冬季供给垫草对扎龙保护区笼养丹顶鹤的作用和影响
2017-11-17杨志宏邹红菲邵淑丽郭立业林春山高忠燕
杨志宏 邹红菲 邵淑丽 郭立业 林春山 高忠燕
(1.齐齐哈尔大学生命科学与农林学院,齐齐哈尔,161006,2.东北林业大学野生动物资源学院,哈尔滨,150040,3.黑龙江扎龙国家级自然保护区,齐齐哈尔,161003)
冬季供给垫草对扎龙保护区笼养丹顶鹤的作用和影响
杨志宏1邹红菲2邵淑丽1郭立业3林春山3高忠燕3
(1.齐齐哈尔大学生命科学与农林学院,齐齐哈尔,161006,2.东北林业大学野生动物资源学院,哈尔滨,150040,3.黑龙江扎龙国家级自然保护区,齐齐哈尔,161003)
供给垫草是冬季扎龙保护区针对笼养丹顶鹤采取的重要保护对策之一。为完善保护对策,本文探讨了冬季供给垫草对笼养丹顶鹤的作用和影响。考虑笼养鹤的安全越冬,本文仅对保护区冬初开始供给和冬末停止供给垫草两个时间节点内有(Y)无(W)垫草组丹顶鹤的能量收支、行为和体征变化进行了比较。结果发现,冬初供给垫草后W组丹顶鹤摄食玉米的量、摄入总能量和摄入玉米的能量均显著高于Y组(P﹤0.001),摄水量和饮水量却显著低于Y组(P﹤0.05),Y组日间排泄次数(笼舍内垫草以外的地面)显著多于W组,白天摄食、静息、警戒和游走的行为时间分配组间差异显著(P﹤0.05)和夜晚Y组丹顶鹤(5/7)趴卧静息于垫草之上;冬末停止供给垫草后W组丹顶鹤摄入玉米的量显著高于Y组(P﹤0.05),但摄入总能量、排泄次数、白天行为时间分配和夜晚静息位点均无组间显著差异;冬季丹顶鹤腹部覆羽的污损与其夜晚趴卧静息于垫草之上有关。本文认为,冬季最冷月份夜晚垫草可以确保笼养丹顶鹤的跗蹠和爪不被冻伤、有利于保温和节省能耗,保护区冬初开始供给垫草时间的选择合理(冬初的冷适应有利于提高丹顶鹤御寒的生理机能、此时气温已与野生丹顶鹤越冬地盐城冬季最低气温接近和W组丹顶鹤未被冻伤),而冬末停止供给垫草时间节点可以适当提前(依据实验结果),缩短供给垫草总时长或增加更换垫草频次均可减少鹤腹部覆羽的污损。总之,从动物福利角度和确保笼养丹顶鹤安全越冬考虑应该供给垫草,同时供给垫草不仅要关注鹤的安全越冬,还应考虑鹤的健康。
丹顶鹤; 扎龙保护区; 垫草
丹顶鹤(Grusjaponensis)属鹤形目(Gruiformes)鹤科(Gruidae)的大型渉禽,具有体表色彩光鲜亮洁和喙、颈、腿3长的特点,为我国Ⅰ级保护鸟类,IUCN(2012)和中国脊椎动物红色名录将其列为濒危(Endangered,E)物种[1-2]。扎龙国家级自然保护区(N 46°52′~47°32′,E 123°47′~123°37′)是中国北方同纬度地区中保留最完整、最原始、最开阔的湿地生态系统。作为丹顶鹤的主要繁殖地之一,每年春季会有包括丹顶鹤和白枕鹤在内的7种鹤类在此繁殖或停歇[3-4]。保护区在丹顶鹤的就地保护和种群扩增等方面已经做了大量工作。至今,每年冬季有300余只(其中大部分为笼养、部分野化放归和少数野生)丹顶鹤留居保护区内越冬,其就地保护一直备受关注。笼养丹顶鹤无法自由选择栖息环境,给予其科学合理的保护意义重大[5-6]。每年冬季,扎龙保护区都会供给笼养鹤垫草以确保其安全越冬。
扎龙保护区属大陆性季风气候区。冬季漫长,气低温、昼夜温差大和最低气温(夜间)可超过-30℃,昼短夜长(光照)和最短时接近8∶16 h(昼∶夜),且经常刮强劲的西北冷风等。冬初开始,气温逐渐下降(期间会有小幅回暖天气),光照时间逐日缩短直至“冬至”。冬至前后气温将降至全年最低。冬至开始,气温逐渐上升(期间会有小幅降温天气),光照时间逐日延长直至“夏至”。留居丹顶鹤需要面对冬日气温低、昼夜温差大和光照昼短夜长的生存压力(夜晚的生存压力远大于白天)。能站不卧是丹顶鹤的习性之一。丹顶鹤长、厚且浓密的体表覆羽足以抵御冬季严寒,但其跗蹠和爪表面无覆羽、终日暴露在外且爪与地面直接接触,所以跗蹠和爪不被冻伤是其成功越冬的关键。
扎龙保护区冬季室外笼养丹顶鹤是否需要供给垫草,供给垫草后它们会有哪些相应行为发生,垫草能否为其带来能量收支收益,保护区供给垫草时间节点的选择及其对丹顶鹤的作用和影响等问题都需要进行探讨和论证。扎龙保护区冬季丹顶鹤全部生存于室外,考虑到冬季最冷月份无垫草可能对笼养丹顶鹤造成伤害,所以本文仅对保护区冬初开始供给垫草和冬末停止供给垫草两时段有无垫草组笼养丹顶鹤的行为和能量收支进行比较(实验组仅在实验期间无垫草),并结合供给垫草时的气候特点来探讨垫草对笼养鹤的作用和影响并进行利弊权衡,以期进一步完善这一就地保护对策。
