植物对土壤硒的吸收转化研究进展
2017-11-23陈锦平刘永贤曾成城潘丽萍鹿士杨黄雁飞梁潘霞江泽普黄太庆
陈锦平, 刘永贤*, 曾成城, 潘丽萍, 鹿士杨, 兰 秀, 黄雁飞, 梁潘霞, 江泽普, 邢 颖, 廖 青, 黄太庆
1.广西农业科学院农业资源与环境研究所, 南宁 530007; 2.广西大学农学院, 南宁 530005
植物对土壤硒的吸收转化研究进展
陈锦平1, 刘永贤1*, 曾成城1, 潘丽萍1, 鹿士杨1, 兰 秀2, 黄雁飞1, 梁潘霞1, 江泽普1, 邢 颖1, 廖 青1, 黄太庆1
1.广西农业科学院农业资源与环境研究所, 南宁 530007; 2.广西大学农学院, 南宁 530005
硒是人类、动物和某些微生物必需的微量元素。植物因其具有吸收和转化土壤硒的能力而成为人类饮食补硒的主要来源。提高植物的富硒水平对于人类健康具有重要意义。为更好地了解影响植物富硒效果的因素,综述了土壤中硒的赋存形态,重点阐述了植物对土壤不同形态硒的吸收、迁移和代谢过程,并由此对硒生物强化未来的研究方向进行了展望。
硒;植物;吸收;转化
硒是维持人体健康的一种必需微量元素,已有研究指出:硒摄入量小于30 μg/d则为摄入量不足,而摄入量大于900 μg/d就可能对人体造成毒害[1]。在某些地区硒中毒是一个不可忽视的问题[2],但硒摄入不足更为普遍,世界范围内约有10亿人受此影响[3]。硒与人类健康息息相关已成为人们的共识,缺硒不仅会导致人体免疫功能下降,还会引发疾病[1]。在低硒饮食摄入群体中,硒剂的额外补充是降低缺硒所引起的健康问题的一种有效途径。对于大部分缺硒人群来说,通过饮食摄入是最主要的补硒方法。不同形态的硒具有不同的营养价值,饮食补硒水平的高低与食用作物的硒浓度和硒形态密切相关[1]。由此看来,植物从根部对外源硒的吸收转化和向可食部分的转移决定了作物的富硒效果[4]。了解植物对土壤硒的吸收转化过程对于提高植物的富硒能力,改善硒的存在形态以及调节硒在可食部位的积聚均具有重要的指导意义。
1 土壤中硒的形态
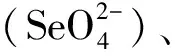
硒化氢(H2Se)是通过土壤微生物活动而生成的挥发性化合物[6]。有机态硒是一种重要的生物有效硒,一部分来源于动植物体含硒组织的分解,一部分来源于土壤微生物的代谢转化[6]。亚硒酸盐和硒酸盐是另外两种重要的生物有效硒,在土壤中大量存在,具有良好的水溶性,因此也容易发生流失[10]。亚硒酸盐易与铁氧化物和氢氧化物等结合生成水溶性极低的氧化物[10]。硒酸盐则不然,研究发现其在施入土壤培养2周后仍能保持高比例的可溶态[11]。因此在土壤中,硒酸盐的生物有效性要高于亚硒酸盐[12]。整体来看,土壤硒形态是土壤酸碱性和氧化还原条件综合作用的结果,在碱性和氧化条件下(Eh+pH>15),硒酸盐是主要形态;在酸性土壤及还原条件下(7.5< Eh+pH<15),亚硒酸盐是主要存在形态;当土壤处于(pH+Eh)<7.5的高度还原条件时,HSe-是pH>3.8的土壤中水溶性硒的主要形态,当pH低于3.8时,H2Se则为土壤硒的主要形态[13]。
2 植物对土壤硒的吸收和转移
2.1植物对土壤硒的吸收
植物只要种植在含硒土壤中都会或多或少地吸收土壤中的硒,根据植物对硒的吸收能力大小可以将植物分为3种类型:非积聚型(<100 mg/kg干重)、积聚型(100~1 000 mg/kg干重)和超积聚型(>1 000 mg/kg干重)[14]。植物对硒的吸收主要取决于环境条件、土壤和植物等因素,其中最重要的是硒在土壤中的存在形态和浓度[5]。硒酸盐、亚硒酸盐和有机硒是植物能够利用的3种主要水溶性硒[4]。
