考虑光盐交互作用的湖泊富营养化数学模型
2017-11-23李桂秋
张 彦,窦 明,李桂秋,孟 猛
考虑光盐交互作用的湖泊富营养化数学模型
张 彦1,2,窦 明1*,李桂秋1,孟 猛1
(1.郑州大学水利与环境学院,河南郑州 450001;2.中国农业科学院农田灌溉研究所,河南新乡 453002)
基于水体光学原理,确定了光照衰减系数与透明度之间的定量关系式;基于质量守恒原理,描述了氮磷营养盐与藻类之间的转化关系;耦合光因子和盐因子对藻类生长的驱动机制,建立了考虑光盐交互作用的富营养化数学模型.结合2015年4~7月在眉湖开展的水质监测数据,对模型进行了参数率定与验证.通过正交设计与情景模拟相结合,研究了光盐条件变化对藻类生长的驱动作用.结果表明,建立的富营养化模型能够较好的模拟不同光盐条件下藻类的生长趋势;低光照强度下营养盐浓度增加对藻类生长起到了抑制作用,营养盐浓度增加相同的倍数时TP浓度变化对藻类生长的影响作用要比TN浓度变化对藻类生长的影响作用大;整体上藻类的生长受到光照强度的影响高于营养盐,受到总磷的影响高于总氮,在设置的情境中光照强度、TP和TN浓度分别为89.6klx、0.168mg/L和2.72mg/L时最利于藻类生长.
富营养化模型;水体光学;营养盐转化;交互作用
富营养化是湖泊、水库和海湾等封闭性或半封闭性水体中氮、磷营养元素富集,水体生产力提高,某些特征性藻类(通常为蓝藻、绿藻)异常繁殖,使水质恶化的过程[1];对富营养化特征、差异分析以及主导机制目前进行了一定的研究[2-3].充足的营养盐是水体富营养化的必要条件,氮是藻细胞的组成物质,磷则参与了藻类的光合作用和能量转化过程[4-5].同时,水体中氮含量受藻类生长死亡和地球化学环境的强烈影响,磷含量也受到季节性水文过程变化的一定影响[6-7].除了营养盐条件外,光照条件是藻类生长的限制因子[8].光合作用是活体藻类的基本生命活动,而水体透明度变化对藻类生长有直接影响[9].相关研究表明太湖蓝藻暴发与氮磷浓度峰值出现时间的关系以及氮磷比对蓝藻优势形成的影响[10-11];当水体中营养盐充足时,光照和温度在一定范围内增加能明显促进水体富营养化的进程[12-13];水温和光照强度对微囊藻水华的生消过程也起到了一定的作用[14-15].总体来看,前期研究从不同角度探析了光照和营养盐对藻类生长的影响机制,但从光盐因子交互作用方面来研究其对藻类生长的驱动作用成果不多.本文在借鉴前期相关研究成果[16-22]的基础上,建立了考虑光盐交互作用的湖泊富营养化模型,以郑州大学人工湖眉湖为研究对象,研究在光照和营养盐条件变化下的湖泊藻类生长规律及主要限制因子,其研究成果可为湖泊水环境治理工作提供参考.
1 模型建立
1.1 模型构建基本原理