1 材料与方法
1.1 动物来源、分组、饲养、处理
丹顶鹤越冬,动物园常会选择暖室或冷室这2种方式,然而扎龙保护区内的丹顶鹤(包括笼养和散养)全部生存于室外。以冬季保护区室外单笼(6 m×4 m×4 m/只)饲养的14只成年丹顶鹤(8♂、6♀)个体为研究对象,分为无垫草(W;4♂、3♀)和供给垫草(Y;4♂、3♀)2组,实验分别于保护区冬初开始向笼舍内供给垫草后一周和冬末停止供给垫草前一周的两时段内进行,共14 d。冬季最冷月份时段,为确保丹顶鹤安全而没有进行有无垫草的对比实验。分组方法:本文采用拟实验室方法对丹顶鹤进行分组。分组时考虑(丹顶鹤)更换笼舍可能会对实验结果产生影响,本文采取就近、有利、减少干扰和误差的原则将两排笼舍丹顶鹤分为有无垫草2组。丹顶鹤体重存在个体差异,初体重的组间显著差异应该会导致终体重和能量收支各参数的实测值与真实值之间出现误差并影响结果,故通过统计学方法分别对冬初和冬末丹顶鹤初体重组间是(以初体重为协变量进行组间协方差分析来消除初体重组间差异对实验结果的影响)否(t-检验)差异显著来进行相关参数的相应处理。实验期间,供给食物(鲫鱼和玉米)及饲喂方法(鲫鱼定时8:30~15:00足量供给和干玉米粒全天足量供给)均相同。
1.2 实验期间气候
保护区供给垫草起始时间为2014年11月29日开始至2015年3月5日结束。2014年11月29日开始至12月6日结束共7 d,日均最高(白天)气温为(-10.00±3.37)℃,日均最低(夜晚)气温为(-17.14±2.73)℃,光照时间平均为8 h 45 min(日出时间从07:08延后至07:14和日落时间从15:57提前至15:55),常刮3~4级的北风(或偏北风)。2014年3月5日开始至3月12日结束共7 d,日均最高气温为(-2.86 ± 2.91)℃,日均最低气温为(-13.51±3.41)℃,光照时间平均为11 h 28 min(日出时间从06:17提前至06:07和日落时间从17:36延后至17:43),前4 d伴有3~4级北风(或偏北风)和后3 d出现2~3级南风。2014年冬季最冷月份1月平均气温-19.2℃,日均最低气温-24℃,最冷日最低气温为-29℃。
1.3 行为差异
利用监控录像观测。白天对丹顶鹤行为及其时间分配进行组间比较,方法见杨志宏等[6]。夜晚采用定时记录方式来确定丹顶鹤栖息位点的选择,即每夜定时(考虑干扰因素,分别选择在20:00~21:00、23:00~00:00和3:00~4:00,3个时间段内)监测(60 min/次)3次(仅记录丹顶鹤处于静息状态时的栖息位点),3次中同1只丹顶鹤于相同位点静息达到2次就认为其存在栖息位点选择。
1.4 体重和体温
采用电子秤(TCS100,浙江)称量体重。称量方法为[6]:A=B-C(其中,A为丹顶鹤的体重;B为饲养员与丹顶鹤体重之和;C为饲养员体重)。分别于两段实验的实验前(初体重)和实验结束(终体重)的相同时间称量丹顶鹤体重,共称量体重4次。称量体重同时测量丹顶鹤体温,共测量体温4次。用电子体温计插入泄殖腔约3 cm测量丹顶鹤体核温度,每次测温探头在泄殖腔内静置约20 s和数值稳定时读数(精确到0.10℃)。
1.5 能量收支
用氧弹热量计(GR-3500型,广东)测定食物热值。排出水热能散失计算中丹顶鹤的体温均为平均体温,即实验前后2次体温的平均值。摄入水水温是以初水温(饲喂时刻测定水温)和终水温(冰水混合时水温记为0℃)的平均值计算。能量收支计算公式如下:
干物质(kg)=物质总重量(kg)-物质中水质量(kg)
摄食量(kg/d)=投食量(kg/d)-剩余食物量(kg/d)
摄入能(kJ/d)=摄入干食物质量(kg/d)×干食物热值(kJ/kg)
摄水量(kg/d) =饮水容器和水量(初重-末重)(kg/d)+摄入食物中的水量(kg/d)
排出水热能散失(kJ/d)=水的比热值(kJ/kg℃) ×摄水量(kg/d)×(鸟体温-摄入水温)(℃)
1.6 排泄次数
