9.3°、99.7°±4.1°)在3种方法之间皆表现出了显著性差异(P<0.01),且其运动范围随着重心高度的降低而逐渐增加。肩髋夹角(高、中、低分别为32.2°±2.0°、43.0°±8.2°、28.5°±6.1°)在3种爬行方法之间没有差异(P=0.051),但中重心表现出更大的角度。
2.4 与直立行走相比
直立行走的重心高度变化范围一般在0.95~0.97 m(图6B)。与直立行走相比,3种爬行皆表现了较大的重心垂直位移变化量(图6A)。本研究中的数据显示,人类自然的步行速度约为1.02 m/s,这与Winter[21]的研究结果(1.33 m/s)相差较大,这可能是由于地域差异、种族及身高、实验条件等的影响,这些具体的原因可以在今后的工作中分析,在这里不再阐述。关节活动范围数据显示,与直立行走相比,无论哪种爬行方法,肩峰和髂前上棘的垂直位移皆显著高于(P<0.001)直立行走(肩峰4.6±0.2 cm,髂前上棘4.4±0.3 cm)(图8,B的上部)。另外,直立行走的肩髋夹角(11.4°±1.1°)、肘(26.6°±4.1°)、肩(13.1°±2.9°)、髋(20.7°±1.7°)、踝关节(30.3°±1.4°)活动范围皆显著小于3种爬行(P<0.001),而膝关节(55.6°±0.8°)活动范围高于高重心爬行,但低于中重心和低重心爬行。
本研究还发现,与直立行走不同的是,爬行时上肢的作用是缓冲能量使身体减速,如表1所示,3种爬行方法之间上肢的主要作用是使身体减速,且上肢与水平面的夹角在3种重心高度爬行之间有显著性差异(P<0.05),但并没有一定的规律性。而下肢的主要作用是推进身体前进,并且下肢角度有随重心下降而逐渐减小的趋势(r=0.836,P<0.05),即,随着重心降低,下肢的推进力逐渐增大。
3 讨论
本研究具体探讨了3种爬行方法下人类手足爬行的运动学模式,以及他们之间变异性的原因,发现人类与动物四肢运动的一些相似之处,但也表现出一些人类的特殊特征。
表1 不同重心高度爬行着地时刻上下肢与水平面夹角Table 1 The Angles between the Horizontal Suface and the Upper and Lower Limbs at Different Gravity Centers ( ±SD)

表1 不同重心高度爬行着地时刻上下肢与水平面夹角Table 1 The Angles between the Horizontal Suface and the Upper and Lower Limbs at Different Gravity Centers ( ±SD)
注:上肢小于90°将起到使身体减速缓冲的作用,等于90°,表明上肢与水平面垂直,大于90°,上肢起到推进的作用;下肢小于90°将起到使身体推进的作用,等于90°表明下肢与水平面垂直,大于90°将起到减速的作用。
高重心 中重心 低重心上肢 51°±2° 33°±3° 48°±2°下肢 80°±4° 67°±2° 6°±1°
3.1 爬行速度与重心高度的比较分析
在之前的研究中并没有发现有爬行速度与重心高度之间关系的研究,但已知的有对爬行速度与协调模式之间的研究。在爬行速度较慢的情况下(0.13 m/s),成年人之间没有明显的肢体配对[18]。在本研究中,高重心与中重心的爬行表明,重心越高,爬行速度越慢,即使在试验人员要求受试者以较慢速度爬行的前提下,受试者依然表现出这种现象。由于在爬行过程中,要求受试者手臂有一个较大范围的摆动(也就是说延长手臂的摆动周期,图5),这在一定程度上也减缓了受试者爬行速度。
在本研究中观察到的四肢爬行的速度明显慢于之前的研究[1,14]中观察到的速度,即使是速度最快的低重心爬行(0.28 m/s),依然较慢。一项研究报告显示最大爬行速度为0.64 m/s[2];在另一项研究中,对于慢、中和快速爬行步态,受试者平均速度为0.38 m/s,0.59 m/s和0.82 m/s[1];第3项研究分析了从0.22~1.34 m/s较宽范围的爬行速度[18]。逃生背景研究的爬行速度报告,正常爬行速度为0.71 m/s,最大爬行速度为1.47 m/s[12]。因此,相较于这些研究,本研究中的爬行皆为较慢速度。