本文提出的富营养化模型主要由二维湖泊水动力学模型[23]和考虑光盐因子驱动的富营养化模型两部分组成.水动力模型是模拟溶质随水流迁移转化过程的基础,结合研究水域特点进行计算单元的划分,并应用二维非恒定流圣维南方程组来模拟水位、流量和流速等水动力学指标的变化过程.富营养化模型是在作者前期研制的汉江富营养化动态模型[21]基础上,增加了对浊度、透明度指标的模拟,并建立了光照衰减因子与透明度之间的定量关系,此外耦合营养盐在“溶解相-生物相-底泥相”等不同相态间转化机制的描述[24-25],由此反映光盐条件变化对藻类生长死亡过程的影响.其内部作用机制如图1所示.
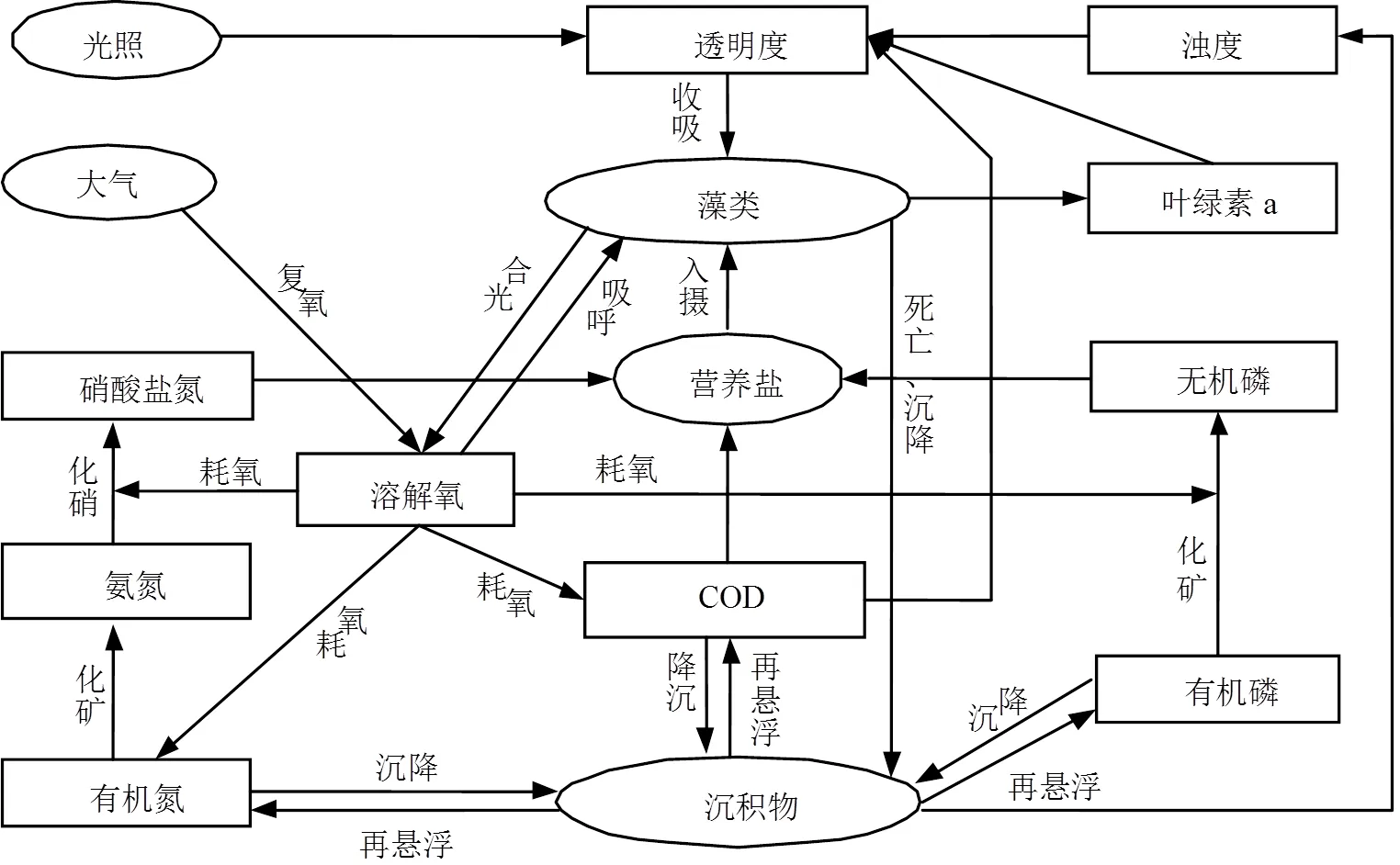
图1 富营养化数学模型结构框架
富营养化模型的基本方程是平移—扩散质量迁移方程,该方程能够描述水质指标在时间和空间上的变化规律.在方程里除了平移和扩散项外,还包括由生物、化学以及物理作用引起的源漏项. 水质变量的基本表达式为:
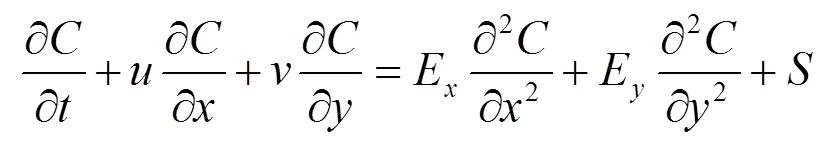
式中:为水质指标的浓度,mg/L;、分别为流速在、方向上的分量,m/s;E、E分别为扩散系数在、方向上的分量,m2/s;为总源漏项,正为源、负为漏,g/(m3·d).
藻类生长动力学过程在水体富营养化模型中占有重要的位置,直接影响到其他水质变量的转化过程.藻类的源漏项1可表示为:

式中:G1为藻类的生长率,d-1;D1为藻类的衰减率,d-1;K4为藻类的降解率,d-1;PYT为藻类在水体中的浓度,cell/mL.
藻类生长率G1是一个复杂的非线性函数,主要受水温、光照强度和营养盐浓度等外部环境条件的制约.Di-Toro等[26]研究认为,藻类生长率G1可表示为上述因素的乘积,即

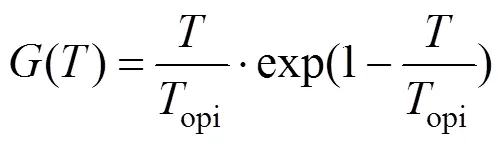
1.2 光因子驱动机制描述
受天气、水体透明度以及水深等因素的影响,水下的光照强度变化较大,因此需弄清光强在水下变化规律.水下光照强度的变化规律可用比尔定律来表示[27]:
研究表明[27],水体透明度对水下光照强度的衰减具有主导作用,而透明度自身又受到叶绿素a、COD等有关水质指标的影响.因此,本文根据实际监测的水下1cm和20cm处光照强度,结合式(4)求出研究水域的光照衰减系数,并建立光照衰减系数与水体透明度的数学关系式.针对水体透明度的变化规律研究,首先根据实验数据识别影响透明度变化的主要水质因子,进而建立水体透明度与主要因子之间的定量关系式,再将该关系式耦合到湖泊富营养化模型中,由此来模拟光照强度变化对藻类生长过程的影响效果.
1.3 盐因子驱动机制描述
水体中氮、磷等营养物质富集是水体富营养化以及藻类生长的重要驱动因素.其中,氮循环过程从藻类对氨氮、硝酸盐氮等无机氮的摄入开始,通过该环节藻类获取能量、合成自身细胞组织,当藻类死亡后沉降到沉积物中,分解转化为有机氮,有机氮通过矿化作用转化为氨氮,氨氮再通过硝化作用转化为硝酸盐氮,由此进行不断地循环转化.各类氮素的源漏项可表示为:

式中:71为有机氮的矿化系数,d-1;71为矿化系数的温度系数,无量纲;12为硝化系数,d-1;12为硝化系数的温度系数,无量纲;NC为藻类的氮碳比,无量纲.
式中:PC为藻类的磷碳比,无量纲.