实验开始,每天累计计数笼舍内垫草外地面的排泄次数(每次计数后均随即清除粪便,避免粪便重合,使累计计数准确)5次(昼4和夜1),24 h内5次计数总和记为日排泄次数/只。丹顶鹤的排泄物多为稀粥状,粪便排泄到垫草之上后会与垫草混合、粘连并冻结于垫草之中而无法准确计数,所以垫草之上的排泄次数并未累计计入该组每只丹顶鹤的日排泄次数。
1.7 统计分析
利用SPSS 17.0软件包进行相关统计分析。初体重、体温、行为时间分配、排泄次数、终体重和能量收支(初体重组间差异不显著)及同组组内能量收支的比较均采用双尾t检验分析。初体重组间差异显著,终体重和能量收支的比较采用以初体重为协变量进行协方差分析(ANCOVA)。文中数据均以平均值±标准误(Mean ±SE)表示,P﹤0.05即认为差异显著。
2 结果
2.1 体重、体温和腹部覆羽
冬初开始供给垫草时,丹顶鹤W组(11.04±0.58)kg和Y组(10.42±0.66)kg初体重组间差异显著(t=2.629,df=12,P=0.042)。7 d后,W组(11.52±0.76) kg和Y组(10.76±0.42) kg终体重增加分别为4.35%和3.26%,组内体重增加均显著(t=-3.398,df=6,P=0.032;t=-2.898,df=6,P=0.028)。当初体重校正至10.72 kg时,终体重组间比较差异不显著(F(1,11)=0.997,P=0.769)。7 d后,丹顶鹤W组(42.88±0.66)℃和Y组(42.54±0.82)℃体温组间差异不显著(t=0.068,df=12,P=0.838)。实验结束时观察可见供给垫草组丹顶鹤的腹部覆羽已有污渍(5/7),而未添加垫草组丹顶鹤腹部覆羽均无污渍。
冬末停止供给垫草时,丹顶鹤W组(10.04±0.66) kg和Y组(10.07±0.43) kg初体重组间差异不显著(t=-0.072,df=12,P=0.945)。7 d后,W组(9.70±0.66) kg和Y组(9.94±0.39) kg 终体重降低分别为3.34%和1.34%,组内体重降低均不显著(t=2.989,df=6,P=0.164;t=1.546,df=6,P=0.220)。终体重组间比较差异不显著(t=-0.308,df=12,P=0.769)。7 d后,丹顶鹤W组(42.54±0.68)℃和Y组(42.36±0.44)℃体温组间差异不显著(t=0.089,df=12,P=0.985)。实验结束时丹顶鹤组间腹部覆羽污损无显著差异。
2.2 能量收支
冬初丹顶鹤初体重校正至10.72 kg时,摄食玉米的量W组显著高于Y组117.88%(F(1,11)=43.560,P=0.000)。摄水量和饮水量W组显著低于Y组8.48%和22.53%(F(1,11)=16.545,P=0.046和F(1,11)=35.986,P=0.008),摄食鲫鱼的量和摄入鲫鱼中的水量组间差异不显著(F(1,11)=0.404,P=0.537和F(1,11)=0.721,P=0.412)(图1)。摄入总能量和摄入玉米的能量W组显著高于Y组18.92%和117.86%(F(1,11)=21.426,P=0.006;F(1,11)=43.350,P=0.001)。W组和Y组摄入玉米的能量分别占其各自摄入总能量的31.36%和17.12%。W组和Y组摄入鲫鱼的能量和排出水热能散失组间差异均不显著(F(1,11)=0.404,P=0.537和F(1,11)=0.854,P=0.374)(图2)。
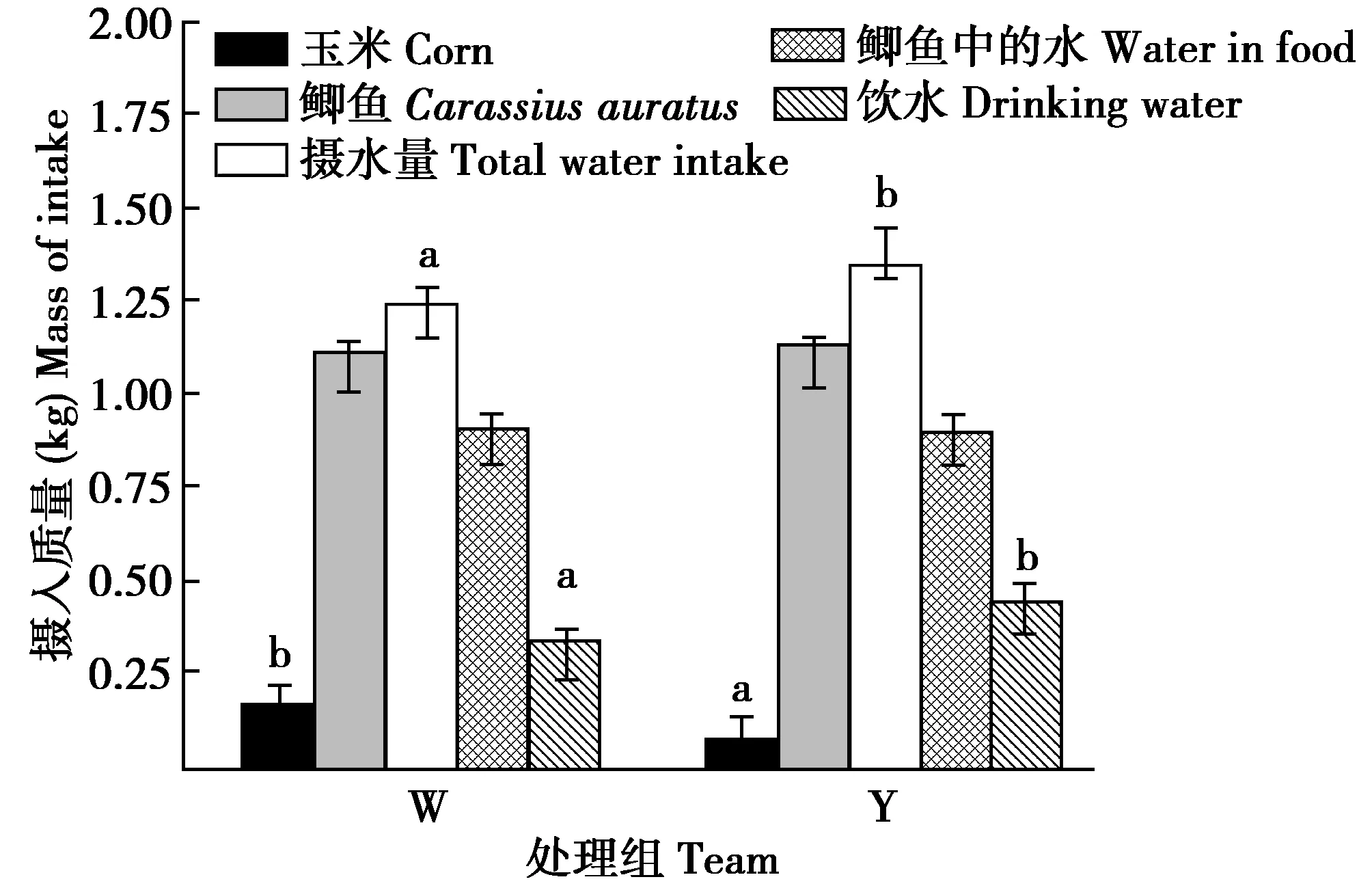
图1 冬初供给垫草时有无垫草组丹顶鹤摄入食物和 水需求的组间比较Fig.1 Comparison of food and water intake in different captive red crowned crane early winter 注:初体重组间差异影响已经消除;W为未供给垫草,Y为供给垫草 Note:The effect of initial body mass difference between groups has been eliminated,W denote no mate grass and Y denote supply mat grass