另外,考虑到不同爬行速度可能会影响人类的运动模式,因此将本研究结果与之前的研究进行对比,以检查速度在本研究中是否会影响到本实验结果。在对比Maclellan 等[14]人的研究结果时发现,爬行速度在0.5 km/h(0.14 m/s)~3 km/h(0.83 m/s)之间变化时,IPL 值(图9A)没有显著性差异(P=0.87),髋关节、膝关节、踝关节及腕关节活动范围皆没有显著性差异(图9B),而肩关节和肘关节活动范围有随速度提升逐渐增加的趋势(P<0.05)。因此,在本研究中排除了运动速度对运动模式的影响,只考虑重心高度对运动模式的影响。
3.2 协调模式与其他哺乳动物具有共同特征
本研究中所有受试者在使用高重心和中重心爬行时几乎完全使用侧向序列步态,这与以前的婴儿[3]、儿童[9]和成人[4]的爬行研究一致。大多数四足动物(如果不是特定的原因)优先选择使用侧向序列步态模式[9]。最值得注意的例外是非人类灵长类动物,通常使用对角序列步态[9],这与本研究中低重心爬行使用的步态模式一致。非人类灵长类动物和其他四足动物之间的这种差异的原因仍然有争议,并且超出了本研究的范围。
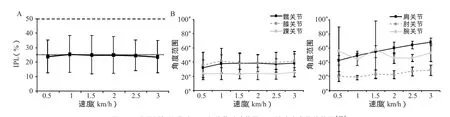
图9 平均同侧相位滞后 (A) 和关节活动范围 (B) 随速度变化趋势图 [22]Figure 9. Trend of Ipsilateral Phase Lag for the Group Average and Ranges of Motion with Speed
高重心和中重心爬行所使用的侧向序列步态模式与人类直立行走步态模式类似,但爬行更多地表现为同侧肢体摆动期之间没有时间间隔,更长的共同支撑时间以及四肢之间没有共同的支撑阶段(图5)。而低重心爬行的对角线序列步态模式更加类似于非人类灵长类动物,更多地表现出对角线肢体的同时摆动与同时支撑,四肢之间有一定时间的共同支撑阶段,这与自然直立行走的双肢移动类似。当然,在人类跑步时,双肢移动会出现腾空阶段(没有共同支撑时间),这是特殊的步态模式,而爬行在快速的条件下(类似于直立小跑)并没有出现这种情况[18]。
非灵长类哺乳动物[7]和非人类灵长类动物[20]常常在不同的运动速度下使用不同的肢体协调模式。由此来看,对于人类来说,特别是成年人,也具有这种特点。在比较慢的爬行速度条件下(高重心和中重心分别为0.15 m/s,0.17 m/s),其更趋向于无肢体配对(图5,IPL分别约为36%和41%)。这与以下事实一致:在较低速度下的运动步态需要提供更高稳定性,例如无肢体配对协调[10],其中个体可能在3个肢体支撑中花费步态周期的大部分时间。相反,低重心爬行速度更快(约为0.28 m/s),表现出对角线序列步态(图5,IPL约为50%),虽然其有四肢共同支撑的节段,但占整个周期很少(约17%),在爬行过程中主要起过渡作用(与直立行走的过渡类似)。
3.3 关节活动范围的对比分析
人类双肢行走是从四肢移动进化而来,但人类依然保留着四肢移动的能力,在人类幼儿无辅助直立行走前通常爬行[19]。之前的研究表明,人类能够使用较多的步态模式完成四肢移动[18,13],在本研究中也发现这种情况(高重心、中重心和低重心不同爬行模式)。相比直立行走,大多数人类爬行表现出更大的关节运动范围(图8B),这种情况通常对人类各关节肌群的伸展性提出了更高的要求,尤其是高重心爬行对人体股后肌群和肩部肌群的柔韧性影响最大。爬行速度与关节活动范围的相关性分析发现,爬行速度与膝关节活动范围呈正相关(r=0.694,P<0.001),即膝关节活动范围越大爬行速度越快;爬行速度与肘关节呈负相关(r=-0.531,P<0.001),即肘关节活动范围越大,爬行速度越慢。而爬行重心高度与关节活动范围之间关系的研究发现,重心高度越高肘关节(r=0.817,P<0.01)和肩关节(r=0.618,P<0.01)活动范围越大;相反,髋关节(r=-0.668,P<0.01)、膝关节(r=-0.940,P<0.01)及踝关节(r=-0.260,P<0.05)随着重心高度的增高,活动范围逐渐降低;肩髋夹角与重心高度之间没有相关性(P=0.231)。
此外,人类爬行可能还受生物力学的影响。在本研究中采用的是手足爬行,目的是与四足动物比较。在人类幼儿中常常使用手膝爬行,但偶尔也会见到手脚爬行[17]。如果当前的方案使用手膝爬行,将减少下肢的功能长度。而这个减少的功能长度可能影响肢体协调模式,但在目前的研究中这不是一个特定的问题,可以在将来的研究中解决。在本研究中,爬行时上肢比下肢短,这将导致上肢以更高的频率爬行。然而,与预测相反的是,上下肢始终以1:1的频率关系前进,与一些动物相似(如,狐猴也有较长的下肢,但依然可以1:1的频率关系移动)。此外,肩胛骨回缩倾向于按照四肢中前肢的长度延伸,进而补偿这个较低的功能范围[6]。在人类爬行期间可能存在类似的机制,这将使肢体长度上的差异最小化。因此,虽然生物力学因素可能影响肢体运动,但是,在肢体之间保持整数比的事实表明这种模式的神经起源[13]。