2.1.1植物根系对有机硒的吸收 硒代蛋氨酸(SeMet)和硒代半胱氨酸(SeCys)是常见的两种有机态硒。一般这两种形态的有机硒是由植物对无机硒进行转化而来,但也有研究表明植物可以直接吸收这些有机硒[4]。Abrams等[15]研究发现植物具有利用细胞膜上的氨基酸转运蛋白吸收有机态硒(如硒代蛋氨酸SeMet)的能力。并有学者猜测植物很有可能是通过脯氨酸通透酶途径对硒代半胱氨酸(SeCys)和硒代蛋氨酸(SeMet)进行吸收,因为在跨膜运输时他们之间存在着强烈的竞争关系[4],但有待进一步证实。不仅如此,关于小麦和油菜的富硒研究发现,它们对有机硒(硒代蛋氨酸和硒代胱氨酸)的吸收速率极高,小麦根部对5.0 μmol/L硒代蛋氨酸和5.0 μmol/L硒代胱氨酸处理的吸收速率分别是1.11 μg/g·min和0.191 μg/g·min,油菜则依次分别为6.53 μg/g·min和0.354 μg/g·min,是硒酸盐或亚硒酸盐吸收速率的20多倍[16]。与无机硒相比,有机硒具有无毒、安全、吸收效果好等诸多优点,李玉梅等[17]用硒代蛋氨酸对水稻进行硒强化发现,有机硒施用量在30~60 g/hm2时可达到富硒水稻生产标准,可见有机硒也可做为作物生物强化的一种重要外源硒。
2.1.2植物根系对硒酸盐的吸收 硒和硫是同主族元素,具有相似的化学结构和性质,植物对硒酸盐和硫酸盐的吸收具有极高的相关性[18]。除了硒超积聚植物外(如沙漠王羽),大部分植物并不能有效区分二者[19,20]。在分辨硒积聚能力的高低上,Se/S值也被认为是极为有效的指标[21]。研究表明,在不同的硒酸盐与硫酸盐供给条件下,硒酸盐与硫酸盐均是通过硫转运通道进入植物体内[22]。这些硫转运子在硒超积聚植物中大量存在,在根部、维管束系统和叶片等不同部位发挥相应的吸收或运输功能,大致可以分为4个类别[4]。其中第一个类别的转运子(SULTR1;1和SULTR1;2)与硫具有高亲和力,在根部对硒酸盐的吸收过程中发挥着重要的作用;第二种类别的转运子(SULTR2;1和SULTR2;2)与硫的亲和力较低,分布在维管束系统和根、叶等器官,主要作用于硒的迁移转运[4,23,24]。第三类转运子SULTR3;1则主要存在于叶绿体中,负责硒酸盐跨膜进入叶绿体的运输,这种转运子的缺失会明显降低叶绿体中硫酸盐的含量[25]。而SULTR4;1和SULTR4;2转运子则有助于硒在根-茎之间的转移[26]。有学者通过对植物进行缺硫处理研究也发现,在硫饥饿处理情况下,植物能够上调根部硫转运基因的表达,从而相应地提高植物对硒酸盐的吸收,处理组对硒酸盐的吸收是对照组的4.81倍[24,27]。此外,硒酸盐进入叶绿体后的生理代谢也仍然是通过硫的同化途径来实现[28]。硒代谢与硫代谢的紧密关系可见一斑。
2.2植物对不同形态硒的转移
植物根系从土壤中吸收硒后,在植物体内的迁移转化直接影响植物的生理代谢和硒的富集积累。硒从植物根部向地上部分转移的快慢和比例也取决于硒的形态,不同形态硒的转运速率大体是硒酸盐>有机硒>亚硒酸盐,三者在植物体内的分配比例大致是1.1~17.2、0.6~1.0和<0.5[37]。
