北江鱼类群落结构及其与环境因子的关系
2017-11-21高天扬彭宁东张少平罗锦桢王春晖
高天扬,谢 迪,彭宁东,李 潮,张少平,罗锦桢,王春晖,赵 俊
(华南师范大学生命科学学院,广东省水产健康安全养殖重点实验室,广东省优质环保水产养殖工程技术研究中心,广州市亚热带生物多样性与环境生物监测重点实验室,广州 510631)
北江鱼类群落结构及其与环境因子的关系
高天扬,谢 迪,彭宁东,李 潮,张少平,罗锦桢,王春晖,赵 俊
(华南师范大学生命科学学院,广东省水产健康安全养殖重点实验室,广东省优质环保水产养殖工程技术研究中心,广州市亚热带生物多样性与环境生物监测重点实验室,广州 510631)

北江;鱼类群落结构;季节变化;典型对应分析
北江是珠江流域的第二大水系,发源于江西省信丰县,主流称浈江,在韶关市沙尾洲与发源于湖南省临武县的武江汇合后始称北江。干流全长573 km,流域面积52 068 km2,在韶关市沙洲尾以上为上游,韶关市沙洲尾至清远市飞来峡为中游,飞来峡至三水市思贤窖为下游[1]。
有关北江鱼类资源研究性报道相对较少,最具有代表性的著作有《珠江水系北江渔业资源》[1]和《广东淡水鱼类资源调查与研究》[2]。对北江鱼类群落结构特征及其与环境因子之间关系的研究未见报道,因此,分析北江鱼类群落结构现状及对北江鱼类群落季节性变化与环境因子之间关系进行探讨,以揭示影响北江鱼类群落结构季节性变化的主要环境因子,具有十分重要的意义。
1 材料与方法
1.1 调查方法
参照《内陆水域渔业自然资源调查手册》[3]的方法。共设置5个调查站点,其中干流4个,分别是韶关站点(24.79°N,113.59°E)、连江口站(24.01°N,113.29°E)、清远站点(23.68°N,113.05°E)和石角站点(23.54°N,112.96°E),一级支流1个,位于连江的阳山站点(24.47°N,112.63°E)(图1)。鱼类资源采集采用常规性的刺网、地笼网及撒网,种类鉴定依据《广东淡水鱼类志》[4]。水温(Tem)、溶氧量(DO)、盐度(Sal)、化学耗氧量(CODMn)、氨氮含量(NH3-N)和pH值共6项数据来源于中华人民共和国环境保护部数据中心(http://datacenter.mep.)。
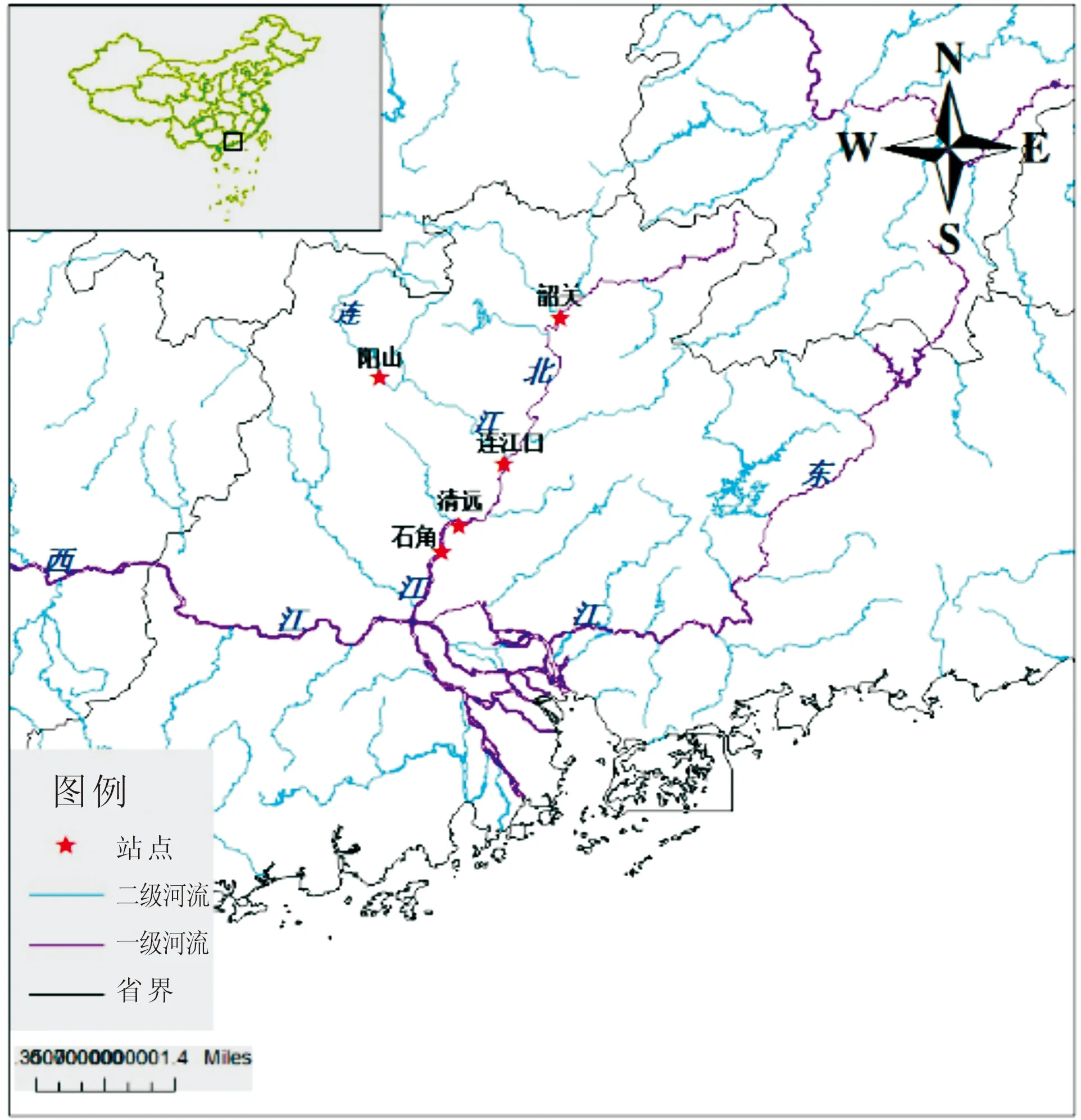
图1 北江鱼类资源调查站点图Fig.1 Beijiang River fish resources survey stance