冬末,W组摄食玉米的量显著高于Y组44.62%(t=4.277,df=12,P=0.044)。摄食鲫鱼的量、摄水量、摄入鲫鱼中的水量和饮水量组间差异均不显著(t=-0.016,df=12,P=0.901;t=0.058,df=12,P=0.811;t=-0.032,df=12,P=0.802;t=0.321,df=12,P=0.664)(图3)。W组和Y组摄入玉米中的能量分别占各自总摄入能量的21.81%和15.87%,组间差异显著(t=4.288,df=12,P=0.036)。摄入总能量、摄入鲫鱼的能量和排出水热能散失组间差异均不显著(t=1.562,df=12,P=0.217;t=1.664,df=12,P=0.245;t=0.058,df=12,P=0.811)(图4)。
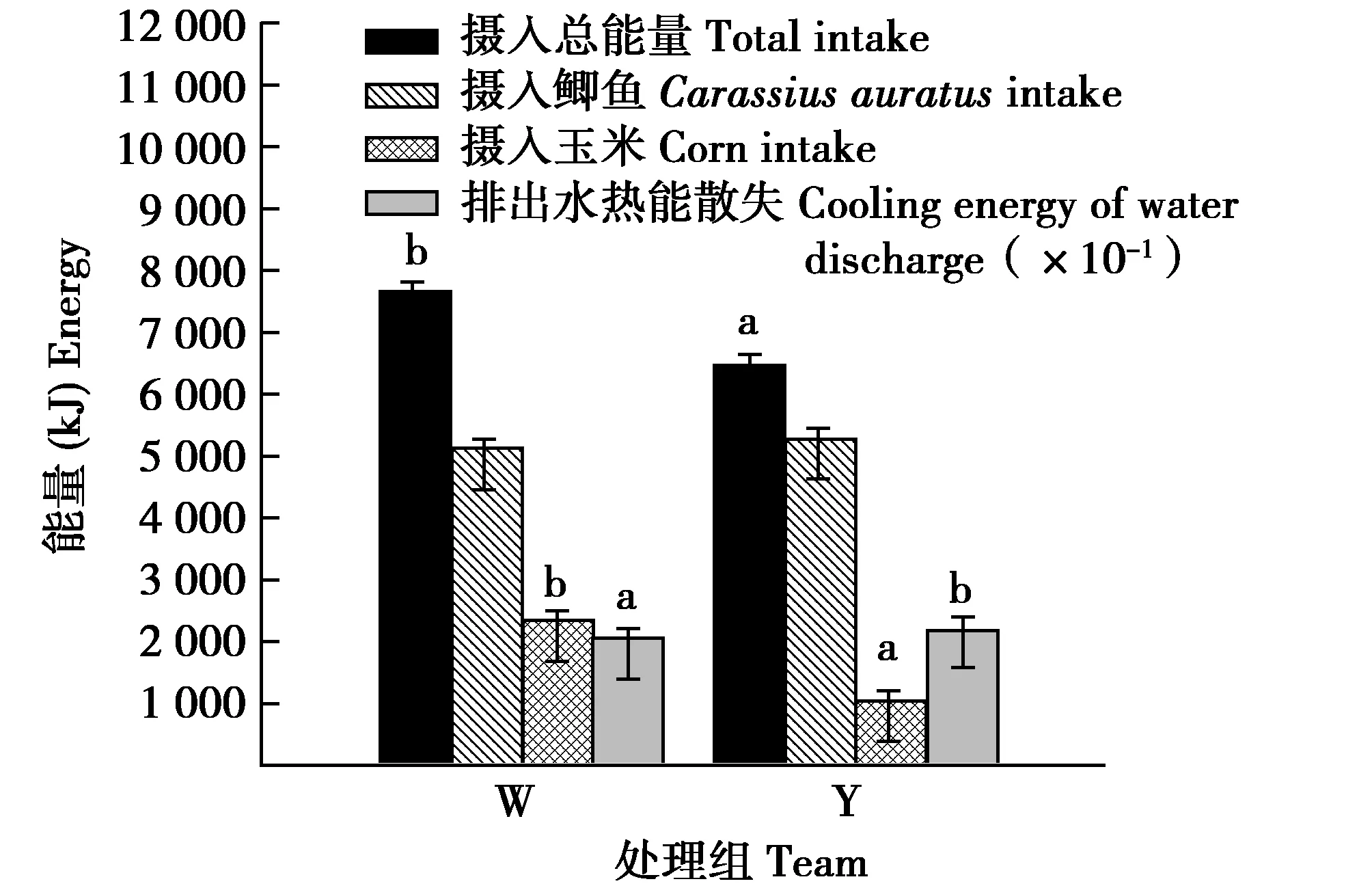
图2 冬初供给垫草时有无垫草丹顶鹤能量收支的组间比较Fig.2 Comparison of energy budget in different captive red crowned crane early winter 注:初体重组间差异影响已经消除;W为未供给垫草,Y为供给垫草 Note:The effect of initial body mass difference between groups has been eliminated,W denote no mate grass and Y denote supply mat grass
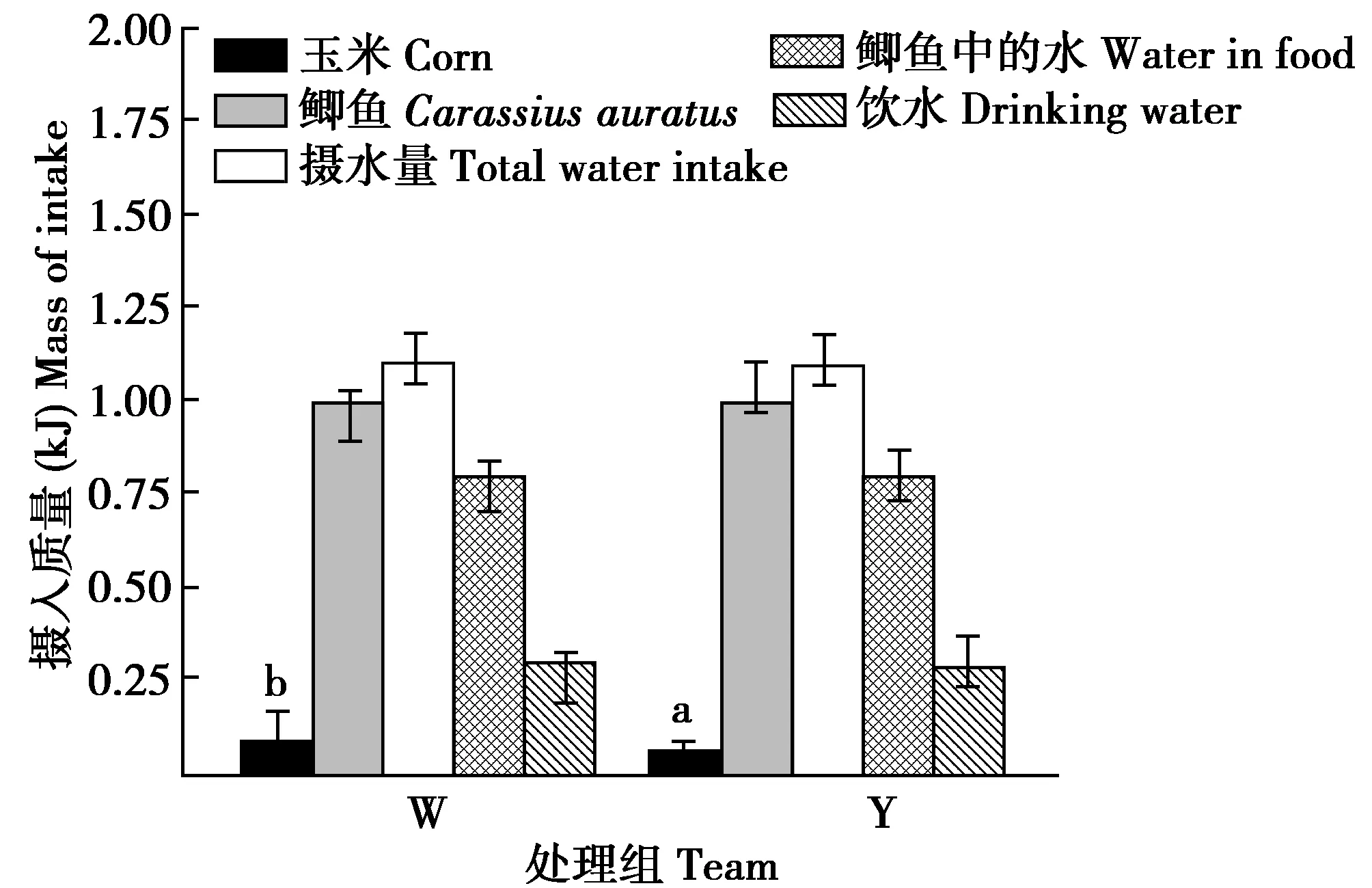
图3 冬末停止供给垫草时有无垫草丹顶鹤摄入食物 和水质量的组间比较Fig.3 Comparison of food and water intake in different captive red crowned crane late winter 注:W为未供给垫草,Y为供给垫草 Note:W denote no mat grass and Y denote supply mat grass
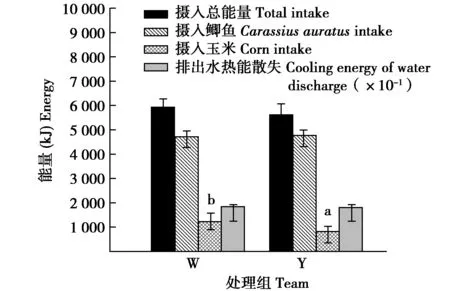
图4 冬末停止供给垫草时有无垫草丹顶鹤能量 收支的组间比较Fig.4 Comparison of energy budget in different captive red crowned crane late winter 注:W为未供给垫草,Y为供给垫草 Note:W denote no mat grass and Y denote supply mat grass
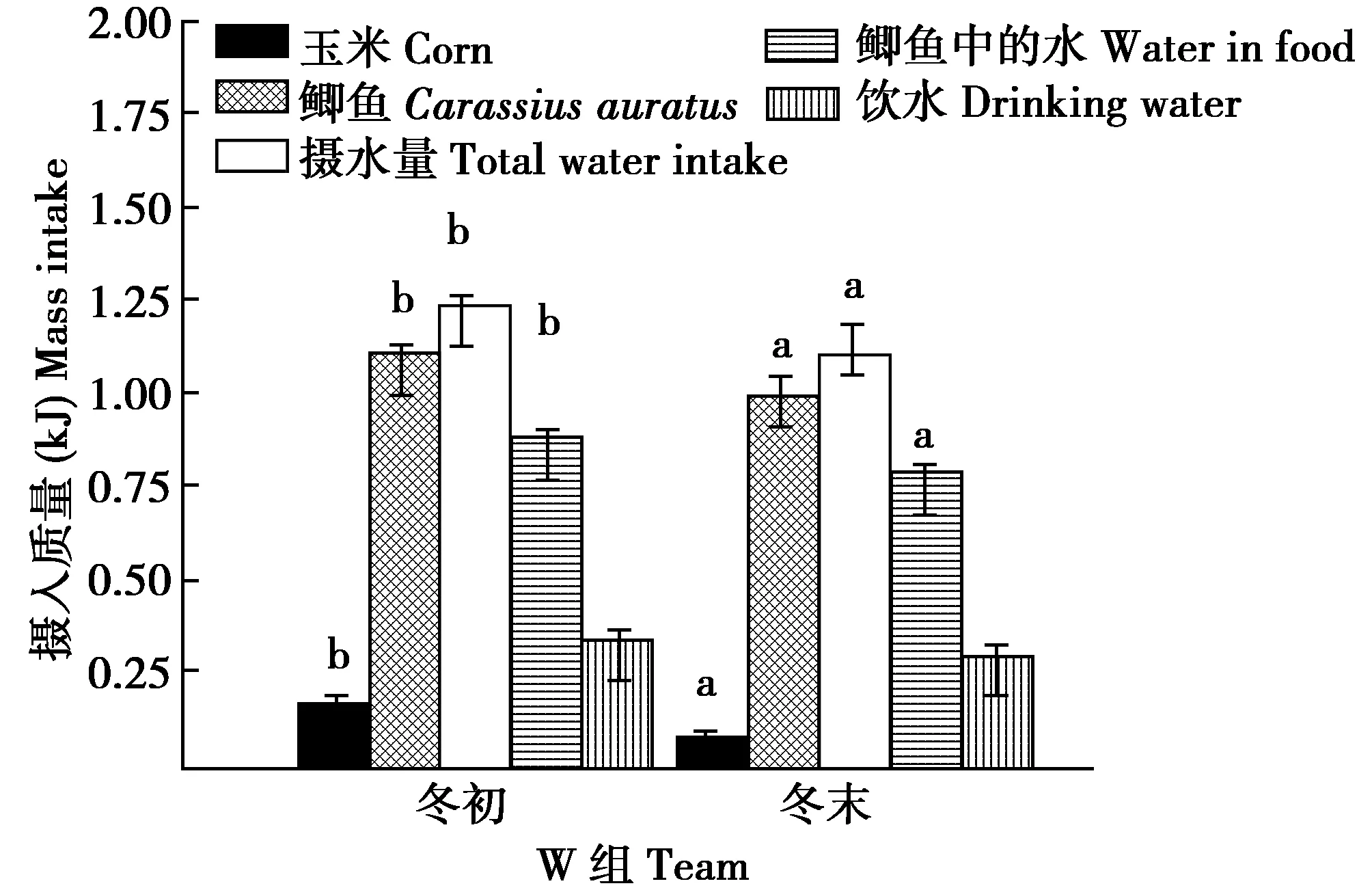
图5 W组丹顶鹤冬初和冬末摄食(水)量的组内比较Fig.5 Comparison of food and water intake within group W captive red crowned crane between early and late winter
W组,冬末比冬初摄食玉米的量、鲫鱼的量、摄水量和摄入鲫鱼中的水量均显著减少(t=6.584,df=12,P=0.028;t=6.985,df=12,P=0.024;t=5.566,df=12,P=0.036;t=6.558,df=12,P=0.022),饮水量减少但不显著(t=4.443,df=12,P=0.052)(图5)。冬末比冬初摄入总能量、鲫鱼的能量、玉米的能量和排出水热能散失的能量均显著减少(t=8.868,df=12,P=0.012;t=6.668,df=12,P=0.022;t=6.524,df=12,P=0.026;t=7.732,df=12,P=0.012)(图6)。
Y组,冬末比冬初摄食玉米的量、鲫鱼的量、摄水量、摄入鲫鱼中的水量和饮水量均显著减少(t=5.558,df=12,P=0.033;t=5.525,df=12,P=0.036;t=6.033,df=12,P=0.028;t=5.518,df=12,P=0.034;t=8.323,df=12,P=0.006)(图7)。冬末比冬初摄入总能量、鲫鱼的能量、玉米的能量和排出水热能散失的能量均显著减少(t=6.665,df=12,P=0.022;t=5.522,df=12,P=0.036;t=5.566,df=12,P=0.032;t=8.989,df=12,P=0.004)(图8)。