植物吸收硒酸盐后,能保持原有形态并随即向地上部转移,根部和枝条中很快就能检测到以硒酸盐为主的硒,同时也有少量的有机硒[27,35]。研究表明,加入硒酸盐处理的菜豆植株中,根系吸收的硒酸盐总量的50%均转移到了植物地上部分[32]。相似的,在花椰菜、印度芥菜、甜菜和水稻等作物的研究中,以硒酸盐为外源硒的处理组其地上部与地下部硒含量比值范围在1.4~17.2,而以亚硒酸盐处理的仅是0.2~0.5,以有机硒(SeMet)处理的是0.6~1.0[31]。硒酸盐虽然具有很高的迁移能力,但其转化为有机硒的速率很低,处理3 d后硒酸盐占总硒的比例仍高达80.9%,只有少数硒酸盐转化为其他形态[27]。
相反,植物对亚硒酸盐的转移能力较低,主要是因为其被根部吸收后极易转化成其他形态,比如有机态硒[35]。Huang等[27]对小麦进行3 d的亚硒酸盐处理后其有机硒含量即接近于总硒的90%。而这些生成的物质大部分可以直接积累在根部,因此不易转移到地上部分,这个结论也得到了Jiang等[38]的印证。
总体来看,植物对硒酸盐向地上转移的能力远大于亚硒酸盐[39]。但也有例外,比如以亚硒酸盐处理的胡萝卜却表现为地上部分硒含量最高,根部最低[40]。有研究表明硒酸盐的转移也会受到硫酸盐施加的影响[30]。因此作物对硒的迁移应该是作物硒吸收转运能力、作物类型、环境因子及土壤供硒能力与类别等综合因素作用的结果。
3 硒在植物体内的形态和转化
3.1硒在植物体内的存在形态
植物在对硒的代谢过程中可能会生成一些挥发态的硒,如二甲基硒醚(DMSe)和二甲基二硒醚(DMDSe),挥发态硒在植物中含量很低,仅占全硒质量比的0.3%~7.0%[41],在硒超积聚植物中较为常见,将植株内的硒转化为挥发态而进行外排是植物降低硒与蛋白质结合的一种有效机制[42]。无机硒的含量也不高,约占全硒质量比的10%~15% ,主要以硒酸盐形式存在[42],这跟硒酸盐的代谢路径受更多的酶限制有关。有机态硒是植物体内含量最高的硒形态,占总硒量的80%以上,其中硒蛋白是含量最高的有机态硒[43]。赵萍等[44]对小麦不同形态的硒进行研究发现,总硒的84.4%是有机硒,有机硒中的54.5%是硒蛋白,其次为硒多糖(15.8%),并存在少量的硒核酸和其他形态的硒。在饮食补硒过程中,人们从食物中获取的硒主要是具有高度生物利用性的硒代氨基酸,植物正因为具有能够将无机硒转化为有机硒的能力,因此植物对硒的吸收代谢功能对人类营养和健康具有重要意义[2]。值得一提的是,Duncan等[45]研究发现,对小麦进行硒生物强化的同时,结合氮肥和硫肥的施加能有效改变小麦不同形态硒的含量,在试验的4个小麦品种中有3个品种的硒代蛋氨酸(SeMet)含量都受到了施肥处理的影响,这为人为调控不同形态硒的含量提供了一种有效方法。
3.2硒在植物体内的转化
根据土壤硒形态的不同,硒在植物体内的代谢转化也存在一定的差异。植物吸收有机硒后,能保持原有的有机硒形态直接参与代谢,非特异性地参与蛋白质的构成。植物通过根部吸收硒酸盐后的同化代谢过程主要发生在地上部分,其中叶片叶绿体是硒发生同化作用的主要场所[20],主要代谢途径见图1。与亚硒酸盐相比,硒酸盐需要首先被还原为亚硒酸盐才能进行下一步转化。
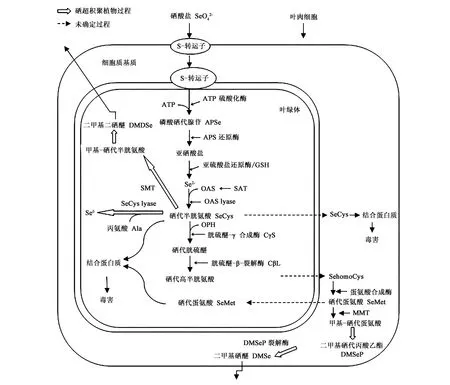
图1 硒酸盐在植物叶肉细胞中的同化代谢 [4]Fig.1 Assimilation and metabolism of selenate in plant mesophyll cells[4].注:SMT:硒代半胱氨酸甲基转移酶;SeCys lyase:硒代半胱氨酸裂解酶;GSH: 谷胱甘肽;SAT:乙酰转移酶;MMT:甲基转移酶;OAS:乙酰丝氨酸;OAS lyase:乙酰丝氨酸裂解酶;OPH:O-磷酸高丝氨酸。