1.2 数据分析
1.2.1 群落优势种分析
鱼类优势度采用Pinkas相对重要性指数(index of relative importance,IRI)衡量[5]。IRI=(N+W)×10 000,N为某一种鱼尾数占总尾数百分比,W%为某一种鱼重量占总重量百分比。IRI≥1 000的种类定义为优势种,500≤IRI<1 000定义为常见种,100≤IRI<500定义为一般种,10≤IRI<100定义为少见种,IRI<10定义为稀有种。
1.2.2 群落结构相似性分析
根据不同站点鱼类相对丰度数据建立矩阵,其中横坐标为种类,纵坐标为站点。以Bray-Curtis相似性系数为基础构建相似性矩阵,采用等级聚类的分类方法和非度量多维标度(non-metric multidimensional scaling,简称NMDS)的排序方法分别构建聚类分析图和NMDS图。NMDS分析结果的优劣由协强系数(stress)值来衡量[6]。
1.2.3 鱼类群落结构稳定性分析
鱼类群落结构稳定性分析基于Warwick的丰度-生物量比较曲线(Abundance-Biomass Comparison Curves,简称ABC 曲线)方法。ABC 曲线是在同一坐标系中比较鱼类数量优势度曲线和生物量优势度曲线,通过两条曲线的分布情况来分析鱼类群落在不同干扰状况下的特征[7-10]。
用W统计值(W-statistic) 作为ABC 曲线方法的一个统计量,其公式为:
式中Bi和Ai为种类序号对应生物量和数量的累积百分比,S为出现物种数。当生物量优势曲线在数量优势度曲线之上时,W为正,反之W为负。
1.2.4 鱼类群落结构季节变化与环境因子之间的相互关系
为探讨鱼类群落结构季节变化与环境因子之间的关系,首先,对逐月调查数据进行束缚型排序。水温(Tem)、溶氧量(DO)等作为环境数据,鱼类种类相对丰度作为物种数据,构建环境因子与鱼类相对丰度之间的矩阵关系。
其次,对物种数据进行趋势对应分析(DCA),以判断采用哪种模型(线性模型或者是单峰模型)进行典型对应分析(CCA)更合适[11-12]。为优化分析,本研究对环境因子数据进行lg(x+1)转化,去除波动因子>20的环境因子[13],利用Monte Carlo检验筛选出具有重要且独立作用的最少变量组合用于最终的典型对应分析(CCA)。