3.4 人类爬行的神经机制
当前的研究显示的上下肢摆动与支撑比和那些在跑步机上爬行期间的研究[14,18]显示完全不同的结果可能与通过大脑皮质发射的信号覆盖了上肢中枢模式发生器的控制有关。在人类中,与猫相比,上肢更多的是非节奏性任务。此外,Nakajima等[15]人比较不同种类的动物显示,手的强大功能和大脑皮质运动神经元直接投射到手的肌肉之间的正相关,以及手的强大功能和脊髓固有信号传递之间的负相关。对于手的精细运动,这种模式被认为是与大脑皮层系统取代了脊髓系统有关。因此,人类上肢这种强烈的大脑皮层影响可能有助于在增加上肢关节角度的同时增加摆动期。
或者说,对于上肢在人类中以不同摆动和支撑比爬行的另一解释,可能是由于人类脊髓固有神经元的上升和下降的比例不同。在人类中,CPGs回路的主要轨迹是腰骶段扩大,并且,颈段和腰骶段CPGs的耦合可以依赖于脊髓神经连接的强度。事实上,人类中的颈段和腰骶段扩大似乎是通过长的脊髓神经元跨过几个脊髓阶段投射到不同的α运动神经元池进行的连接[8]。此外,反射研究表明,在有节律活动期间,刺激上肢神经可以影响下肢肌肉激活,反之亦然[16,22]。有趣的是,当受试者站立或坐着时,Dietz等[5]人在胫骨神经刺激期间观察到小的或缺乏上肢反射反应,提示此类反应的任务依赖性。相比之下,对孤立的新生大鼠脊髓的体外研究表明,腰段CPGs对颈段CPGs产生了更大的影响[11]。然而,这些体外研究和本研究中提出的建议之间的结论差异可能是由于研究的准备、实施或任务依赖。根据本研究的发现,推测这些上升的脊髓固有连接的强度可能在人类中较弱,并且最终允许大脑皮层信号对上下肢的摆动支撑比产生影响。
3.5 研究局限性及展望
1)本研究虽然进行了爬行学习,但测试的受试者皆属于爬行初学者,对于长期坚持爬行的人,可能会表现出一定的差异,因此,本研究应该被理解为相对新手爬行的结果。2)本研究的生物力学研究还缺少动力学设备的测试,这对理解爬行时重心变化,肢体和关节承受力的情况产生影响,以及由于场地设备限制,缺少受试者爬行速度的控制,也造成不能较为严谨、全面描述运动模式的改变。3)本研究所测试的对象为成年健康男性,那么,对于婴幼儿、女性爬行或肢体功能障碍患者或其他人群的爬行,本研究的结果可能并不完全适用。
因此,考虑到以上因素,在未来的爬行研究中,应对长期坚持爬行的人和不同健康程度、性别、年龄人群的爬行进行研究,并加入动力学测试设备,及加入跑步机等设备控制爬行速度,更加科学、清晰、全面的理解人类爬行时的生物力学变化。
4 结论
人类爬行时,重心越高步态模式越趋向于无肢体配对模式,即3点交替支撑模式;反之,越趋向于对角线步态模式,即2点交替支撑步态模式,表明在一定范围内,重心越高支撑面越小,稳定性差,需要更多的肢体支撑保持稳定。
相比直立行走,大多数人类爬行表现出更大的关节运动范围,且随着爬行时重心高度降低,肩、肘关节活动范围逐渐减小,髋、膝、踝关节活动范围逐渐增大,表明重心越低,越需要肩、肘关节的稳定性及髋、膝、踝关节的灵活性;反之,则需要肩、肘关节的灵活性和髋、膝、踝关节的稳定性。
人类爬行时,上肢主要起缓冲和减速身体的作用,下肢起推进身体前进的作用,且随着重心高度降低,下肢着地角逐渐减小,表明重心越低,下肢水平向前的推进力越大。
本研究结果可能会对临床应用和仿生机器人的构建起到作用,当需要增加难度时,增加重心高度;反之,则降低重心高度。
[1] BABIČ J,KARČNIK T,BAJD T. Stability analysis of four-point walking[J]. Gait Posture,2001,14(1):56-60.
[2] BAJD T,ZEFRAN M,KRALJ A. Timing and kinematics of quadrupedal walking pattern[J]. Intell Robot Syst,IEEE/RSJ Int Confer,1995,3(3):303-307.
[3] BURNSIDE L H. Coordination in the locomotion of infants[J].Genet psychol Monogr,1927,2(5):360-360.
