金沙江下游圆口铜鱼生境适宜度曲线的构建
2017-11-10杨志张鹏唐会元龚云董纯陈小娟赵娜
杨志,张鹏,唐会元 龚云 董纯 陈小娟 赵娜
1.水利部中国科学院水工程生态研究所, 武汉 430079 2.水利部水工程生态效应与生态修复重点实验室, 武汉 430079
金沙江下游圆口铜鱼生境适宜度曲线的构建
杨志1,2,*,张鹏1,2,唐会元1,2, 龚云1,2, 董纯1,2, 陈小娟1,2, 赵娜1,2
1.水利部中国科学院水工程生态研究所, 武汉 430079 2.水利部水工程生态效应与生态修复重点实验室, 武汉 430079
金沙江下游的水电梯级开发对长江上游重要特有鱼类圆口铜鱼的自然栖息生境造成了严重影响, 开展生境恢复是保护其资源的重要手段之一。为获得圆口铜鱼的生境恢复目标, 构建圆口铜鱼不同生活史阶段对关键环境因子的适宜度曲线具有重要意义。基于2012—2014年5—7月在金沙江下游宜宾、巧家和皎平渡断面的早期资源采样结果, 以及2016年5—7月在金沙江下游攀枝花至金阳干流江段的野外个体采集和栖息地生境调查结构, 分别构建了圆口铜鱼亲鱼和幼鱼群体对水温、水深、流速和底质类型的适宜度曲线, 结果显示: 圆口铜鱼亲鱼的最适繁殖水温范围为20—25.2℃, 最适栖息水深为1.2—11.5 m, 最适栖息流速为0.2—1.3 m/s, 最适栖息底质类型为小型卵石、大型卵石和巨石,而圆口铜鱼幼鱼栖息的最适水温范围为19.8—25.4℃, 最适水深为0.4—3.95 m, 最适流速为0.1—0.7 m/s, 最适底质类型为细小砾石、中型砾石、大型砾石、小型卵石和大型卵石。上述结果不仅能够为水电工程影响下关键鱼类生境质量评价提供基础数据支撑, 而且能够为鱼类生境保护和修复政策制定提供科学依据。
金沙江下游; 圆口铜鱼; 栖息地评估; 生境恢复; 产卵水温
1 前言
河流生境作为鱼类种群维系的客观载体, 其的完整性对鱼类生物多样性的保护以及河流生态系统服务功能的发挥均具有十分重要的意义[1-2]。尽管如此, 由于水电工程、河道渠道化、围湖开垦、水域污染、航道运输等因素, 许多河流均面临自然生境丧失或改变和生境破碎化等问题[3-7]。作为我国重要的水电能源基地, 目前金沙江流域也不例外地面临自然生境改变、丧失以及生境破碎化等诸多人为导致的问题[8]。已有研究表明, 由于金沙江中下游水电开发导致的水文情势改变、生境阻隔、水温“滞冷”或“滞热”效应等[8]因素, 金沙江中下游许多江段的鱼类资源已受到明显影响, 这其中就包括圆口铜鱼Coreius guichenoti、长鳍吻鮈Rhinogobio ventralis、齐口裂腹鱼Schizothorax prenanti等部分在蓄水前鱼类群落结构中具有优势地位的长江上游特有鱼类[8-10]。