数据分析软件为Canoco5.0 for Windows软件包[14]。
2 结果分析
2.1 鱼类种类组成
本研究共采集鱼类179 076尾,16 101.93 kg,隶属于8目16科56属77种。其中鲤形目44种,占总数57.1%;鲈形目10种,占总数13.0%;鲇形目8种,占总数10.3%;其余鳉形目、合鳃鱼目、鲱形目、鲑形目和鲻形目种数均少于5。对其按相对重要性指数(IRI)排名,结果显示、鲤和鲮属于优势种,麦瑞加拉鲮、鲢等9种鱼为常见种,赤眼鳟、黄颡鱼等11种鱼为一般种,大眼鳜、斑鳜等26种鱼为少见种,粗唇鮠、丁鱥等28种鱼为稀有种(附件1)。
2.2 鱼类群落生态类型
按洄游类型将北江鱼类分为江海洄游性、河湖洄游性和定居性;按摄食食性分为植食性、肉食性和杂食性。按洄游类型来分,北江鱼类中数量和生物量方面均以鲤、鲫、黄颡鱼等定居性鱼类最多,鲢、鳙、草鱼等河湖洄游性次之,以花鰶等为代表的江海洄游性鱼类最少;按摄食食性来分,以、鲤、鲮等杂食性鱼类为主,黄颡鱼、斑鱯、鲇等肉食性鱼类次之,草鱼、黄尾鲴等植食性鱼类最少。
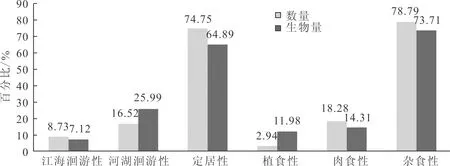
图2 北江鱼类生态类型Fig.2 The fish ecological type of Beijiang River
2.3 各季节鱼类优势种
各季节鱼类种类优势种分析表明:北江各季节鱼类优势种差异不明显。夏季鱼类优势种的种类最多。、鲤和鲮是整个北江流域的年度优势种(表1)。
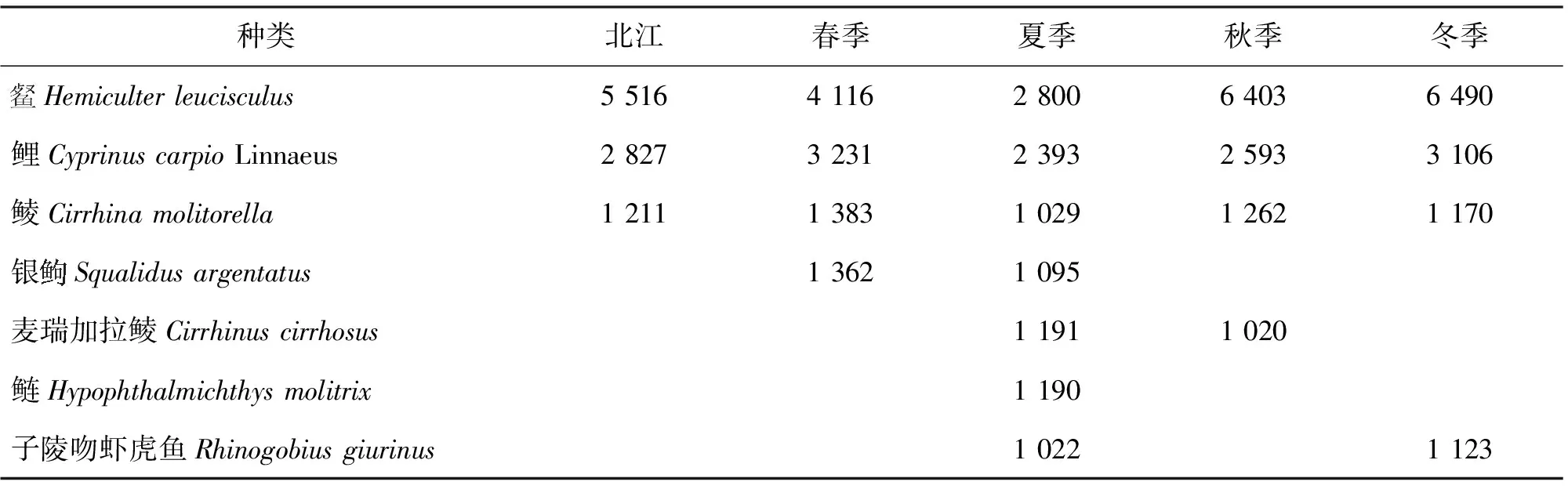
表1 北江各季节鱼类优势种(IRI>1000)Tab.1 Dominant fish species of Beijiang River in each season (IRI>1 000)
2.4 鱼类群落聚类分析
聚类分析显示,以相似性系数64.4%为标准,可将各站点鱼类群落结构分为两个类群(图3)。类群Ⅰ包括韶关站点和阳山站点,类群Ⅱ包括石角站点、连江口站点和清远站点。NMDS排序的协助强迫系数stress=0.01(stress值小于0.05),表明排序结果较高可信度(图4)。

图3 北江各站点鱼类群落结构等级聚类图Fig.3 Beijiang River fish community structure hierarchy clustering figure of each site