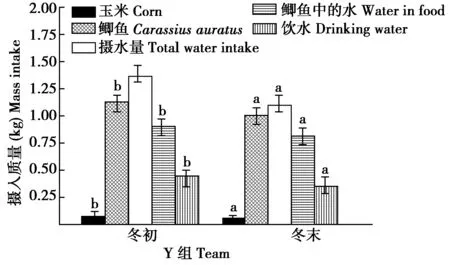
图7 Y组丹顶鹤冬初和冬末摄食(水)量的组内比较Fig.7 Comparison of food and water intake within group Y captive red crowned crane between early and late winter
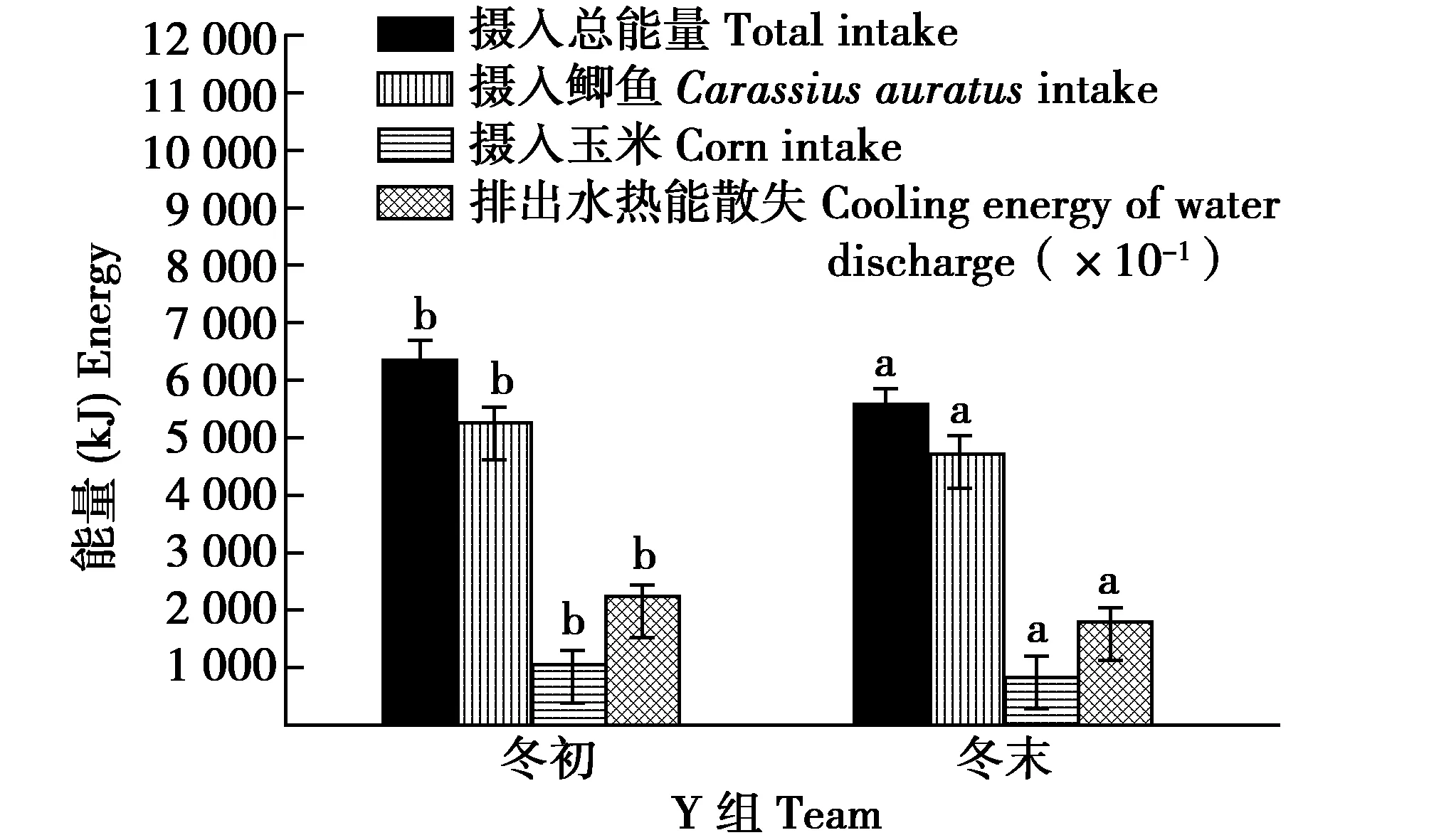
图8 Y组丹顶鹤冬初和冬末能量收支的组内比较Fig.8 Comparison of energy budget within group Y captive red crowned crane between early and late winter
2.3 排泄次数
垫草中的排泄除外,冬初丹顶鹤的排泄次数W组显著多于Y组70.90%(t=9.932,df=12,P=0.000)和冬末W组高于Y组15.26%但不显著(t=2.433,df=12,P=0.068)(表1)。
表1 有无垫草组丹顶鹤排泄次数的比较
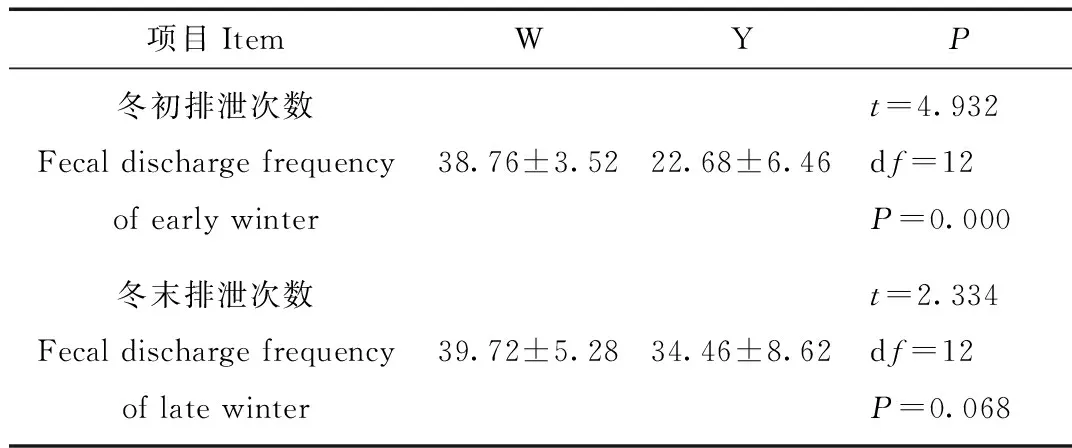
Tab.1 Comparison of discharge in different captive red crowned crane both early and late winter
注:不包含垫草之中的排泄次数;W为未添加垫草组和Y为添加垫草组;数值为平均值±标准误
Note:The fecal discharge frequency in mat grass was ignored.W denote no mate grass and Y denote supply mat grass.Date are presented as Mean±SE
2.4 行为谱与时间分配
冬初初次供给垫草时笼养丹顶鹤(大部分)随即会围绕垫草出现一系列舞蹈行为,即半伸翅膀、啄草-跳跃-高抛并用喙空中接草,或围着垫草一边扇动翅膀一边往返奔跑跳跃,或半伸展翅膀、长颈的伸缩伴随着身体的起伏,此异常兴奋的行为表现会一直持续数分钟之久。冬初,丹顶鹤白天行为时间分配Y组比W组警戒和游走显著增加63.47%和46.90%(t=9.986,df=12,P=0.006和t=5.988,df=12,P=0.034),摄食和静息显著减少24.48%和15.33%(t=10.322,df=12,P=0.004和t=6.658,df=12,P=0.032),理羽及其他组间差异不显著(t=0.633,df=12,P=0.564)(图9)。丹顶鹤夜晚以静息(藏头于背部)为主,Y组鹤选择趴卧(5/7)或单足站立(2/7)于垫草之上,W组鹤单足站立静息的位点无规律。
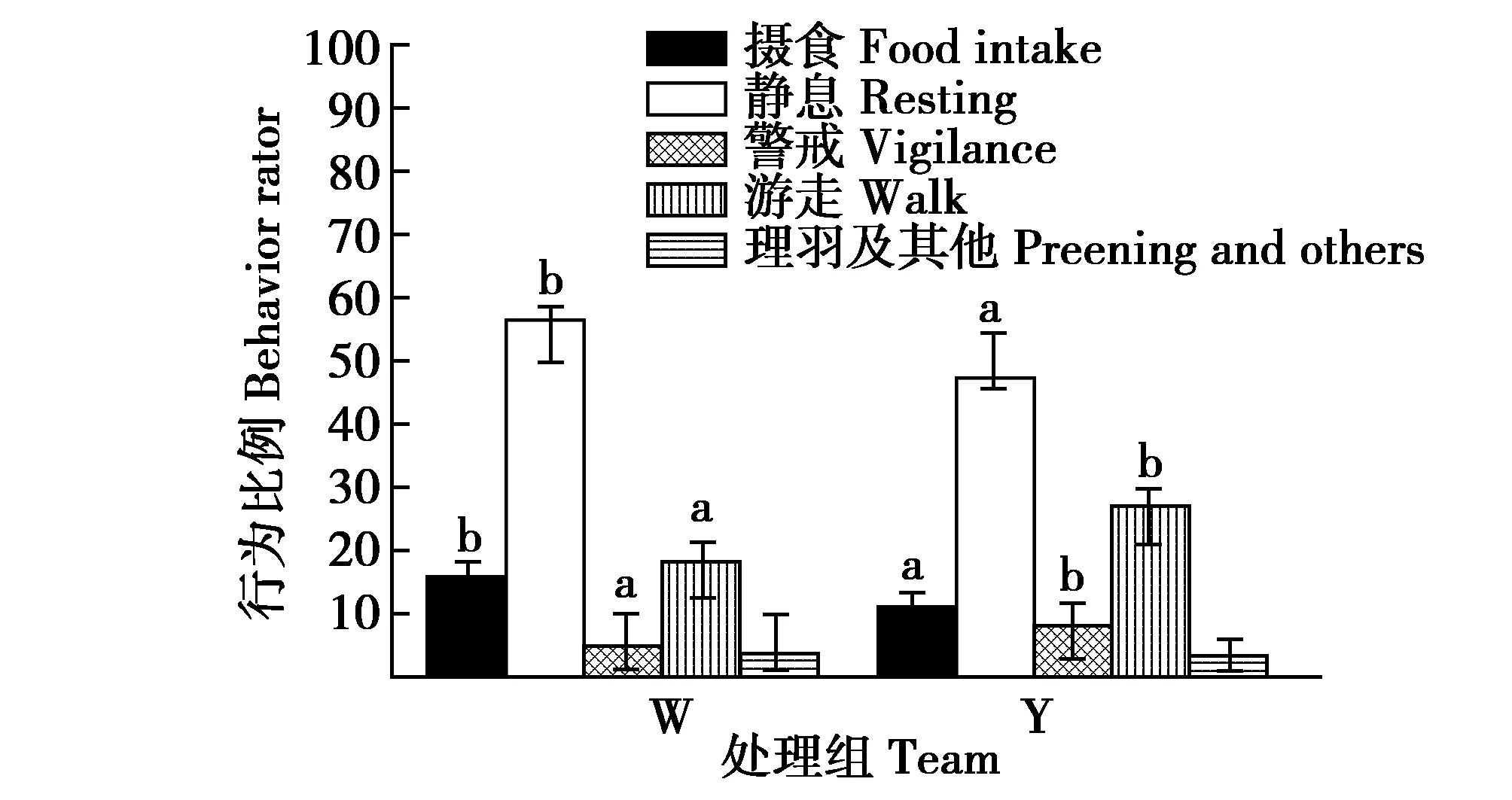
图9 行为及时间分配的组间比较Fig.9 Comparison of behaviors in different captive red crowned crane early winter 注:W为未供给垫草,Y为供给垫草 Note:W denote no mat grass and Y denote supply mat grass
冬末,丹顶鹤白天行为时间分配Y组比W组警戒、游走、摄食、静息、理羽及其他组间差异均不显著(t=2.269,df=12,P=0.065;t=2.233,df=12,P=0.068;t=1.162,df=12,P=0.084;t=0.968,df=12,P=0.088;t=0.535,df=12,P=0.096)。丹顶鹤夜晚的行为组间均是以单足站立静息(藏头于背部)为主和静息位点无规律。
3 讨论
扎龙笼养丹顶鹤能够成功越冬应该具备2个必要条件,即供给足够的食物和施以科学有效的保护对策。冬季供给垫草是扎龙保护区的就地保护对策之一,而这一保护对策是否科学合理的关键却在于供给垫草时间节点的选择与更换垫草的频次。缘于丹顶鹤是国家Ⅰ级保护鸟类和保护区针对笼养丹顶鹤越冬保护的管理要求,在冬季最冷月份本文未能直接对其进行比较研究,并且仅是通过冬初和冬末(各7 d)2时段内垫草对笼养丹顶鹤的作用和影响进行探讨。7 d的实验时间较短,因此也存在导致部分实验结果组间差异不明显的可能性。