圆口铜鱼属于鲤形目, 鲤科, 鮈亚科, 铜鱼属鱼类, 是金沙江中下游许多干支流江段的重要渔获对象[9-10], 其产卵场目前均仅分布在宜宾以上的金沙江干流和支流雅砻江部分江段[11-12], 但其幼鱼个体在金沙江一期工程蓄水运行前能够分布在三峡库区库中及其以上的2000 km左右江段[13-14]。保护圆口铜鱼的自然种群规模, 不仅可以保护该区域河流食物网结构中重要的组成部分, 而且也可以为人工繁殖群体提供优质的天然种质资源个体[15-16]。尽量恢复因水电建设而导致的自然生境改变或丧失, 对于圆口铜鱼的保护具有重要的意义[11-12], 而自然生境的恢复需要首先了解圆口铜鱼所需要的适宜生境, 以便于在采取适当的恢复措施进行生态恢复时能够有明确的生境恢复目标。因此, 开展适当的鱼类栖息地适宜度评估是重要的, 其中的关键环境因子适宜度曲线的构建更是鱼类栖息地适宜度评估中最重要的工作内容[17-18]。上世纪 80年代以来, 国外学者在对溪红点鲑Salvelinus fontinalis、欧洲茴鱼Thymallus thymallusL.、褐鳟Salmo trutta fario等许多淡水鱼类的栖息地适宜指数(HSI)建模或形成栖息地适宜标准(HSC)时[19-21], 构建了这些鱼类对水温、水深、流速等环境因子的适宜度曲线, 而国内淡水鱼类中除少数几种鱼类(如中华鲟Acipenser sinensis[22]、四大家鱼[22]、洛氏鱥Phoxinus lagowskii[23])外, 圆口铜鱼等绝大多数长江上游重要特有鱼类对环境因子的适宜度曲线缺乏, 这对采取合适的生境恢复措施保护或恢复长江上游特有鱼类资源是不利的。因此, 本文以圆口铜鱼为研究对象, 采用早期资源、野外个体采集以及栖息地生境调查方法, 获取不同水温、水深、流速和底质类型下的圆口铜鱼亲鱼和幼鱼数量, 并基于曲线拟合方法构建圆口铜鱼亲鱼和幼鱼群体对水温、水深、流速和底质类型的适宜度曲线, 旨在为圆口铜鱼栖息地的保护或恢复提供基础数据支撑。
2 材料与方法
2.1 采样区域
采样区域主要为溪洛渡库区金阳县至攀枝花市之间的金沙江下游干流江段(图 1)。采样区域地处四川盆地南缘和云贵高原向四川盆地的过渡区,河道山高谷深, 岭谷高差可达1000—2000 m以上,两岸陡崖连绵, 河道内则滩潭交替, 滩上水浅流急,底质主要由基岩、巨石和卵石组成, 而潭内水深,水流稍缓, 底质多变、复杂, 主要由卵石和砾石构成。采样区域干热河谷气候特征明显, 两岸植被稀疏[24], 有利于鱼类的越冬。由于地形地貌以及海拔高度纵向差异较大, 该区域河流形成了多样化的小生境, 孕育了丰富多样的水生生物, 其中鱼类资源尤为丰富[9-11]。目前, 由于金沙江中下游的梯级开发以及长期的过度捕捞等因素, 该区域的鱼类资源已受到明显的影响[9-11]。
2.2 调查方法
金沙江下游不同水温下圆口铜鱼繁殖群体数量通过2012—2014年间每年5月22日—7月6日在宜宾、巧家、皎平渡断面进行的早期资源调查获得。早期资源的调查方法参考曹文宣等(2007)[25]和唐会元等(2012)[11]等文献, 即在各个调查断面左岸悬挂采样网具进行固定采样, 捞取顺水漂流进网的圆口铜鱼鱼卵, 并同时每天记录各个调查江段的日平均水温(℃)。
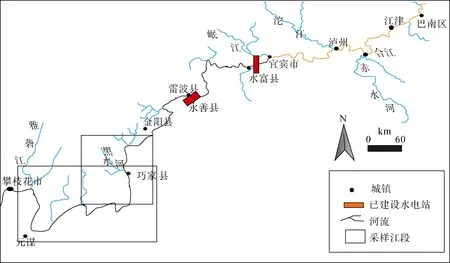
图1 金沙江下游调查区域分布示意图Fig.1 Study area in the lower Jinsha River