图4 北江各站点鱼类群落结构NMDS排序图Fig.4 Beijiang River fish community structure NMDS sorting figure of each site
2.5 北江鱼类群落丰度-生物量比较曲线
整个北江流域鱼类群落的生物量优势度曲线和丰度优势度曲线处于交叉状态,结合W值可知,北江鱼类群落结构处于中度干扰状态(图5)。

图5 北江ABC曲线Fig.5 The ABC curve of Beijiang River
2.6 北江鱼类群落与环境因子之间的典型对应分析(表2)
本研究对6项环境因子(图6)进行初选,按其对物种季节变化的解释力大小排序依次为水温(Tem)、溶氧量(DO)和盐度(Sal),最终保留这3个环境因子重新执行CCA分析,结果如图7所示。
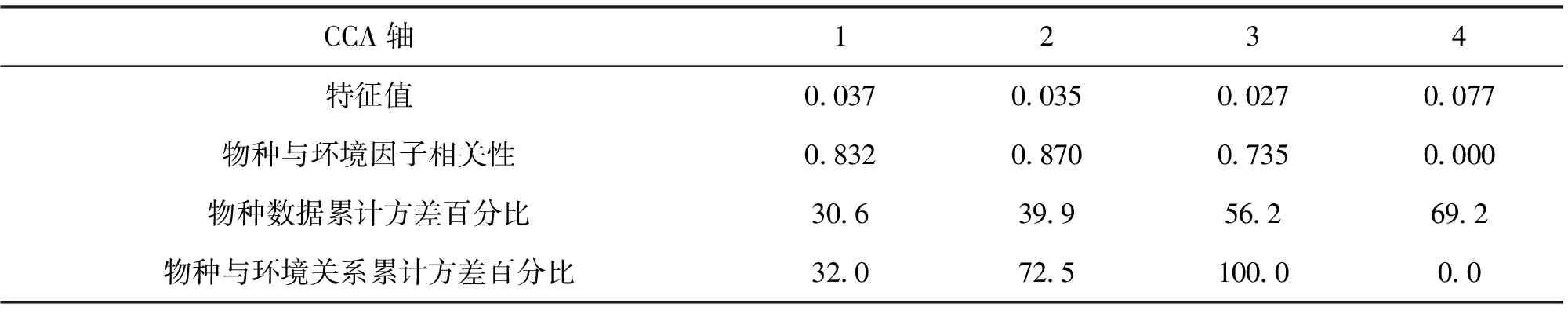
表2 北江鱼类群落结构变化与环境因子关系的CCA分析Tab.2 CCA analysis of the relationship between fish community structure variation and environmental factors in the Beijiang River
由表2可知,北江环境变量与物种季节变化的典型对应分析(CCA)的第一、第二和第三轴上物种与环境因子的相关性分别为0.832、0.870和0.735,三轴共解释了鱼类物种数据56.2%的变化。
3 讨论
3.1 鱼类种类与优势种组成分析
潘炯华[1]于20世纪80年代初对北江进行了鱼类资源调查,共采集鱼类142种,本次调查鱼类77种,明显少于上次调查种类数。这可能与调查范围、调查强度以及调查频次等有关。吕振波等[15]研究认为,过度捕捞和环境退化造成鱼类资源衰退,种群数量减少,会降低被捕获几率。因此这可能与北江近年来鱼类栖息环境发生显著恶化,种群资源量减少,使得被捕获率降低有关。
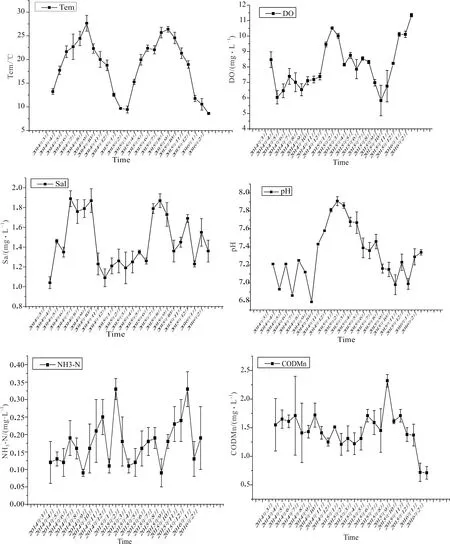
图6 北江水质参数月季变化Fig.6 The change of water quality parameters monthly in the Beijiang River
优势种是影响鱼类群落特征因素之一,其种类变化可能影响到群落种类组成、能量流动和物质循环[16]。北江各季节鱼类优势种差异不大,这与董静瑞等[16]研究结果相差明显,可能是由于内陆江段鱼类在水利大坝阻隔下,不像沿海鱼类具有较明显的季节性生殖洄游及索饵活动,使得鱼类优势种季节性差异明显。