如图1所示,根系通过硫酸盐转运子(SULTR1;1和SULTR1;2)吸收土壤硒酸盐后,经木质部运输,通过SULTR3;1转运子进入叶片的叶绿体中,在ATP参与下硒酸盐首先被 ATP硫酸化酶激活形成 5′-磷酸硒代腺苷(APSe)[46],随后在5′-磷酸腺苷还原酶(APR)的作用下形成亚硒酸盐。由于细胞质基质也有APS和APR同工酶的存在,因此硒酸盐的还原过程很可能也存在于细胞质基质中[18]。在这些过程中硒酸盐还原为亚硒酸盐是硒酸盐在植物体内进行生理代谢的限速步骤,参与反应的ATP硫酸化酶和APR是这些限速步骤的关键酶[28]。硒酸盐转化成亚硒酸盐后则能进入下一步的还原反应,还原反应过程与硫酸盐类似,由亚硫酸盐还原酶介导,仅在叶绿体中进行[33,47]。此外,还原型谷胱甘肽(GSH)通过非酶促反应在还原亚硒酸盐的过程中同样具有重要作用[33,48]。还原反应所产生的硒化物在乙酰丝氨酸(OAS)巯基裂解酶(半胱氨酸合成酶)的作用下与乙酰丝氨酸 (OAS)反应生成硒代半胱氨酸(SeCys)[49],参与反应的半胱氨酸合成酶同时存在于细胞质和线粒体中[50]。其中的乙酰丝氨酸(OAS)由乙酰转移酶(SAT)合成,并扮演着上调硫同化速率的信号分子角色[47]。
生成的硒代半胱氨酸(SeCys)可以直接(或跨膜进入细胞质)非特异性地参与蛋白质的构成,也可以继续转化为硒代蛋氨酸(SeMet),部分植物能将其转化为二甲基硒醚(DMSe)或二甲基二硒醚(DMDSe)等挥发性硒化物。转化为挥发性化合物主要发生在硒超积聚植物中,生成二甲基二硒醚(DMDSe)需要通过甲基转移酶(SMT)介导,并首先转化为甲基-硒代半胱氨酸MeSeCys,其反应所需的SMT在超积聚植物对硒的富集和耐受中扮演着重要角色[50]。硒代半胱氨酸(SeCys)若要转化为硒代蛋氨酸(SeMet)则需要通过3种酶依次参与发生反应才能生成,首先需在胱硫醚-γ-合成酶(CγS)的作用下与O-磷酸高丝氨酸(OPH)结合生成硒代胱硫醚,接着在胱硫醚-β-裂解酶(CβL)的作用下转化为硒代高半胱氨酸(sehomoCys),叶绿体是这两步反应所在的场所,细胞质基质是接下来反应的主要场所[28]。硒代高半胱氨酸(sehomoCys)需要通过跨膜运输进入细胞质基质,才能在蛋氨酸合成酶作用下生成硒代蛋氨酸(SeMet)。同样地,硒代蛋氨酸(SeMet)一方面能够直接(或跨膜进入质体)参与蛋白质合成,另一方面在某些硒超积聚植物中也可以在甲基转移酶及 DMSeP 裂解酶作用下继续发生转化生成挥发性的二甲基硒化物(DMSe)[18],从而降低硒与蛋白质的结合。
从以上转化途径来看,硒超积聚植物为了减少硒非特异性地参与蛋白质构成,具有自身独特的代谢途径。研究表明这些特异性代谢途径能有效区分硫元素与硒元素,这对保证正常的蛋白质功能具有重要意义[23,47]。转化为挥发性硒化物是硒超积聚植物降低硒毒害的一种重要机制,其中硒化物的挥发速率与物种差异有关,并受到土壤硒、硫形态和浓度以及季节等多种因素的影响[50]。总的来说,不管是硒超积聚植物还是非积聚植物,植物的硒形态与植物物种类型和外源硒形态都紧密相关。
4 展望
综上所述,亚硒酸盐在植株体内的迁移能力较低,但更容易转化为有机态硒,在还原性强的土壤中种植以根部为可食部位的作物,能达到更有效的富硒效果。硒酸盐往地上转移的比例和速率较高,但转化为有机硒时会受到相关酶的限制,增强这些酶的活性是提高地上可食部分有机硒的关键。植物对有机硒的吸收效率极高,但相关研究甚少,对有机硒的吸收途径有待进一步确认。