1.4 光盐交互作用情景设计
采用正交设计和情景模拟相结合的方式,来研究光照和营养盐条件变化对藻类生长的驱动效果.正交设计是一种研究多因素影响的试验设计方法,它是从全面试验中挑选出部分有代表性的因素组合来进行试验分析[28].其特点是仅通过少数代表性很强的试验即可摸清各因子对试验结果的影响程度,筛选出较好的试验条件组合[29].为了描述光盐因子交互作用下藻类的生长规律,分别设定原始监测光照强度的0.5倍、1倍、1.5倍和2倍4种情况,总氮或总磷为原始监测数据的1倍、2倍、3倍和4倍4种情况,进而根据正交试验设计原理,得到在3个因素(光照强度、总磷浓度和总氮浓度)、4个不同水平条件下的16组光盐条件组合情景(表1).
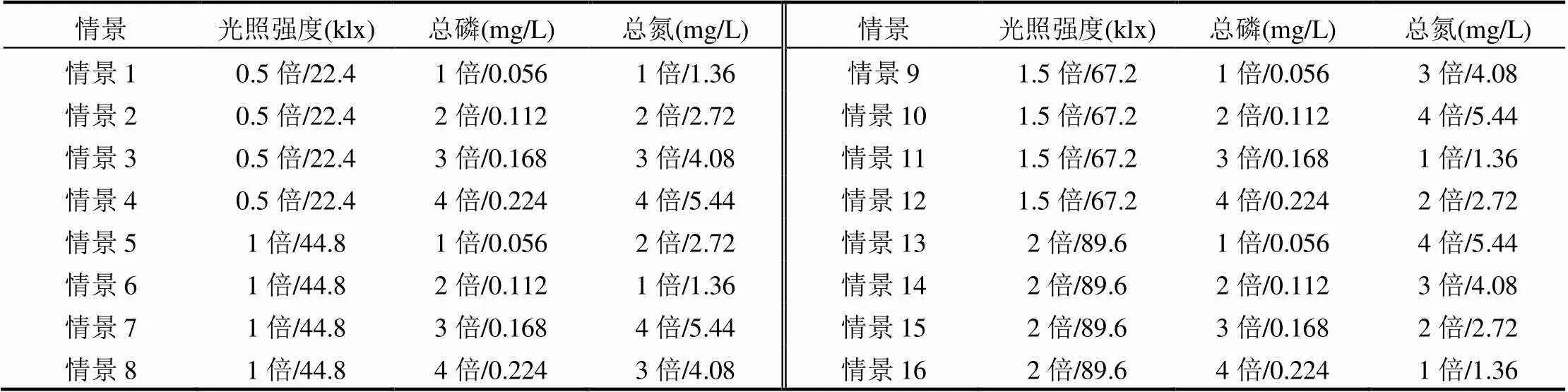
表1 光盐因子交互影响情景设计
注:各指标均用“倍数/平均值”表示,其中光照强度、总磷和总氮平均值表示为各情景下指标系列监测数据的平均值.
2 应用研究
2.1 实验设计与数据收集
本文以郑州大学新校区人工湖—眉湖为研究对象,该湖是一个典型的人工景观湖泊,湖泊水面宽度30~60m,湖长约为500m,湖面面积约为2.2万m2.眉湖中配置了水循环系统,包括局部喷泉和上扬式曝气管循环和整体的南北循环.供水水源包括地下水和雨水两部分,并以地下水补给为主,雨水则来自处理过的贮存雨水.湖水分为北、中、南3 段,中段较长且水流缓慢,北端地势较高,南端设有高低阶梯.监测实验的范围为整个湖面,共设置了5个监测断面(I、II、III、IV、V)、5个取样点(1#、2#、3#、4#、5#),监测断面以及取样点的布设如图2所示.

图2 监测断面及取样点分布示意
2015年4~7月,作者在眉湖先后开展了10次监测实验,对I、II、III、IV和V断面进行了现场监测.监测指标包括光照强度、透明度(SD)、浊度、溶解氧(DO)、水温、叶绿素a(Chl-a)、藻类(PYT)、水深和流速;4月30日、5月28日和6月29日分别在取样点1#、2#、3#、4#和5#处进行水体取样,5月16日和6月15日分别在取样点2#、3#和5#处进行水体取样,共采集了21个水样,对样品的化学需氧量(COD)、总磷(TP)、总氮(TN)、氨氮(NH3-N)、硝氮(NO3--N)等指标进行了检测.其中光照强度由便携式光照测定仪进行检测,主要测量水下1cm和20cm处的光照强度;透明度由SD-20塞氏盘测定,浊度由WGZ–2B便携式浊度仪测定,藻类和叶绿素a由HACH Hydrolab DS5X多参数水质监测仪测定,其他水质指标由HACH水质监测组件测定;水样指标的检测分析方法主要参照《水和废水监测分析方法》中的相关方法.
2.2 光照衰减系数的获取
光照衰减系数是计算水下光强的重要参数. 根据实验监测的水下1cm处和20cm处光照强度,应用式4计算出水体中光照衰减系数.通过分析光照衰减系数与水体透明度的相关性,得到二者间的拟合方程及拟合图,具体如式(10)和图3所示.拟合方程的拟合度为0.89,相关性较好.由式(9)可知,光照衰减系数与水体透明度呈负相关关系,即当水体透明度增大时,光照衰减系数减小.
借助实验数据,对水体透明度和其他水质指标之间进行相关性分析后发现,水体透明度与浊度、Chl-a、COD和TN均呈现显著性相关,且相关系数也比较大.依据水体透明度与各单因子间的拟合关系,对浊度、Chl-a、COD和TN数据处理后再进行多元线性回归分析,得到数据处理前和处理后不同的多元线性拟合方程,其各因子和常量的非标准系数及显著性如表2所示.