2016年5月13日至7月6日在金沙江下游采样区域(图 1)采用刺网等工具行圆口铜鱼亲鱼采集。刺网网目大小为8—12 cm, 网长80—130 m, 网高2—5 m; 自制电捕鱼机采用变频供电方式, 输出电压 220 V。刺网采集时, 根据河流地形特征, 将刺网沉入不同水深的水层中, 同时采用便携式测深仪SM-5A或卷尺测量刺网放置的水深, 采用便携式流速仪LGY-II测量流速, 采用肉眼观察或底泥采集器确定底质类型。对采集到的圆口铜鱼进行解剖或观察第二性征,确定其性腺发育期, 并对发育期III期以上的个体的体长(精确到1 mm)和体重(精确到0.1 g)进行逐尾测量。
2016年5月13日至7月6日在金沙江下游采样区域(图 1)采用虾笼、罾网、底层拖网等行圆口铜鱼幼鱼(1—2龄未成熟个体)采集。虾笼长宽为35 cm×45 cm, 高3 m; 罾网面积为10 m2; 底层拖网也即江津、合江等江段俗称的“奶子网”, 是由一系列带沉子的小型网兜组成, 其规格为高1.5 m, 长约100 m,网目大小2 cm。圆口铜鱼采集到时, 同时采用便携式测深仪 SM-5A、深水温度计 WD-SWJ-73、便携式流速仪LGY-II和肉眼观察或底泥采集器测定该处的水深、水温、流速和底质类型。对采集的圆口铜鱼幼鱼个体进行基础生物学测量, 测量其体长(精确到 1 mm)和体重(精确到 0.1 g)。参考 Schnider et al.(2010)[18]将底质分为 10个类型, 按底质直径从小到大依次为: 有机质及碎屑(org.mat., detritus,0&D)、淤泥(slit, clay, loam, SCL)、细砂(sand<2mm,S)、细小砾石(fine gravel 2—6 mm, FG)、中型砾石(medium gravel 6—20 mm, MG)、大型砾石(large gravel 2—6 cm, LG)、小型卵石(small stones 6—12 cm, SS)、大型卵石(large stones 12—20 cm, LS)、巨石(boulders >20 cm, B)和基岩(Rock, R)。
2.3 数据分析
通过参考余志堂等(1985)[26]所提出的计算公式,计算每天通过宜宾、巧家和皎平渡监测断面的圆口铜鱼鱼卵径流量, 并基于下式估算金沙江下游某一采样断面上游所有产卵场某一天内完成繁殖活动亲鱼的数量:

上式中,Ni为某一采样日i在某一采样断面上游各产卵场完成繁殖活动的圆口铜鱼亲鱼数量(单位: 尾),Ri为某一采样日i通过某一采样断面的圆口铜鱼鱼卵径流量(单位: 粒),AF为金沙江下游圆口铜鱼的平均绝对怀卵量(22496粒/尾)(另文待刊),2.06为金沙江下游圆口铜鱼亲鱼雌雄性比(1.06: 1)的分子和分母之和, 1.06为金沙江下游圆口铜鱼亲鱼雌雄性比的分母[13]。如2012年6月24日在巧家断面采集到8粒圆口铜鱼鱼卵(鱼卵鉴定参考曹文宣等(2007)[25]), 按余志堂等(1985)[26]所提出的计算公式推算出当日通过巧家断面的圆口铜鱼鱼卵径流量为 1369224粒, 根据(1)式可以推得该断面上游各个产卵场完成繁殖活动的亲鱼数量约为 118尾。同时, 该日的平均水温为 24.5 ℃, 因此我们可以近似认为在24.5 ℃有118尾圆口铜鱼亲鱼完成了繁殖。同理, 其它水温下圆口铜鱼的繁殖亲鱼数量也可以估计出来, 从而形成水温与繁殖亲鱼数量的一一对应关系。