3.2 鱼类群落聚类及稳定性分析
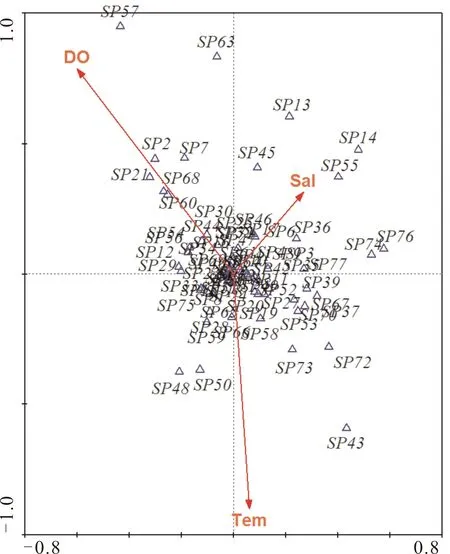
图7 北江鱼类群落结构变化与环境因子关系的CCA 排序图
鱼类群落多样性和生活环境呈正相关,鱼类群落结构主要受生境复杂性的影响,在相似生境中鱼类群落组成也非常相似,许多鱼类仅在特定生境出现[17-18]。本研究中韶关站点和阳山站点分别位于北上游及支流,两站点河流水质污染程度相对较小,鱼类栖息地结构复杂,河床有大小不等的石块堆积,岸边水草相对丰富,这些条件能为鱼类栖息、产卵等提供优良条件。此外,两站点渔获物组成相似,群落中以银、蛇等小型亚科种类相对较多,因此聚类分析聚为一组;连江口站点、清远站点和石角站点位于北江中下游,河道较深,水流较急,河床结构相对简单,小型杂鱼种类相对较少,鱼类多样相对较低,鱼类群落中多以鲤、鲫、鲢、鳙、鲮等鲤亚科、鲢亚科及野鲮亚科大中型经济鱼类占主导,在等级聚类中聚为另一组。
由ABC曲线分析可知,北江鱼类群落结构处于干扰状态。这可能是过度捕捞和生境退化导致鱼类优势种由长生命周期、高营养层次、大中型种类向短生命周期、低营养层次、中小型种类转变,鱼类群落营养级下降,高营养级捕食者对低营养级被捕食者的数量影响机制弱化,使得北江鱼类小型化和低龄化现象严重,从而导致群落稳定性和抗干扰能力下降。
3.3 鱼类群落结构及其与环境因子的关系
鱼类多样性和河流生态环境密切相关,环境因子通过影响鱼类对栖息地的偏好,塑造了多样性的地理分布格局[19-23]。CCA分析表明,水温对北江鱼类群落季节性变化影响最大。这与李捷等[24]以及帅方敏等[25]的研究结论基本一致。水温是影响鱼类关键生理、生化和生活史过程的重要环境因子,它在促使鱼类性腺发育和诱使鱼类产卵繁殖方面起着重要作用[26]。由图7可知,SP43(中华鳑鲏)、SP48(小鰁)和SP50(粗唇鮠)等种类相对丰度与水温(Tem)呈正相关性,SP2(鲤)、SP7(鲫)和SP21(蛇)则更趋向于低温季节出现。这与鱼类生活史受温度调控有关。每月渔获物调查发现,北江一些鲤亚科(如:鲤、鲫)和亚科(如:蛇)繁殖活动相对较早,每年3月初,水温达到15 ℃即可开始产卵,繁殖季节鱼类群聚的高度集中现象使得在水温较低季节被捕获机率增大。而一些小型鲤科鱼类(如:中华鰟鮍、小鰁)和部分鲿科鱼类(如:粗唇鮠)在每年5月中下旬水温超过20 ℃才能达到其产卵条件,因此其相对丰度在CCA分析中显示与水温呈正相关。