由表2可得,眉湖水体透明度与其影响因子间的多元回归方程如下,其中式(11)为各因子处理前即原始数据下得到的多元回归方程;式(12)为各因子处理后的多元回归方程,即水体透明度取自然对数值、浊度取四次方根、叶绿素和COD取以10为底的对数值.
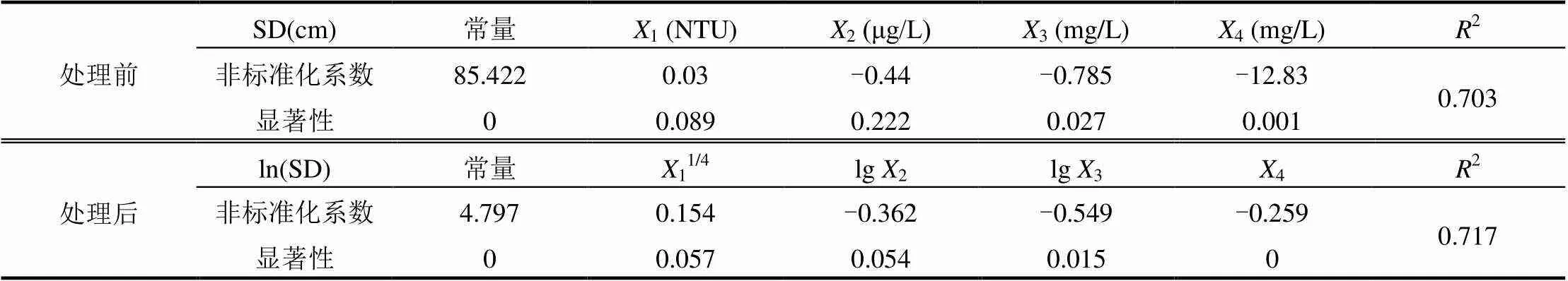
表2 常量和影响因子的非标准系数和显著性
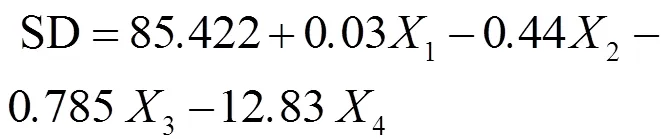
式中:SD为水体透明度,cm;1为浊度,NTU;2为叶绿素含量,μg/L;3为COD浓度,mg/L;4为TN浓度,mg/L.
通过对比发现,处理前的水体透明度拟合方程式(11)与处理后的拟合方程式(12)拟合度分别达到了0.703和0.717,均满足要求,但总体来看处理后的拟合效果更好.根据处理前各因子的多元线性回归分析可知,浊度和Chl-a的非标准化系数的显著性大小分别为0.089和0.222,显著性水平均大于0.05,说明在此回归方程中浊度和Chl-a的非标准化系数的显著性不太明显;而根据处理后数据的多元线性回归分析可知,浊度和Chl-a的非标准化系数的显著性大小分别为0.057和0.054,显著性水平已经非常接近0.05,说明在处理后的回归方程中浊度和Chl-a的非标准化系数的显著性较为明显.因此,在后续的计算过程中采用监测数据处理后得到的多元线性回归方程来模拟眉湖水体透明度指标的变化情况.
2.3 参数率定与模型验证
2.3.1 水动力学模型参数率定与验证 根据2015年4~7月期间眉湖流量、水位等实测资料,选取不同的湖底糙率和阻力系数,并设定监测断面V为入流断面、监测断面I为出流断面,来模拟各个监测断面的水动力学指标数值.通过模型率定,当眉湖湖底糙率和阻力系数分别为0.024和0.6时,监测断面的流量和水位的模拟结果比较理想.图4是监测断面II和IV的模拟结果验证情况.