以不同水温、水深、流水和底质类型下采集到的圆口铜鱼亲鱼或幼鱼数量为因变量, 以水温、水深、流速和底质类型为自变量, 采用线性方程、对数方程、2次方程、3次方程、复合模型、幂函数、S曲线、生长方程和指数方程9种方程进行曲线拟合, 并选择R2最大的方程为最优拟合方程[27]。曲线拟合前, 不同水温、水深、流水和底质类型下采集到的亲鱼或幼鱼数量均采用函数 log(N+1)进行标准化[28]。最优拟合方程被用来计算单一因子的适宜度,其计算公式为[28]:
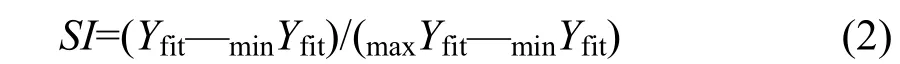
上式中,SI为某一环境因子的适宜度,Yfit为圆口铜鱼亲鱼或幼鱼数量预测值,maxYfit和minYfit分别为圆口铜鱼亲鱼或幼鱼数量预测值中的最大值和最小值。
取SI>0.6之间的水温、水深、流速和底质类型为圆口铜鱼亲鱼或幼鱼群体的最适水温、水深、流速和底质类型区间, 此区间水温、水深、流速和底质类型的适合度均赋值为 1[28], 并参考 Wang &Li(2013)[29]所描述的方法, 最终获得沙江下游圆口铜鱼亲鱼或幼鱼对水温、水深、流水和底质类型的适宜度曲线。
数据统计分析以及数据绘图采用 SPSS 16.0,Excel 2010以及SigmaPlot 11.0软件完成。
3 结果与分析
3.1 鱼类与环境因子的关系
3.1.1 繁殖群体
2012—2014年 5—7月在金沙江下游各产卵场完成繁殖活动的圆口铜鱼亲鱼共有2234尾, 其产卵水温处于 18—26 ℃之间, 其中圆口铜鱼在22.1—23 ℃之间发生繁殖活动的个体数量最多, 为680尾, 占调查期间产卵个体总数的 30.44%, 其次为20.1—21 ℃区间, 共有591尾圆口铜鱼完成繁殖活动, 占 26.45%, 最少为 26.1—27.0区间, 仅有 1尾圆口铜鱼完成繁殖活动, 占0.04%(图2)。
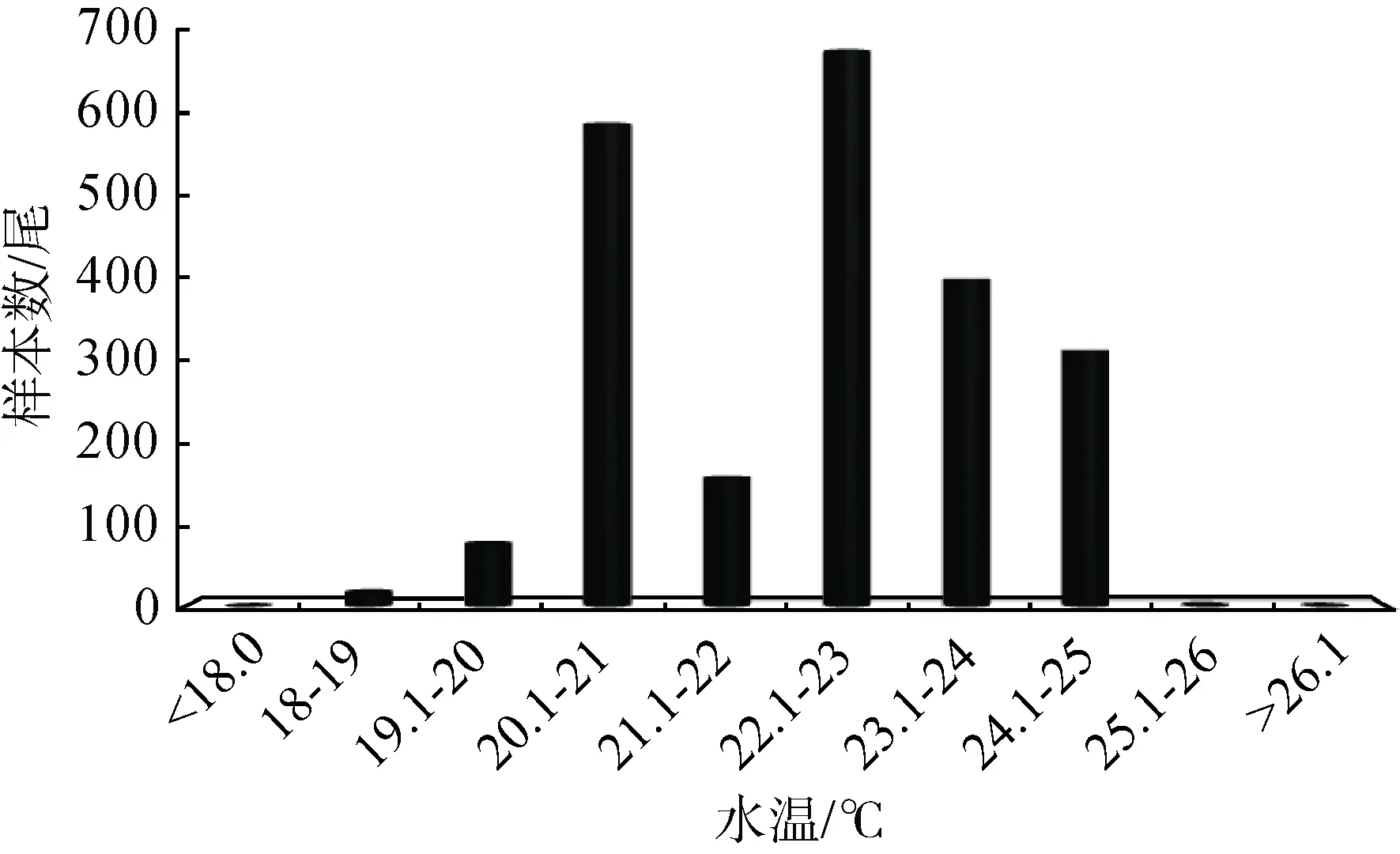
图2 2012—2014年间金沙江下游各产卵场圆口铜鱼繁殖群体数量与水温的关系Fig.2 The relationships between the individual numbers of mature C.guichenoti in the spawning grounds of the lower Jinsha River and the water temperatures during the period 2012-2014