[4] DEMES B,LARSON S G,STEMN J T,et al. The kinetics of Primate quadrupedalism:“hindlimb drive” reconsidered[J]. J Hum Evol,1994,26(5/6):353-374.
[5] DIETZ V,FOUAD K,BASTIAANSE C M. Neuronal coordination of arm and leg movements during human locomotion[J].Eur J Neurosci,2001,14(11):1906-1914.
[6] FISCHER M S,BLICKHAN R. The tri-segmented limbs of therian mammals:Kinematics,dynamics,and self-stabilization—A review[J].J Exp Zool A Comp Exp Biol,2006,305(11):935-952.
[7] GRILLNER S. Control of Locomotion in Bipeds,Tetrapods,and Fish Handbook of Physiology,the Nervous System,Motor Control[M]. Bethesda:Am Physiol Soc,1981:1179-1236.
[8] HARIDAS C,ZEHR P E. Coordinated interlimb compensatory responses to electrical stimulation of cutaneous nerves in the hand and foot during walking[J]. J Neurophysiol,2003,90(5):2850-2861.
[9] HILDEBRAND M. Symmetrical gaits of primates[J]. Am J Phys Anthropol,1967,26(2):119-130.
[10] HILDEBRAND M. The quadrupedal gaits of vertebrates[J].Biosci,1989,39(11):766-775.
[11] JUVIN L,LE GAL J P,SIMMERS J,et al. Cervicolumbar coordination in mammalian quadrupedal locomotion:Role of spinal thoracic circuitry and limb sensory inputs[J]. J Neurosci,2012,32(3):953-965.
[12] KADY R A,DAVIS J. The effect of occupant characteristics on crawling speed in evacuation[J]. Fire Saf J,2009,44(4):451-457.
[13] MACLELLAN M J,QADERDAN K,KOEHESTANIE P,et al.Arm movements during split-belt walking reveal predominant patterns of interlimb coupling[J]. Hum Mov Sci,2012,32(1):79-90.
[14] MACLELLAN M J,IVANENKO Y P,CAPPELLINI G,et al.Features of hand-foot crawling behavior in human adults[J]. J Neurophysiol,2012,107(1):114-125.
[15] NAKAJIMA K,MAIER M A,KIRKWOOD P A,et al. Striking differences in transmission of corticospinal excitation to upper limb motoneurons in two primate species[J]. J Neurophysiol,2000,84(2):698–709.