植物硒代谢是生成有机硒的必经过程,了解这些过程对于作物有针对性地进行硒生物强化具有重要的指导意义。从人类营养学的角度来看,植物可食部位有机硒的富集对人体的健康有着重要影响。结合影响植物硒吸收和转化的因素,为了使作物达到更优质的富硒效果,物种筛选、硒代谢调控、土壤性质调整以及施肥管理等方向值得人们进一步探究。
[1] Fairweather-Tait S J, Bao Y, Broadley M R,etal.. Selenium in human health and disease[J]. Antioxid. Redox. Sign., 2011, 14(7): 1337-1383.
[2] Zhu Y G, Pilonsmits E A M, Zhao F J,etal.. Selenium in higher plants: Understanding mechanisms for biofortification and phytoremediation.[J]. Trends Plant Sci., 2009, 14(8):436-442.
[3] Combs G F. Selenium in global food systems [J]. Brit. J. Nutr., 2001, 85(5): 517-547.
[4] Guignardi Z, Schiavon M. Biochemistry of plant selenium uptake and metabolism[A]. In: Elizabeth A H P, Lenny H E,etal.. Selenium in Plants[M]. Springer International Publishing, 2017, 21-34.
[5] 陈松灿, 孙国新, 陈 正, 等. 植物硒生理及与重金属交互的研究进展[J]. 植物生理学报, 2014, 50(5): 612-624.
[6] Fernández-Martínez A, Charlet L. Selenium environmental cycling and bioavailability: A structural chemist point of view[J]. Rev. Environ. Sci. Biol., 2009, 8(1):81-110.
[7] Sharma V K, McDonald T J, Sohn M,etal.. Biogeochemistry of selenium. A review [J]. Environ. Chem. Lett., 2015, 13(1): 49-58.
[8] Jones G D, Droz B, Greve P,etal.. Selenium deficiency risk predicted to increase under future climate change [J]. Proc. Natl. Acad. Sci. USA, 2017, 114(11): 2848-2853.
[9] Zawislanski P T, Benson S M, Terberg R,etal.. Selenium speciation, solubility, and mobility in land-disposed dredged sediments [J]. Environ. Sci. Technol., 2003, 37(11):2415-2420.