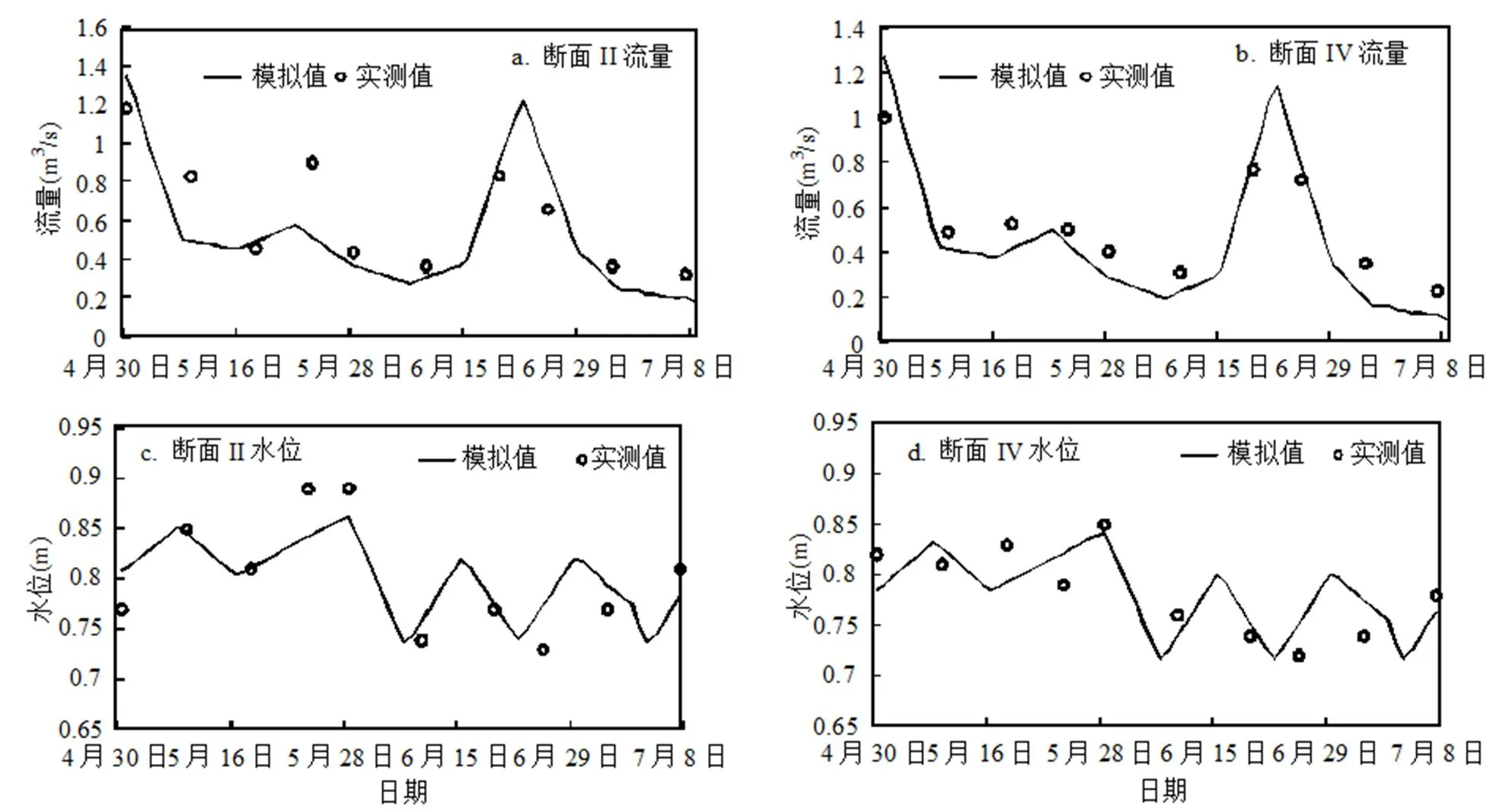
图4 水动力学模型参数率定和验证结果

表3 富营养化数学模型的参数项
注:*表示该项为非恒定值; —表示该项为无量纲参数.
采用最大相对误差和平均相对误差来分析水动力模型的模拟效果:监测断面II和IV流量的最大相对误差分别为16.12%和28.5%,平均相对误差分别为11.63%和19.8%;水位的最大相对误差分别为5.8%和4.78%,平均相对误差分别为2.96%和3.2%.总体来看,模型计算结果的误差相对较小,模拟精度满足要求.
2.3.2 富营养化模型参数率定与验证 在开展富营养化模型模拟时,以4月30日各监测断面的实测资料作为初始条件,以监测断面V的水质浓度变化过程作为边界条件,并将模型的时间步长设定为1d.由于本次建立的富营养化模型参数较多,逐个率定比较繁琐,因此首先进行参数敏感性分析,再根据相关研究成果给出的参数推荐值作为初始值,对敏感性参数不断调整其数值大小,并根据模拟结果分析取值合理性,而对不敏感性参数则维持初始值不变,最后将敏感性参数作为整体统一进行调整,以使模型的整体拟合效果达到最优,此时得到的参数值即为最终率定的参数(表3).
同样采用最大相对误差和平均相对误差来分析水质模型的模拟效果:监测断面II和IV的PYT最大相对误差分别为12.55%和13.75%,平均相对误差分别为5.46%和7.35%;TN的最大相对误差分别为14.35%和9.18%,平均相对误差分别为4.96%和6.28%;TP的最大相对误差分别为28.71%和12.5%,平均相对误差分别为9.35%和7.72%;COD的最大相对误差分别为10%和11.18%,平均相对误差分别为6.89%和7.27%;SD的最大相对误差分别为29.02%和17.03%,平均相对误差分别为14.15%和7.83%.总体来看,模型计算结果的误差相对较小,模拟精度满足要求.监测断面II和IV模拟验证结果如图5所示.