溶氧量(DO)和盐度(Sal)也是影响北江鱼类群落结构季节性变化的重要环境因子。北江溶氧量均在6 mg/L以上(图6),不会直接对鱼类活动造成不良影响。因此,溶氧量可能是间接反映河流水温和流速综合变化情况。沈忱等[27]研究表明,随着河流水温增加,水体溶解氧量会随之降低。结合北江溶氧量监测数据可知,每年冬季北江溶氧量最高。SP57(鲻)、SP2(鲤)和SP21(蛇)等种类相对丰度与溶氧量呈正相关,鲻鱼属咸淡水洄游性鱼类,每年鱼苗发生季节为1-3月,鲤在北江每年2-3月开始分批产卵,蛇在北江每年3月初水温达到12 ℃即可开始繁殖,这就使得这些鱼在该时期捕获率增加,表现出相对丰度与水体溶氧呈正相关趋势。盐度(Sal)季节性变化表现出夏季较高的趋势,本研究认为盐度变化可能是由于夏季水温较高,水体蒸发量较大,导致水体盐度上升。由图7可知,SP14(黄颡鱼)、SP55(瓦氏黄颡鱼)和SP13(赤眼鳟)趋向于盐度较高的季节出现。在调查期间发现,黄颡鱼、瓦氏黄颡鱼等一些鲿科鱼类及赤眼鳟等部分雅罗鱼亚科鱼类,夏秋季节摄食代谢活动极为旺盛,在该季节渔获均占有较高比例。综上所述,水温是对北江鱼类群落结构季节性变化影响最大的水环境因子,本研究推测,其它水环境因子对北江鱼类群落结构的季节性影响可能是由水温季节性变化直接或间接导致,这有待于进一步探究。
致谢:蒙美国阿肯色大学小石城分校(University of ArKansas at Little Rock)汤伏生(Fusheng Tang)副教授对中英文摘要进行润色修改,特致谢意!
[1]潘炯华.珠江水系北江渔业资源[M].广州:广东科学技术出版社,1987.
[2]李桂峰.广东淡水鱼类资源调查与研究[M].北京:科学出版社,2013.
[3]张觉民,何志辉.内陆水域渔业自然资源调查手册[M].北京:中国农业出版社,1991.
[4]潘炯华.广东淡水鱼类志[M].广州:广东科学技术出版社,1991.
[5]Pinkas L,Oliphant M S,Iverson I L K.Food habits of albacore,bluefin tuna,andbonito in California waters[J].Fish B-Noaa,1971,152:1-105.