[10] Barrow N J, Whelan B R. Testing a mechanistic model. VII. The effects of pH and of electrolyte on the reaction of selenite and selenate with a soil [J]. Eur. J. Soil Sci., 1989, 40(1):17-28.
[11] 郭 璐, 满 楠, 梁东丽, 等. 小白菜对外源硒酸盐和亚硒酸盐动态吸收的差异及其机制研究[J]. 环境科学, 2013, 34(8): 3272-3279.
[12] Bitterli C, Bauelos G S, Schulin R. Use of transfer factors to characterize uptake of selenium by plants[J]. J. Geochem. Explor., 2010, 107(2):206-216.
[13] Neal R H, Sposito G, Holtzclaw K M,etal.. Selenite adsorption on alluvial soils: I. Soil composition and pH effects [J]. Soil Sci. Soc. Am. J., 1987, 51(5): 1161-1165.
[14] Baker A J M, Mcgrath S P, Reeves R D,etal.. Metal hyperaccumulator plants: A review of the ecology and physiology of a biological resource for phytoremediation of metal-polluted soils[C]. In: Terry N, Banuelos G, Gary S,etal.. Phytoremediation of contaminated soil and water[C]. Florida: Lewis Publishers, 2000, 85-107.
[15] Abrams M M, Shennan C, Zasoski R J,etal.. Selenomethionine uptake by wheat seedlings [J]. Agron. J., 1990, 82(6): 1127-1130.
[16] Kikkert J, Berkelaar E. Plant uptake and translocation of inorganic and organic forms of selenium [J]. Arch. Environ. Con. Tox., 2013, 65(3): 458-465.
[17] 李玉梅, 王根林, 李 艳, 等. 水稻对有机态硒的吸收与积累[J]. 中国农学通报, 2017, 33(10): 7-11.
[18] 姜 英, 曾昭海, 杨麒生,等. 植物硒吸收转化机制及生理作用研究进展[J]. 应用生态学报, 2016, 27(12):4067-4076.
[19] Cappa J J, Cappa P J, El Mehdawi A F,etal.. Characterization of selenium and sulfur accumulation across the genusStanleya(Brassicaceae): A field survey and common-garden experiment [J]. Am. J. Bot., 2014, 101(5):830-839.
[20] Pilon-Smits E A H. Selenium in plants [A]. In: Lüttge U, Cánovas F M, Matyssek R. Progress in Botany[M]. Springer International Publishing, 2015, 93-107.
[21] Statwick J, Majestic B J, Sher A A. Characterization and benefits of selenium uptake by anAstragalus[J]. Plant Soil, 2016, 404(1-2): 345-359.
[22] White P J, Bowen H C, Parmaguru P,etal.. Interactions between selenium and sulphur nutrition inArabidopsisthaliana[J]. J. Exp. Bot., 2004, 55(404):1927-1937.
[23] Schiavon M, Pilon M, Malagoli M,etal.. Exploring the importance of sulfate transporters and ATP sulphurylases for selenium hyperaccumulation-a comparison ofStanleyapinnataandBrassicajuncea(Brassicaceae) [J]. Front. Plant Sci., 2015, 6: 2-2.
[24] Buchner P, Stuiver C E E, Westerman S,etal.. Regulation of sulfate uptake and expression of sulfate transporter genes inBrassicaoleraceaas affected by atmospheric H2S and pedospheric sulfate nutrition [J]. Plant Physiol., 2004, 136(2): 3396-3408.
[25] Cao M J, Wang Z, Wirtz M,etal.. SULTR3; 1 is a chloroplast-localized sulfate transporter inArabidopsisthaliana[J]. Plant J., 2013, 73(4): 607-616.
[26] Zuber H, Davidian J C, Wirtz M,etal.. Sultr4; 1 mutant seeds ofArabidopsishave an enhanced sulphate content and modified proteome suggesting metabolic adaptations to altered sulphate compartmentalization [J]. BMC Plant Biol., 2010, 10: 78.