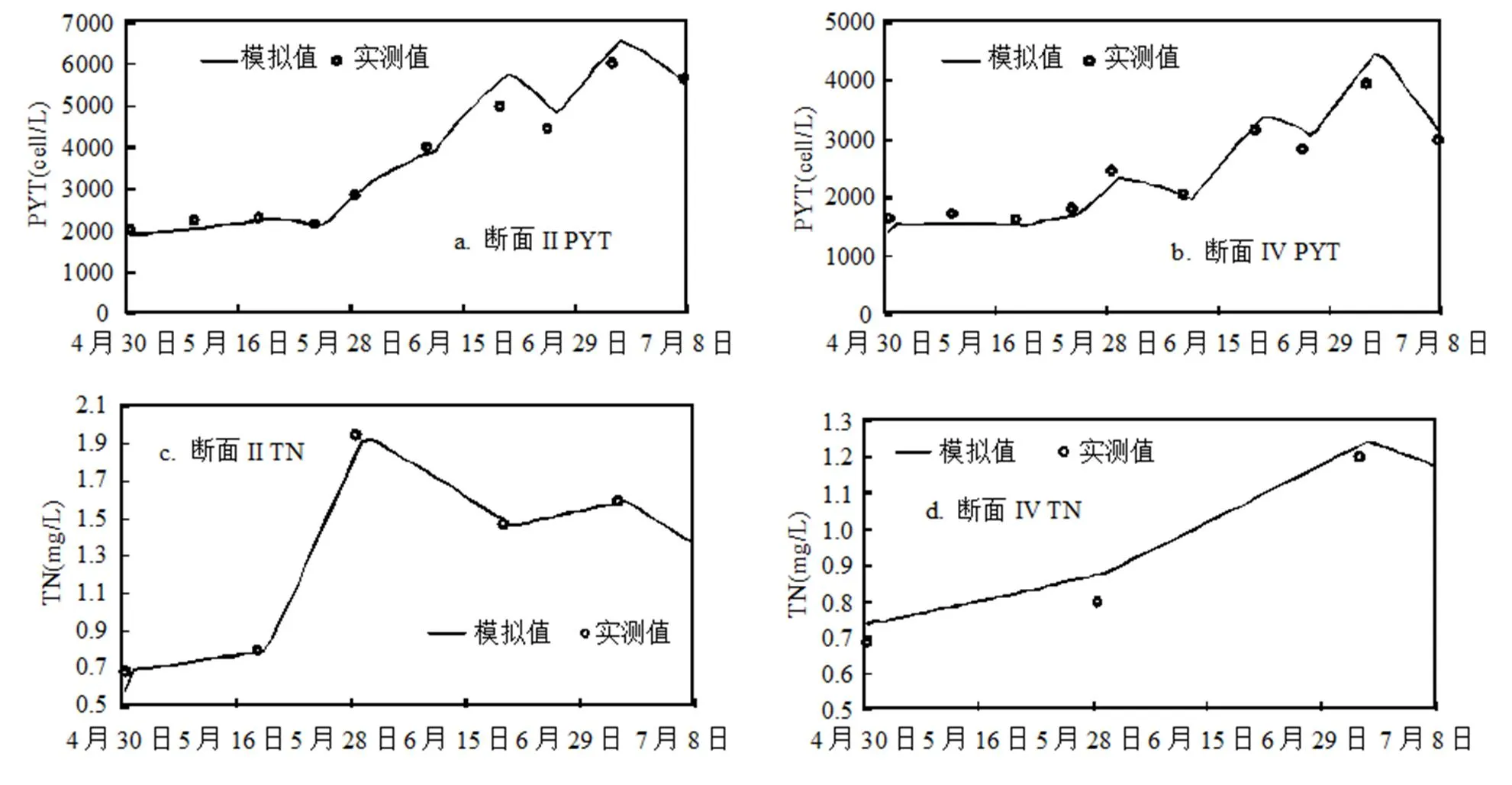
根据模拟结果可知,监测断面II和IV处PYT模拟的平均含量分别为3845,2492cell/mL,TN浓度模拟的平均值分别为1.34,0.97mg/L,TP浓度模拟的平均值分别为0.056,0.039mg/L,COD浓度模拟的平均值分别为27.76,17.46mg/L,SD模拟的平均值分别为39.1,60.9cm.监测断面II处各指标的模拟平均值比监测断面IV处的模拟平均值大,这是由于监测断面II处圈养了禽类和观赏鱼类,投放饵料和食物较多,使水体中的COD、TN和TP浓度增加即水体中营养物质增加,SD减小,进而促进了藻类生长使监测断面II处水体中藻类含量增加;而监测断面IV处有大量的水生植物,对水体具有一定的净化作用,降解了水体中相关的营养物质,在一定程度上抑制了藻类生长.对于PYT,整体上其含量呈现逐步上升的趋势,6月份藻类的含量明显比5月份藻类含量高,说明气象条件的变化对藻类生长也有较大的影响作用.监测断面II处的TN、TP和COD浓度的模拟结果呈现出先增加后减小的波动趋势;监测断面IV处TN浓度的模拟结果呈现出增加的趋势,而TP和COD浓度的模拟结果呈现出先减小后增加的波动趋势;SD在监测断面IV整体上呈现减小的趋势,而在监测断面II呈现出先减小后增加的趋势.
2.4 光盐交互对藻类驱动作用分析
根据正交试验设计得到光照强度和营养盐交互的16种情景,并通过富营养化模型模拟得到监测断面II光照强度和营养盐交互情景下眉湖中藻类生长的规律,具体情况如图6所示,16种情境下藻类模拟的平均值及峰值如表4所示.
根据文献[22-23],当藻类含量超过10000cell/ mL时则湖泊会出现水华现象.由表4给出的各情景藻类含量峰值可知,情景1、情景2、情景5、情景6、情景8、情景9以及情景11~16在模拟期中的某段时间内出现了水华现象,其中在情景15条件下模拟藻类含量的平均值和峰值均最大,分别为22384,49233cell/mL,说明在该情景条件下(即光照强度89.6klx、TP和TN浓度分别为0.168,2.72mg/L),眉湖水体中藻类生长的最快.而在情景4条件下藻类生长模拟的平均值和峰值均最小,分别为3663,6261cell/mL,说明此时(光照强度22.4klx、TP和TN浓度分别为0.224, 5.44mg/L),眉湖水体中藻类生长的最慢,即低光照强度和高营养盐条件下对藻类生长的抑制作用明显.从情景1~4可知,当光照强度的平均值为22.4klx时,随着TP和TN浓度的增加,藻类含量模拟的平均值呈现减小的趋势,说明在光照强度较低的情况下,营养盐浓度增加对藻类生长起到了抑制作用,即此时光照强度对藻类的生长起到了重要的作用.从情景2和情景5、情景12和情景15对比分析,TN浓度为2.72mg/L时,光照强度增大时藻类模拟的平均值比TP浓度增加时藻类模拟的平均值高,说明光照强度变化对藻类生长的影响作用比TP浓度变化对藻类生长的影响作用大.从情景5和情景6、情景7和情景8对比可知,当TP和TN浓度都增加相同的倍数时,TP浓度增加时藻类模拟的平均值比TN浓度增加时藻类模拟的平均值高,即TP浓度变化对藻类生长的影响作用要比TN浓度变化对藻类生长的影响作用大,说明眉湖水体中促进藻类生长的营养盐TP占较大的比重,另外在情景9和情景11、情景10和情景12、情景13和情景16、情景14和情景15的对比中也呈现出此种现象.