[6]Clarke K R,Warwick R M.Changes in marine communities:an approach to statistical analysis and interpretation (2nd edition)[M].PRIMPER-E:Plymouth,2001.
[7]Warwick R M.A new method for detecting pollution effects on marine macrobenthic communities[J].Mar Biol,1986,92:557-562.
[8]刘春池,高 欣,林鹏程,等.葛洲坝水库鱼类群落结构特征研究[J].长江流域资源与环境,2012,21 (7):843-849.
[9]孙莎莎,唐文乔,郭弘艺,等.靖江沿岸秋季鱼类群聚的组成特点及其丰度生物量变[J].生物多样性,2013,21(6):688-698.
[10]何勇凤,李昊成,王旭歌,等.长湖鱼类群落结构的时空变化[J].长江流域资源与环境,2016,25(2):265-273.
[11]Leps J,Smilauer P.Multivariate analysis of ecological data using CANOCO[M].London:Cambridge Univ.Press,2003.
[12]刘 飞,娄必云,刘焕章,等.赤水河赤水市江段鱼类群落结构季节变化特征[J].淡水渔业,2015,45(1):35-41.
[13]McCune B,Grace J B.Analysis of Ecological Communities[M].MjM SoftwareDesign,USA,2002.
[14]Ter B C J F,Smilauer P.CANOCO Reference Manual and User′s Guide:Software for Ordination Version 5[M].New York:Microcomputer Power Ithaca,2002.
[15]吕振波,李 凡,王 波,等.黄海山东海域春、秋季鱼类群落结构[J].水产学报,2011,35(5):692-699.
[16]董静瑞,胡成业,水玉跃,等.温州南部沿岸海域鱼类群落特征及其与环境因子的关系.中国水产科学,2017,24(2):209-219.
[17]Ornellas A B,Coutinho R.Spatial and temporal patterns of distribution and abundance of a tropical fish assemblage in a seasonal Sargassum bed,Cabo Frio Island,Brazil [J].Fish Biol,1998,53:198-208.
[18]李 捷,李新辉,贾晓平,等.西江鱼类群落多样性及其演变[J].中国水产科学,2010,(2):298-311.
[19]王雪辉,邱永松,杜飞雁,等.北部湾鱼类群落格局及其与环境因子的关系[J].水产学报,2010,(10):1579-1586.
[20]He Y F,Wang J W,Lek S,et al.Structure of endemic fish assemblages in the upper Yangtze River Basin[J].River Res Appl,2011,27(1):59-75.
[21]Trumpickas J,Mandrak N E,Ricciardi A.Nearshore fish assemblages associatedwith introduced predatory fishes in lakes[J].Aquat Conserv,2011,21:338-347.
[22]王 生,段幸斌,陈文静,等.鄱阳湖湖口鱼类资源现状调查[J].淡水渔业,2016,46( 6) :50-55.
[23]林小植,李冬梅,刘焕章,等.广东韩江潮州江段鱼类多样性及季节变化[J].生物多样性,2016,24(2):185-194.
[24]李 捷,李新辉,贾晓平,等.连江鱼类群落多样性及其与环境因子的关系[J].生态学报,2012,32(18):5795-5805.
[25]帅方敏,李新辉,刘乾甫,等.珠江水系鱼类群落多样性空间分布格局[J].生态学报,2017,37(9):3182-3192.
[26]殷名称.鱼类生态学[M].北京:中国农业出版社,1993:64-88.
[27]沈 忱,吕平毓,冯顺新,等.向家坝水库蓄水对下游江段溶解氧饱和度影响研究[J].淡水渔业,2014,44(6) :31-36.