3.1 垫草的作用
垫草可以隔凉(降低热传导)和保温。丹顶鹤站立垫草之上,一方面,垫草保温,可避免鹤爪与地面直接接触、降低爪的热传导和减少能耗;另一方面,冬季最冷月份的夜晚,笼养丹顶鹤藏头趴卧于垫草之上,不仅有利于其保持体温和减少能耗,还能确保其跗蹠和爪不被冻伤,这种行为选择对策对于其安全越冬意义重大。笼养丹顶鹤夜晚趴卧行为发生存在个体差异,通常体健个体趴卧行为发生会迟于体弱个体,趴卧时间也会相对缩短。
3.2 供给垫草的必要性
笼养丹顶鹤(体表覆羽足以御寒)成功越冬的关键是确保其跗蹠和爪不被冻伤。丹顶鹤长的跗蹠和爪主要是由骨、筋和角质鳞状表皮及贯穿其内的血管等组织构成,其表面无覆羽。鹤跗蹠关节处及其内侧的静脉血管较粗,肉眼可见血管处表皮微凸(用手轻触能感觉到血管的存在),上跗蹠内侧中部通常也是体检采血的最佳部位。丹顶鹤跗蹠和爪内血管中血液的循环流动应该是其跗蹠和爪能够抵御低温和提高寒冷耐受性的生理基础(热循环)。如果其血管存在“冷涨热缩”的器官响应,那么血管增粗和血流量增加后跗蹠和爪的御寒能力也会随之增强。丹顶鹤跗蹠和爪内血管、血流量与流速是否存在季节性器官响应及其御寒机理等方面都尚未验证。丹顶鹤跗蹠和爪应该具有较强的寒冷耐受性,然而至今未见其寒冷耐受性上限的相关报道。冬日除寒冷压力外,寒风(常刮)也会加速丹顶鹤跗蹠和爪的热散失。丹顶鹤单足交替站立的行为可以减缓其跗蹠和爪的冷压力。但是,若无垫草,笼养丹顶鹤只能一直站立于笼内被冰雪覆盖并冻结的地面之上,一旦被冻伤后严重者无法成功越冬甚至存活,所以其跗蹠和爪应该是其应对冬季严寒的薄弱环节。供给垫草后,笼养丹顶鹤可以选择站立或于夜晚趴卧于垫草之上,使其跗蹠和爪不被冻伤。
总而言之,从动物福利和确保室外笼养丹顶鹤安全越冬考虑(包括保温、减少能耗、确保其跗蹠和爪不被冻伤以及有利于笼养鹤抵御漫长冬夜严寒的生存压力),本文认为越冬期(尤其是冬季最冷月份)保护区有必要为其笼养鹤提供必要的保护——如供给垫草。
3.3 垫草的影响
排泄次数多和不定时随机排泄也是丹顶鹤生物学习性之一。笼养丹顶鹤站立于垫草之上后会发生随机排泄,稀粥状排泄物会混合、粘连并冻结于垫草之中,夜晚鹤趴卧后就会因沾染粪便而使其腹部覆羽发生污损,继之至春季时容易诱发体表寄生虫类疾病。
3.4 体重、行为、排泄和能量收支的比较
3.4.1 体重
动物体重变化是其应对环境变化与自身生理状态改变的一种重要适应[6-10]。丹顶鹤体重(组内)冬初增加和冬末下降(见2.1)应该与冬季气温变化有关。丹顶鹤终体重和体温组间差异均不显著(见2.1),表明有无垫草并未引起其组间体重增减和体温调节的显著变化。但是,本文并不排除冬季最冷月份有无垫草将导致其体重出现显著变化的可能性。
3.4.2 行为发生
丹顶鹤羽色素朴纯洁,体态飘逸雅致,鸣声超凡不俗(诗经·鹤鸣)。丹顶鹤能站不卧的生物学习性有利于其保持体羽清洁、扩大视野范围和提高警戒效率。丹顶鹤会通过行为调节来适应气候变化[11-13]。如冬季丹顶鹤的单足交替站立、藏头静息或藏腿飞行,又如风环境(3~4级以上)中丹顶鹤会选择迎风站立、觅食和警戒时间分配减少、停止飞行和选择避风环境静息等。那么,冬季严寒来临时笼养丹顶鹤对垫草是否会有与之相应的行为发生?
丹顶鹤的行为调节与其应对冬季寒冷的生存压力有关[14]。扎龙保护区冬季气候,冬初和冬末寒冷的生存压力都无法与冬季最冷月份相比,气温低(日平均温度约为-20℃)、漫漫长夜、昼夜温差大(夜晚降温可达10℃左右)、冰天雪地、天寒地冻和常刮3~4级西北冷风等是冬季最冷月份的气候特点。首先,冬初初次供给垫草时丹顶鹤随之发生“舞蹈”行为,可能是其对垫草的好奇,但本文更倾向是其对垫草的渴望和有效御寒的生理需求获得满足。其次,冬初丹顶鹤白天行为时间分配组间差异显著(图9),夜晚有垫草组丹顶鹤则会选择趴卧静息于垫草之上(见2.4),冬末丹顶鹤组间白天行为时间分配和夜晚静息位点选择均无显著差异(见2.4),这表明其行为发生应该与垫草有关。
3.4.3 静息位点与排泄物分布