图6 不同光盐交互下藻类生长模拟结果

表4 不同情景下藻类模拟的平均值及峰值
从情景1、情景5、情景9和情景13可知,当TP浓度为0.056mg/L时,在一定范围内随着光照强度和TN浓度的增加水体中藻类模拟的平均值增加,而光照强度和TN浓度超出一定范围后水体中藻类的含量出现减少现象,说明一定范围内光照强度和TN浓度有促进藻类生长的作用,而光照强度和TN浓度相对较大时又呈现出抑制藻类生长的作用.从情景3、情景7、情景11和情景15可知,当TP浓度为0.168mg/L时,情景3和情景7条件下藻类模拟的平均值要比情景11和情景15条件下藻类模拟的平均值低,即当光照强度较高和TN浓度较低时对藻类生长的促进作用要高于光照强度较低和TN浓度较高时对藻类生长的促进作用,说明当TP浓度增大到一定程度时,低光照强度和高浓度TN对藻类生长的抑制作用更加显著.从情景1、情景6、情景11和情景16可知,当TN浓度为1.36mg/L时,在一定范围内随着光照强度和TP浓度的增加水体中藻类模拟的平均值增加,而光照强度和TP浓度超出一定范围后水体中藻类模拟的平均值出现减少现象,说明一定范围内光照强度和TP浓度有促进藻类生长的作用,而光照强度和TP浓度相对较大时又呈现出抑制藻类生长的作用.从情景4、情景7、情景10和情景13可知,当TN浓度为5.44mg/L时,情景4和情景7条件下藻类模拟的平均值要比情景10和情景13条件下藻类模拟的平均值低,即当光照强度较高和TP浓度较低时对藻类生长的促进作用要高于光照强度较低和TP浓度较高时对藻类生长的促进作用,说明当TN浓度增大到一定程度时,低光照强度和高浓度TP对藻类生长的抑制作用更加显著.
总体来说,在光照强度和营养盐的适宜范围内均有促进藻类生长的作用,而光照强度或营养盐超出一定范围后又对藻类生长起到了抑制作用,这与相关研究成果是一致的[9,30-31];通过以上对比分析可知光照强度对藻类生长的影响作用比营养盐的影响作用大,总磷对藻类生长的影响作用比总氮的影响作用大.但在不同水平光照强度和营养盐交互作用下,由于它们之间的相互影响对藻类生长起到了不同的影响作用,本文主要是通过设置不同的光盐交互情景初步模拟了藻类生长的情况,因此想要更加清晰的了解光照强度和营养盐交互作用对藻类生长的影响,首先要延长监测数据的系列以使富营养化模型更加精确,其次要增加光照强度和营养盐的不同水平以通过更多的光盐交互情景来说明光照强度和营养盐下对藻类生长的影响作用.
3 结论
3.1 结合水动力学和富营养化基本模型,在建立水体透明度与光照衰减系数、水体透明度与主要因子数学关系的基础上,构建了考虑光盐影响的湖泊富营养化数学模型,并利用实验数据对模型进行了参数率定和验证.
3.2 经验证结果表明构建的模型在模拟眉湖水动力指标和水体富营养化指标时具有较好的精度,能够较好的描述眉湖富营养化水质指标的变化规律.
3.3 通过不同光盐交互情景模拟分析初步了解了光盐交互对藻类生长的驱动机制,即在光照强度和营养盐的适宜范围内均有促进藻类生长的作用,而光照强度和营养盐超出一定范围后又对藻类生长起到了抑制作用.光照强度对藻类生长的影响作用比营养盐的影响作用大,总磷对藻类生长的影响作用比总氮的影响作用大.
[1] 秦伯强,高 光,朱广伟,等.湖泊富营养化及其生态系统响应[J]. 科学通报, 2013,58(10):855-864.
[2] 李俊龙,郑丙辉,张铃松,等.中国主要河口海湾富营养化特征及差异分析[J]. 中国环境科学, 2016,36(2):506-516.
[3] 李 颖,曹文志,张玉珍,等.九龙江流域上游浅水湖泊富营养化机制[J]. 中国环境科学, 2012,32(5):906-911.
[4] Smayda T J. Reflections on the ballast water dispersal-harmful algal bloom paradigm [J]. Harmful Algae, 2007,6(4):601-622.
[5] 陈能汪,章颖瑶,李延风.我国淡水藻华长期变动特征综合分析[J]. 生态环境学报, 2010,19(8):1994-1998.
[6] Schindler D W. Evolution of phosphorus limitation in lakes [J]. Science, 1977,195:260-262.
[7] Oliver A A, Dahlgren R A, Deas M L. The upside-down river: Reservoirs, algal blooms, and tributaries affect temporal and spatial patterns in nitrogen and phosphorus in the Klamath River, USA [J]. Journal of Hydrology, 2014,519:164-176.
[8] Ihnken S, Eggert A, Beardall J. Exposure times in rapid light curves affect photosynthetic parameters in algae [J]. Aquatic Botany, 2010,93(3):185-194.
[9] 王 菁,陈家长,孟顺龙.环境因素对藻类生长竞争的影响[J]. 中国农学通报, 2013,29(17):52-56.
[10] 杨贵山,王德建.太湖流域经济发展.水环境.水灾害 [M]. 北京:科学出版社, 2003:20-80.