[27] Huang Q Q, Wang Q, Wan Y N,etal.. Application of X-ray absorption near edge spectroscopy to the study of the effect of sulphur on selenium uptake and assimilation in wheat seedlings [J]. Biol. Plantarum., 2017, 61(4): 726-732.
[28] Sors T G, Ellis D R, Salt D E. Selenium uptake, translocation, assimilation and metabolic fate in plants [J]. Photosynth. Res., 2005, 86(3): 373-389.
[29] Hopper J L, Parker D R. Plant availability of selenite and selenate as influenced by the competing ions phosphate and sulfate [J]. Plant Soil, 1999, 210(2): 199-207.
[30] Liu X, Zhao Z, Hu C,etal.. Effect of sulphate on selenium uptake and translocation in rape (BrassicanapusL.) supplied with selenate or selenite [J]. Plant Soil, 2016, 399(1-2): 295-304.
[31] Zayed A, Lytle C M, Terry N. Accumulation and volatilization of different chemical species of selenium by plants [J]. Planta, 1998, 206(2): 284-292.
[32] Arvy M P. Selenate and selenite uptake and translocation in bean plants (Phaseolusvulgaris) [J]. J. Exp. Bot., 1993, 44(6): 1083-1087.
[33] Terry N, Zayed A M, De Souza M P,etal.. Selenium in higher plants [J]. Ann. Rev. Plant Boil., 2000, 51(1): 401-432.
[34] Ulrich J M, Shrift A. Selenium absorption by excisedAstragalusroots [J]. Plant Physiol., 1968, 43(1): 14-20.
[35] Li H F, McGrath S P, Zhao F J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite [J]. New Phytol., 2008, 178(1): 92-102.
[36] Zhang L, Hu B, Li W,etal.. OsPT2, a phosphate transporter, is involved in the active uptake of selenite in rice [J]. New Phytol., 2014, 201(4): 1183-1191.
[37] 陈大清. 植物硒同化的研究进展及其耐硒突变体的筛选[J]. 氨基酸和生物资源, 2004, 26(2): 65-71.
[38] Jiang Y, Zeng Z H, Bu Y,etal.. Effects of selenium fertilizer on grain yield, Se uptake and distribution in common buckwheat (FagopyrumesculentumMoench) [J]. Plant Soil Environ., 2015, 61: 371-377.
[39] Mombo S, Schreck E, Dumat C,etal.. Bioaccessibility of selenium after human ingestion in relation to its chemical species and compartmentalization in maize [J]. Environ. Geochem. Health., 2016, 38(3): 869-883.
[40] 彭 琴, 李 哲, 梁东丽, 等. 不同作物对外源硒动态吸收, 转运的差异及其机制[J]. 环境科学, 2017, 38(4): 1667-1674.
[41] 覃爱苗, 唐 平, 余卫平. 硒在植物中的生物学效应[J]. 东北农业大学学报, 2011, 42(10):6-11.
[42] 李春霞, 曹 慧. 植物硒的营养特点及吸收转化机理研究进展[J]. 农业科学研究, 2006, 27(4): 72-76.
[43] 胡秋辉, 潘根兴, 丁瑞兴, 等. 富硒茶硒的浸出率及其化学性质的研究[J]. 中国农业科学, 1999, 32(5): 69-72.
[44] 赵 萍, 刘笑笑, 王 雅, 等. 富硒小麦提取物中硒含量及其抗氧化特性[J]. 食品科学, 2014(15): 94-98.
[45] Duncan E G, Maher W A, Jagtap R,etal.. Selenium speciation in wheat grain varies in the presence of nitrogen and sulphur fertilizers [J]. Environ. Geochem. Health., 2017, 39(4):955-966.
[46] Wilson L G, Bandurski R S. Enzymatic reactions involving sulfate, sulfite, selenate, and molybdate [J]. J. Biol. Chem., 1958, 233(4): 975-981.