排泄次数多、随机和无位点选择也是鸟类的生物学习性之一[6,15]。笼养丹顶鹤垫草之上静息时间延长时垫草中的排泄粪便次数(量)会随之增多,垫草之外的排泄次数就会相应减少。因此可以通过笼养丹顶鹤的粪便痕迹来判断其位移轨迹及其行为变化。冬初有垫草组丹顶鹤笼舍内垫草外地面上排泄次数显著减少(表1),这应该与垫草中排泄次数增多有关。冬末笼舍内排泄次数组间差异不显著(见2.3),这表明此时排泄物已随机分布。总之,冬季笼养丹顶鹤排泄物的分布与有无垫草及其行为变化有关。
3.4.4 能量收支
冬季摄入足够的能量和保证能量收支平衡是鸟类能够成功越冬的必要条件之一[16-19]。相关研究表明,扎龙保护区所采用的混合饲喂方法(每日足量供给玉米和定时足量供给鲫鱼)比较科学合理和能够满足笼养丹顶鹤越冬期的摄入能量需求[6]。冬初,摄入玉米的能量、摄入总能量和排出水热能散失(图2)及摄水量(饮水)(图1)组间差异均显著,这表明此时笼养丹顶鹤的能量收支水平及收益会因有无垫草而发生显著变化。冬末,摄入玉米的能量组间差异显著及摄入总能量和排出水热能散失组间差异并不显著(图3,图4),这表明此时笼养丹顶鹤的能量收支水平及收益已与垫草无关。但是,冬初和冬末的生存压力远不及冬季最冷月的生存胁迫,如环境低温,昼短夜长(冬至时接近8 h∶16 h)、昼夜温差大(夜晚降温10℃左右)、夜晚停止摄食(昼出夜伏)和生存能耗增加及漫长黑夜需要消耗更多体内储备的能量。本文认为冬季最冷月份供给垫草能够增加也有必要增加笼养丹顶鹤能量收支的收益。
3.5 建议
3.5.1 供给垫草时间节点的选择
冬季保护区(饲养员)供给垫草起始时间节点的选择主要是依据气温变化并结合经验来确定,主要是为了确保笼养丹顶鹤能够安全越冬。冬初,保护区开始供给垫草1周内夜晚平均最低气温为-17℃,该气温接近野生丹顶鹤越冬地如江苏盐城冬季最低气温(-20℃左右)。但是,笼养鹤(生存空间受限)与野生鹤(可自由选择栖息地)不同;两地气候差异也较大。一方面扎龙保护区从冬初开始光照时间逐渐缩短(直至冬至)、气温还会降低和生存压力将继续增大;另一方面,有无垫草组笼养鹤的行为和能量收支组间差异显著。所以保护区冬初供给垫草时间节点的选择合理。冬末,光照时间逐渐延长、气温逐渐升高、生存压力也将逐渐减小,且笼养鹤的行为和能量收支组间已无显著差异。W组和Y组笼养鹤组内冬末比冬初,摄入能量需求和能量收支水平(图5~图8)均呈现相同的显著递减趋势,这也间接反映出笼养鹤冬末的生存压力远低于冬初。因此保护区停止供给垫草时间节点选择可以适当提前。本文认为供给垫草时间节点的选择既要确保鹤的安全越冬(可考虑采用3.4.3方法来作为时间节点选择的依据之一),还要考虑鹤的健康。首先,冬初供给垫草宜迟不宜早,这样有利于提高笼养丹顶鹤对寒冷的耐受性、适应能力、生理机能及增强体质。其次,冬末停止供给垫草宜早不宜迟。最后,如遇天气骤变和降温幅度较大时还应考虑是否需要供给垫草。总之,缩短供给垫草总的时长对鹤健康有利。
3.5.2 垫草选择、每次添加多少、更换频次和铺放位置
保护区供给的垫草主要是以矮生的芦苇为主并混合其他杂草,本文认为这样垫草选择的优点是芦苇重叠有利于排泄物(稀的粪便)的渗漏和混合杂草可增强趴卧后的保温效果。保护区供给笼养丹顶鹤的垫草量、更换垫草频次和铺放位置都具有较大的随机性,本文认为每次供给每只鹤的草量应以能够满足笼养鹤的趴卧需要(厚度约50 cm(趴卧后厚度会降至10~20 cm)和直径约1.0 m2/只)即可,并根据笼舍内有多少只丹顶鹤来适量增减垫草数量(大量铺放垫草会增加清理笼舍的工作量而并不会提高保护效果)。更换垫草频次应3~4 d更换1次为宜,即少添勤换为原则。垫草最好铺放于笼舍内远离食物和水的供给端(有利于保持垫草清洁和干爽)和靠近挡风板侧(安装挡风板也是保护区的保护对策之一)。
3.5.3 体表寄生虫病的预防
本文认为冬末停止供给垫草后针对笼养丹顶鹤腹部覆羽的污损应进行及时除污、消毒和灭菌处理,以预防、避免或减少春季体表寄生虫病的发生。
总之,虽然本文并没有在冬季最冷月份进行垫草对笼养丹顶鹤作用和影响的比较研究(考虑丹顶鹤的安全),但是本文仍然坚持认为中国北方冬季供给垫草是对室外笼养鹤的必要的保护对策。这不仅仅是动物福利,也是为了能够确保笼养丹顶鹤等鹤类的安全越冬。
[1] IUCN.IUCN Red List categories and criteria,Version 3.1[M].2nd ed.IUCN,Gland,Switzerland and Cambridge,UK,2012.