[11] 许 海,朱广伟,秦伯强,等.氮磷比对水华蓝藻优势形成的影响[J]. 中国环境科学, 2011,31(10):1676-1683.
[12] 刘 国,何 娟,李 兵,等.光照对底泥中氮、磷动态变化的影响研究[J]. 工业安全与环保, 2015,41(8):50-53.
[13] 刘玉生,韩 梅,梁占彬,等.光照、温度和营养盐对滇池微囊藻生长的影响[J]. 环境科学研究, 1995,8(6):7-11.
[14] 黄钰铃,陈明曦.水华生消模拟机其溶解氧变化过程分析[J]. 环境科学与技术, 2013,36(10):67-72.
[15] 张艳晴,杨桂军,秦伯强,等.光照强度对水华微囊藻()群体大小增长的影响[J]. 湖泊科学, 2014,26(4): 559-566.
[16] Lorenzen M W, Smith O J, Kimmel L V. A long-term phorphorus model for lakes: Application of Lake Washington. Canale R P. Modeling Biochemical Processes in Aquatic Ecosystems [M]. Michigan: Ann Arbor Science, 1976:75-92.
[17] Imboden D M, Gachter R A dynamic lake model for trophic state prediction [J]. Ecological Modeling, 1978,4(2/3):77-98.
[18] Cerco C F, Cole T. Three-dimensional eutrophication model of Chesapeake bay [J]. Environmental Engineering, 1993,119(6): 1006-1025.
[19] Lino J, Alvarez-Vazquez, Francisco J. Fernandez, et al. An Arbitrary Lagrangian Eulerian formulation for a 3D eutrophication model in a moving domain [J]. Journal of Mathematical Analysis and Applications, 2010,366:319–334.
[20] Nyholm N. A simulation model for phytoplankton growth and nutrient cycling in eutrophic, shallow lakes [J]. Ecological Modeling, 1978,4(2/3):279-310.
[21] 夏 军,窦 明.汉江富营养化动态模型研究[J]. 重庆环境科学, 2001,23(1):20-23.
[22] 窦 明,谢 平,夏 军,等.南水北调中线工程对汉江水华影响研究[J]. 水科学进展, 2002,13(6):714-718.
[23] 李畅游,史小红.干旱半干旱地区湖泊二维水动力模型[J]. 水利学报, 2007,38(12):1482-1488.
[24] 窦 明,米庆彬,李桂秋,等.闸控河段水质转化机制研究I:模型研制[J]. 水利学报, 2016,47(4):527-536.
[25] 窦 明,米庆彬,李桂秋,等.闸控河段水质转化机制研究II:主导反应机制[J]. 水利学报, 2016,47(5):635-643.
[26] Di Tiro D M. Applicability of cellular equilibrium and Monod theory to phytoplankton growth kinetics [J]. Ecological Modeling, 1980,8:201-218.
[27] 王 华,逢 勇,刘申宝,等.沉水植物生长因子研究进展[J]. 生态学报, 2008,28(8):3958-3968.
[28] 唐启义.DPS数据处理系统—实验设计、统计分析及数据挖掘 [M]. 北京:科学出版社, 2010:238-242.
[29] 杨澄宇,代 超,伊 璇,等.基于正交设计及EFDC模型的湖泊流域总量控制—以滇池流域为例 [J]. 中国环境科学, 2016, 36(12):3696-3702.
[30] 宋玉芝,杨美玖,秦伯强.苦草对富营养化水体中氮磷营养盐的生理响应 [J]. 环境科学, 2011,32(9):2569-2575.
[31] 于 婷,戴景峻,雷腊梅,等.温度、光照强度及硝酸盐对拟柱孢藻(N8)生长的影响 [J]. 湖泊科学, 2014,26(3):441-446.
A mathematical model of eutrophication lake accounting for the light-nutrients interaction.
ZHANG Yan1,2, DOU Ming1*, LI Gui-qiu1, MENG Meng1
(1.College of Water Conservancy and Environment, Zhengzhou University, Zhengzhou 450001, China;2.Farmland Irrigation Research Institute, Chinese Academy of Agricultural Sciences, Xinxiang 453002, China)., 2017,37(11):4312~4322
A quantitative relation between light attenuation coefficient and water transparency was established based on the principle of water optics; described the relationship between nitrogen-phosphorus nutrients and algae based on the principle of mass conservation; constructed an eutrophication model accounting for the interaction effect of light and nutrients, which coupled the driving mechanisms of light and nutrients on algae growth. The model was calibrated and verified using water quality data monitored from April to July 2015 in the Meihu Lake. The model was then used to analyse the effect of change in light-nutrient content on algae growth, under different scenarios. Results showed that the algae had a good growth tendency under different light-nutrient conditions which was simulated by the established eutrophication model; there was an inhibitory action for algae under low light intensity when the nutrient concentrations increased, the effect of TP concentration was more important than TN concentration on algal growth when nutrient concentrations increased the same multiple; as a whole, algae growth is more affected by light intensity than nutrients, and more affected by total phosphorus than total nitrogen. The best scenario for algae growth is where illumination is 89.6klx, TP concentration is 0.168mg/L and TN concentration is 2.72mg/L.
eutrophication model;water optics;nutrient transformation;interaction
X524
A
1000-6923(2017)11-4312-11
张 彦(1989-),男,河南新乡人,研究实习员,硕士,主要从事水资源和水环境研究.发表论文10余篇.
2017-05-05
国家自然科学基金面上项目(51679218);河南省高校科技创新人才支持计划项目(17HASTIT031);郑州大学优秀青年教师发展基金资助项目(1521323001)
* 责任作者, 教授, dou_ming@163.com