2016年5—7月在巧家至攀枝花江段采集到圆口铜鱼繁殖个体 129尾, 其体长范围为 213—433 mm, 平均体长为356 mm, 体重范围为220.9—1105.0 g, 平均体重为649.6 g。同时, 129尾圆口铜鱼被采集到时的流速分布范围为 0.26—1.67 m·s-1, 平均值为 0.64 m·s-1, 水深分布范围为2.02—19.2 m, 平均值为6.22 m(图3A)。此外, 还记录了 129尾圆口铜鱼被采集时的底质类型, 其中大型卵石上采集到的圆口铜鱼亲鱼数量最多, 共32尾,占总采集尾数的 24.81%, 其次为巨石上, 共 28尾,占 21.71%, 最少为中型砾石上, 仅采集到 3尾, 占2.33%(图 3B)。
3.1.2 幼鱼群体
2016年5—7月在金沙江下游巧家至攀枝花江段采集到圆口铜鱼幼鱼 388尾, 其体长分布范围为39—160 mm, 平均体长为111 mm, 体重分布范围为0.5—60.6 g, 平均体重为15.8 g。水温在22.1—23℃之间采集的幼鱼个体数量最多, 共143尾, 占总采集数量的 36.86%, 其次为 23.1—24.0 ℃区间, 共采集到幼鱼个体 92尾, 占 23.71%, 最少为<19 ℃和26.1—27 ℃组, 均仅采集到1尾, 占0.26%; 水深在0.91—1.20 m时采集到的圆口铜鱼幼鱼个体数最多,共 98尾, 占总采集数量的 25.26%, 其次为1.21—1.50 m组, 76尾, 占19.59%, 最少为<0.1 m组,仅1尾, 占0.26%; 流速在0.41—0.50 m·s-1时采集到的圆口铜鱼幼鱼个体数最多, 共 145尾, 占总采集数量的 37.37%, 其次为 0.31—0.40 m·s-1组, 90尾,占 23.20%, 最少为>0.71 m·s-1组, 5尾, 占 1.29%;在大型砾石(Large gravel, LG)上采集到的圆口铜鱼幼鱼个体数最多, 共 120尾, 占总采集数量的30.93%, 其次为小型卵石(Small stones, SS), 95尾,占24.38%, 最少为基岩(Rock, R), 仅1尾, 占0.26%(图 4)。
3.2 单因子适宜度曲线的建立
3.2.1 繁殖群体
环境因子和繁殖群体数量的最优拟合曲线如下表1所示。各个环境因子与繁殖群体数量的拟合曲线均以三次曲线为最优, 其中底质类型与繁殖群体数量的拟合程度最高,R2为0.990(表1)。

图3 2016年5—7月金沙江下游圆口铜鱼亲鱼采集时的水深和流速分布(A)和不同底质类型上采集到的圆口铜鱼数量(B)(MG:中型砾石; LG: 大型砾石; SS: 小型卵石; LS: 大型卵石; B: 巨岩; R: 基岩)Fig.3 Distributions of water depths and velocities when mature individuals of C.guichenoti were sampled in the lower Jinsha River from May to July in 2016 (A) and the quantities of mature individuals of C.guichenoti which were sampled on the different substratum types in the same sampled sites and times(MG: medium gravel; LG: large gravel; SS: small stones; LS:large stones; B: boulder; R: rock)