[2] 蒋志刚,江建平,王跃招,等.中国脊椎动物红色名录[J].生物多样性,2016,24(5):500-551.
[3] 邹红菲,吴庆明.扎龙湿地丹顶鹤和白枕鹤求偶期觅食生境对比分析[J].应用生态学报,2006,17(3):444-449.
[4] 邹红菲,吴庆明,史蓉红.扎龙湿地恢复初期丹顶鹤孵化期觅食生境选择[J].东北林业大学学报,2007,35(7):55-56,58.
[5] 林宝庆,李红魁,刘伟,等.人工饲养丹顶鹤的越冬期管理[J].野生动物,2010,31(5):256-258.
[6] 杨志宏,邹红菲,高忠燕,等.食物可获得性对冬季丹顶鹤能量收支的影响——以扎龙自然保护区人工辅助繁育丹顶鹤为例[J].生态学报,2015,35(13):4408-4415.
[7] Else P L,Brand M D,Turner N,et al.Respiration rate of hepatocytes varies with body mass in birds[J].Journal of Experimental Biology,2004,207(13):2305-2311.
[8] Koenig W D,Walters E L,Walters J R,et al.Seasonal body weight variation in five species of woodpeckers[J].Condor,2005,107(4):810-822.
[9] Burns D J,Ben-Hamo M,Bauchinger U,et al.Huddling house sparrows remain euthermic at night,and conserve body mass[J].Journal of Avian Biology,2013,44(2):198-202.
[10] Wu M X,Zhou L M,Zhao L D,et al.Seasonal variation in body mass,body temperature and thermogenesis in the Hwamei,Garrulaxcanorus[J].Comparative Biochemistry and Physiology Part A:Molecular & Integrative Physiology,2015,179:113-119.
[11] 吴建平,刘振生,李晓民,等.扎龙保护区丹顶鹤繁殖行为观察[J].动物学杂志,2002,37(5):42-46.
[12] 吕士成,陈卫华.环境因素对丹顶鹤越冬行为的影响[J].野生动物,2006,27(6):18-20.
[13] 田秀华,石全华,吕士成,等.半散放丹顶鹤越冬日活动行为[J].生态学杂志,2006,25(9):1077-1081.
[14] 舒莹,胡远满,冷文芳,等.黄河三角洲丹顶鹤秋冬季生境选择机制[J].生态学杂志,2006,25(8):954-958.
[15] 杨志宏,吴庆明,杨渺,等.[树]麻雀羽再生的能量预算和水代谢散热调节[J].生态学报,2014,34(10):2617-2628.
[16] Guillemette M,Butler P J.Seasonal variation in energy expenditure is not related to activity level or water temperature in a large diving bird[J].Journal of Experimental Biology,2012,215(18):3161-3168.
[17] Hegemann A,Matson K D,Versteegh M A,et al.Wild skylarks seasonally modulate energy budgets but maintain energetically costly inflammatory immune responses throughout the annual cycle[J].PLoS One,2012,7(5):e36358.
[18] Li Z Q,Wang Z,Ge C.Time budgets of wintering red-crowned cranes:effects of habitat,age and family size[J].Wetlands,2013,33(2):227-232.
[19] Wu M S,Xiao Y C,Yang F,et al.Seasonal variation in body mass and energy budget in Chinese bulbuls(Pycnonotussinensis)[J].Avian Research,2014,5(1):4.
Positive and Negative Effects of Substrate Grass on Captive Red-Crowned Crane(Grusjaponensis)
Providing grass mats as a substrate for captive red-crowned crane(Grusjaponensis)in winter is an important protection countermeasure at Zhalong National Nature Reserve.To evaluate the effects of substrate grass on captive birds,and to perfect the protection countermeasures,we observed and compared the difference of energy balance,behaviors and body changes of red-crowned crane in cages(6 m×4 m×4 m)with and without mat grass.We recorded data over a period of 7 days in early winter in December 2013 and again in late winter in March 2014 in their natural habitat in Zhalong National Nature Reserve.Fourteen adults older than 5 years of age were divided into two groups,group Y with substrate grass(3 females and 4 males)and group W without substrate grass(3 females and 4 males).After substrate grass was provided in early winter,the corn intake,gross energy intake,and energy of intake corn by group W were significantly greater than by group Y(P﹤0.001).Gross water intake and fluid water intake was significantly lower for group W than for group Y(P﹤0.05).Diurnal frequency of defecation by group Y(on the ground without substrate grass in the cage)was significantly greater than for group W.Diurnal time budgets for feeding,rest,vigilance,and wandering were significantly different between two groups(P﹤0.05).The 5 cranes of group Y lay prone and rested on the substrate grass at night.After the substrate grass was removed in late winter,corn intake was significantly greater for group W than for group Y(P﹤0.05),while gross energy intake,frequency of defecation,diurnal time budget,and nocturnal standing spot of the two groups were similar.In winter,the abdominal coverts were stained due to lying prone and resting on substrate grass at night.In the coldest month of winter,the substrate grass protected the cranes’ instep,metatarsus and claws from frostbite,and aided thermal insulation while reducing energy consumption.It proved best to provide substrate grass in early winter because this enabled cranes to adapt to the cold.Temperatures in early winter in Zhalong approach the lowest temperatures recorded on the wintering grounds of wild cranes that migrate southward to Yancheng City.Cranes of group W did not show frostbite.Based on our results,removal of substrate grass could be ahead of schedule in late winter.Shortening the period of substrate grass and replacing the substrate grass more frequently can reduce the stain on abdominal coverts.In general,substrate grass should be provided in consideration of animal welfare and wintering safety of the cranes.Not only the wintering safety but the health of crane should be taken into consideration when providing substrate grass.
Red-crowned crane(Grusjaponensis);Zhalong Nature Reserve;Substrate grass
稿件运行过程
2016-10-04
修回日期:2016-11-25
发表日期:2017-08-10
Q958.1
A
2310-1490(2017)03-391-09
黑龙江省自然科学基金(C2016059);国家自然科学基金(31470016,31070345)
杨志宏,男,42岁,博士,副教授;主要从事鸟类生理生态学和能量学研究。E-mail:yzh0452@163.com
Wintering in Zhalong Nature Reserve Yang Zhihong1Zou Hongfei2Shao shuli1Guo Liye3Lin Chunshan3Gao Zhongyan3
(1.College of Life Science,Agriculture and Forestry,Qiqihaer University,Qiqihaer, 161006,China;2.College of Wildlife Resources,Northeast Forestry University, Harbin,150040,China;3.Zhalong Nature Reserve,Qiqihaer,161003,China)