[47] Pilon-Smits E A H, Quinn C F. Selenium metabolism in plants [A]. In: Hell R, Mendel R R. Cell Biology of Metals and Nutrients[M]. Springer Berlin Heidelberg, 2010, 225-241.
[48] Anderson J W. Selenium interactions in sulfur metabolism [JA]. In: Sulfur Nutrition and Assimilation in Higher Plants: Regulatory, Agricultural and Environmental Aspects[M]. The Netherlands The Hague: SPB Academic Publishing, 1993, 49-60.
[49] De Souza M P, Lytle C M, Mulholland M M,etal.. Selenium assimilation and volatilization from dimethylselenoniopropionate by Indian mustard [J]. Plant Physiol., 2000, 122(4): 1281-1288.
[50] El-Ramady H, Abdalla N, Taha H S,etal.. Selenium and nano-selenium in plant nutrition [J]. Environ. Chem. Lett., 2016, 14(1): 123-147.
广西富硒功能农业研究团队介绍
广西富硒功能农业研究团队主要研究方向为土壤硒资源高效利用及特色优质富硒农产品研发。自2013年以来,本团队主持国家自然科学基金、国家重点研发计划、广西创新驱动发展专项(科技重大专项)、广西科技重点研发计划、广西科学与技术开发计划、广西自然科学基金等硒相关项目50多项。有关硒学领域的主要研究成果包括:①揭示了以广西典型土壤为代表的土壤硒素特征;②阐明了硒在水稻、玉米等多种特色作物中的积累与分布特征;③探明了硒与重金属镉在水稻、玉米、火龙果与香蕉等亚热带水果、辣木等植物中的协同与拮抗生理效应;④研制出了一系列鳌合态生物有机硒素强化剂与土壤硒素生物活化剂;⑤指导涉农企业开发出了近50种特色优质富硒农产品,实现了产品的标准化定量富硒;⑥成功的从水稻、玉米、马铃薯、辣木等作物中提取出富硒蛋白,进行富硒精深加工型产品研发。
AdvanceonUptakeandTransformationofSeleniumfromSoiltoPlants
CHEN Jinping1, LIU Yongxian1*, ZENG Chengcheng1, PAN Liping1, LU Shiyang1, LAN Xiu2, HUANG Yanfei1, LIANG Panxia1, JIANG Zepu1, XING Ying1, LIAO Qing1, HUANG Taiqing1
1.AgriculturalResourcesandEnvironmentResearchInstitute,GuangxiAcademyofAgriculturalSciences,Nanning530007,China;2.CollegeofAgronomy,GuangxiUniversity,Nanning530005,China
As a trace element, Selenium (Se) is indispensable for humans, animals and some microorganisms. The main dietary Se for human comes from plants which are capable to uptake and transform Se from soil. It is significant for human health to enhance the level of Se accumulation in plants. For a better understanding of the influencing factors on the efficiency of plant accumulation of Se, the forms of Se in soil, main mechanisms relative to Se uptake, transformation and metabolism in plants were summarized in this paper. The prospects on the biofortification of Se were proposed at last.
selenium; plant; uptake; transformation
2017-08-04;接受日期2017-08-22
国家重点研发计划项目(2016YED0800705-01);广西重点研发计划项目(桂科AB16380088;桂科AB16380084);广西科学研究与技术开发计划项目(桂科合415104001-22);广西农业重点科技计划项目(201528;201604);广西富硒特色作物试验站(桂TS2016011);广西农业科学院基本科研业务专项(桂农科2017YZ03);广西农业科学院科技发展基金(2017JM07);广西农业科学院成果转化项目(2017NZ04);南宁市青秀区重点研发计划(2016039);南宁市西乡塘区科学研究与技术开发计划项目(2015312)资助。
陈锦平,硕士研究生,研究方向为植物逆境生理生态学。E-mail:447355175@qq.com。*通信作者:刘永贤,副研究员,研究方向为农业资源高效利用与功能性农产品开发。E-mail:liuyx27@163.com
10.19586/j.2095-2341.2017.0104