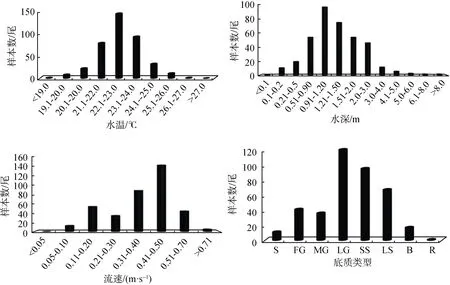
图4 2016年5—7月在金沙江下游不同水温、水深、流速和底质类型下采集到的圆口铜鱼幼鱼个体数(S: 细砂;FG:细小砾石;MG: 中型砾石; LG: 大型砾石; SS: 小型卵石; LS: 大型卵石; B: 巨岩; R: 基岩)Fig.4 The numbers of juveniles of C.guichenoti sampled under the different water temperatures, water depths, velocities and substratum types in the lower Jinsha River from May to July in 2016(S: sand; FG: fine gravel; MG: medium gravel; LG: large gravel; SS: small stones; LS: large stones; B: boulder; R: rock)
基于最优拟合方程以及公式 (2), 计算不同环境因子下圆口铜鱼亲鱼对环境条件的适宜度, 并最终得到各个因子的适宜度曲线(图5)。圆口铜鱼亲鱼的最适繁殖水温范围为20—25.2 ℃, 最适栖息水深为 1.2—11.5 m, 最适栖息流速为0.2—1.3 m·s-1,最适栖息底质类型为小型卵石、大型卵石和巨石(图5)。

表1 不同环境因子与圆口铜鱼繁殖群体数量的最优拟合曲线Tab.1 The optimal fitted curves for the relationships between water temperatures, water depths, velocities,substratum types and corresponding numbers of mature individuals of C.guichenoti
3.2.2 幼鱼群体
环境因子和幼鱼群体数量的最优拟合曲线如下表2所示。各个环境因子与幼鱼群体数量的拟合曲线均以三次曲线为最优, 其中水温与幼鱼群体数量的拟合程度最高,R2为0.983(表2)。
基于最优拟合方程以及公式(2), 计算不同环境因子下圆口铜鱼幼鱼对环境条件的适宜度, 并最终得到各个因子的适宜度曲线(图6)。圆口铜鱼幼鱼栖息的最适水温范围为 19.8—25.4 ℃, 最适水深为0.4—3.95 m, 最适流速为0.1—0.7 m·s-1, 最适底质类型为细小砾石、中型砾石、大型砾石、小型卵石和大型卵石(图6)。