[16] NATHAN P W,SMITH M,DEACON P. Vestibulospinal,reticulospinal and descending propriospinal nerve fi bres in man[J].Brain,1996,119(6):1809-1833.
[17] PATRICK S K,NOAH J A,YANG J F. Developmental constraints of quadrupedal coordination across crawling styles in human infants[J]. J Neurophysiol,2012,107(11):3050-3061.
[18] PATRICK S K,NOAH J A,YANG J F. Interlimb coordination in human crawling reveals similarities in development and neural control with quadrupeds[J]. J Neurophysiol,2009,101(2):603-613.
[19] TAN U. Uner tan syndrome:history,clinical evaluations,genetics,and the dynamics of human quadrupedalism[J]. Open Neurol J,2010,4(1):78-89.
[20] VILENSKY J A,LIBII J N,MOORE A M. Trot-gallop gait transitions in quadrupeds[J]. Physiol Behav,1991,50(4):835-842.
[21] WINTER D A. The Biomechanics and Motor Control of Human Gait:Normal,Elderly,and Pathological[M]. 2nd ed.Ontario:University of Waterloo Press,1991:369-384.
[22] ZEHR E P,FRIGON A,HOOGENBOOM N,et al. Facilitation of soleus H-re fl ex amplitude evoked by cutaneous nerve stimulation at the wrist is not suppressed by rhythmic arm movement[J].Exp Brain Res,2004,159(3):382-388.
Kinematical Analysis of Adult Crawling with Different Heights of Gravity Center
MENG Huan-huan1,ZHOU Zhe-xiao2,BAI Rui1,TIAN Zhong-ming1,ZHAO Huan-bin1
Human,other primates and non-primate quadrupeds share common kinematic characteristics of limb coordination,which has been proven that they have a similar central pattern generators (CPGs).Objective:Based on previous studies,we found and compared the kinematic characteristics of the movement among crawling human,erectly walking human and moving quadrupeds by studying handand-foot crawling of 8 healthy male adults with different heights of gravity center(~0.65m,~0.55m and ~0.45m,respectively). Methods:In this study,kinematic data of limbs and torso were synchronously collected from 8 healthy adult subjects through a three-dimensional dynamic capture devices equipped with 8 IR capture lenses. All subjects were involved in a 9-weeks reptile study and testing(every 3 weeks to learn and test a crawling method). All tests were performed after the subjects were familiar with the crawl,and the data were collected for upright walking after three kinds of crawling tests. The selected kinematic parameters mainly include ipsilateral phase lag and range of joint motion. Results and Conclusion:Compared to other studies,this research crawling slower;the subjects showed a larger range of kinematic coordination modes(diagonal pattern and lateral sequence pattern) and a greater range of motion of the major joints,compared with the typical kinematic characteristics of upright walking,And the shoulder joint and elbow joint activities gradually decreases,the hip,knee and ankle motion range increased gradually with the crawling center of gravity height reduced. It showed that in a certain range, the higher the center of gravity, the smaller the bearing surface, poor stability,which need more limb support stability. The body center of gravity is lower,the more you have to the stability of the shoulder, elbow,and the fl exibility of the hip,knee and ankle;on the contrary,body will need to shoulder, elbow joint fl exibility and stability of the hip,knee,ankle.
crawling;center of gravity;kinematic characteristics;range of motion
G804.6
A
1000-677X(2017)11-0030-10
10. 16469/j. css. 201711004
2016-12-21;
2017-10-23
国防科技项目(17-163-12-ZT-002-124-01)。
孟欢欢,男,在读硕士研究生,主要研究方向为运动生物力学和大众健康,E-mail:mhyyse@126.com;周喆啸,男,讲师,博士,主要研究方向为身体功能性训练和儿童动作发展,E-mail:zhouzhexiao@126.com;赵焕彬,男,教授,博士,博士研究生导师,主要研究方向为运动生物力学和运动训练学,Tel:(0311)8078779,E-mail:huanbinzhao@126.com。
1. 河北师范大学 体育学院,河北 石家庄 050024;2.宁波大学 体育学院,浙江 宁波 315211
1. Hebei Normal University,Shijiazhuang 050024,China;2.Ningbo University,Ningbo 315211,China.