附件1:
FishcommunitystructureanditsrelationshipswithenvironmentalfactorsintheBeijiangRiver
GAO Tian-yang,XIE Di,PENG Ning-dong,LI Chao,ZHANG Shao-ping,LUO Jin-zhen,WANG Chun-hui,ZHAO Jun
(KeyLaboratoryforHealthyandSafeAquacultureofGuangdongProvince;GuangdongProvinceEngineeringTechnologyResearchCenterforEnvironmentally-friendlyAquaculture;GuangzhouKeyLaboratoryofSubtropicalBiodiversityandBiomonitoring;SchoolofLifeScience,SouthChinaNormalUniversity,Guangzhou510631,China)
To fully understand the community structure of fishes,we conducted a montly field-survey of fish resources in Beijing River from March 2014 to February 2016.During this survey,77 species of fishes were captured,belonging to 8 orders,16 families and 56 genera.Among these fishes,there were 44 species of Cypriniformes,which account for 57.1% of all fish species in this survey;10 species of Perciformes,which account for 12.9% of all fish species in this survey.8 species of Siluriformes,which account for 10.3% of all fish species in this survey.The rest Cypriniformes,Synbranc,Clupeiformes,Salmoniformes and Mugiliformes species were less than 5.Through the total 77 species of fishes onlyHemiculterleucisculus,CyprinuscarpioandCirrhinamolitorellawere the annual dominant species of the Beijiang River.Typical correspondence analysis (CCA) revealed that water temperature (Tem),dissolved oxygen (DO) and salinity (Sal) were the main environmental factors on how the seasonal variations affect the Beijiang River fish community structure.
Beijiang River;fish community structure;seasonal variation;typical corresponding analysis
2017-06-22;
2017-08-24
国家公益性行业(农业)科研专项(201303048);国家科技基础条件平台工作重点项目(2005DKA21402)
高天扬(1991- ),男,硕士,主要从事水产种质资源研究。E-mail:gaotianyang163@163.com
赵 俊。E-mail:zhaojun@scnu.edu.cn
S932.4
A
1000-6907-(2017)06-0047-09

附件1 北江调查鱼类名录(依IRI指数排序)Appendix1list of Beijiang Riverfishes (ranked according to IRI)

编号种类数量百分比/%重量百分比/%相对重要性指数(IRI)重要性级别SP41花鰶Clupanodonthrissa0180102759少见种SP42半Hemiculterellasauvagei0190012028少见种SP43中华鳑鲏Rhodeussinensis0170011840少见种SP44间Hemibarbusmedius0090081737少见种SP45南方白甲鱼Varicorhinus0040131713少见种SP46翘嘴鲌Culteralburnus0040091301少见种SP47马口鱼Opsariichthysbidens0100021151少见种SP48小鰁Sarcocheilichthysparvus011<0011096少见种SP49海南华鳊Sinibramamelrosei0050061066少见种SP50粗唇鮠Leiocassiscrassilabris004006953稀有种SP51丁鱥Tincatinca001008950稀有种SP52七丝鲚CoiliagrayiRichardson007002914稀有种SP53南方拟Pseudohemiculterdispar005004872稀有种SP54唇Hemibarbuslabeo005002707稀有种SP55瓦氏黄颡鱼Pelteobagrusvachelli006001704稀有种SP56颌针鱼Tylosurusstrongylurus006001702稀有种SP57鲻鱼MugilcephalusLinnaeus001006690稀有种SP58尖头塘鳢Eleotrisoxycephala006001673稀有种SP59下口鲇Hypostomusplecostomus002005653稀有种SP60鳡Elopichthysbambusa<001004448稀有种SP61棒花鱼Abbottinarivularis004<001425稀有种SP62条纹鲮脂鲤PlineatusProchiloduslineatus002003414稀有种SP63白肌银鱼Leucosomachinensis003<001378稀有种SP64间下鱵Hyporhamphusintermedius002<001266稀有种SP65花鲈Lateolabraxjaponicus001002242稀有种SP66长吻鮠Leiocassislongirostris<001002185稀有种SP67银鲴Xenocyprisargentea<001001102稀有种SP68东方墨头鱼Garraorientalis001<001101稀有种SP69大眼华鳊Sinibramamacrops<001<001086稀有种SP70条纹小鲃Puntiussemifasciolatu<001<001072稀有种SP71花鳗鲡Anguillamarmorata<001<001016稀有种SP72珠江长臀鮠Cranoglanisbouderiusbouderius<001<001012稀有种SP73中华花鳅Cobitissinensis<001<001006稀有种SP74中国少鳞鳜Sinipercawhiteheadi<001<001002稀有种SP75波纹鳜Sinipercaundulata<001<001002稀有种SP76纹唇鱼Osteochilussalsburyi<001<001001稀有种SP77卷口鱼Ptychidiojordani<001<001001稀有种