表2 不同环境因子与圆口铜鱼幼鱼群体数量的最优拟合曲线Tab.2 The optimal fitted curves for the relationships between water temperatures, water depths, velocities ,substratum types and corresponding numbers of juvenileindividuals of C.guichenoti
4 讨论
水温作为影响鱼类生长发育最重要的生态因子之一, 其的变化不仅会影响到鱼类个体不同生活史阶段的摄食、呼吸等生命活动, 而且也会对鱼类的繁殖造成明显的影响[30-31]。因此, 在构建某一鱼类的适宜度曲线时, 水温通常是优先考虑的环境因子[19-21]。Hightower et al.(2012) 在采用贝叶斯模型对美国鲥鱼Alosa sapidissima产卵栖息地适宜度模型构建的过程中, 基于专家咨询的方式选择了影响美国鲥鱼产卵最重要的环境因子,其结果也显示100%的专家选择了水温和流速作为美国鲥鱼产卵最重要的环境因子[32]。圆口铜鱼作为广泛分布在长江中上游的鱼类, 其的产卵场分布范围广泛, 历史上从金沙江中游金安桥库区到金沙江下游屏山附近江段均有其分布[12], 这将导致其繁殖水温具有较宽的分布范围, 因此对所有圆口铜鱼产卵场在整个繁殖期间的连续调查是必须的。然而, 随着金沙江中下游水电开发的进行,特别是2012年10月至2014年10月之间金沙江下游向家坝、溪洛渡水电站以及金沙江中游龙开口、鲁地拉和观音岩水电站的蓄水发电, 导致原有分布在金沙江中下游的圆口铜鱼产卵场分布范围在2014年后大大缩小(产卵场压缩到巧家至攀枝花之间江段), 从而使得在2014年后现场测定的繁殖水温范围变得狭窄, 其不能真实反映圆口铜鱼自然繁殖的分布范围, 因此在本文中采用2012年10月以前多个早期资源监测断面的连续水温数据替代2016年现场水温测定数据。
在本文中, 其它3个选择的环境因子(水深、流速和底质类型)也是构建鱼类适宜度曲线最优先选择的环境变量[18-22,32]。已有研究表明, 鱼类的关键生活史阶段如洄游、迁徙、产卵行为、幼鱼摄食生长、群体补充等, 都与水深、流速和底质类型有密切关系[33-35]。圆口铜鱼作为典型江河流水性底层鱼类[12], 其成鱼对流速的偏好, 决定了这种鱼类空间分布与流速、水深的直接关系,而且圆口铜鱼的产卵活动与流速的改变具有明显的相关关系[12,36], 这也决定了流速大小对其生活史的重要影响。同时, 圆口铜鱼产卵场一般分布于江面由宽变窄, 水深流急, 滩多、碛坝多、底为砾石的江段[12], 显示了圆口铜鱼在关键生活史阶段对底质类型的选择。
本文中的单因子适合度曲线采用生境利用法拟合环境与物种数量获得, 而不考虑某一有限生境类型的可获取性[17], 这与实际采样的可行性有关。由于圆口铜鱼个体主要分布在干流河段中, 这些河段生境复杂, 河面宽阔, 因此在缺乏高精度实测的河道地形特征下, 无法准确分类某一采样江段的生境类群。而且对于圆口铜鱼这类栖息于河道中间的物种(midstream species)[37]而言, 其的采样在高流量时(产卵季节)也是比较困难的, 这也会增加生境类型识别与面积估算时所出现的误差。同时,样本数量的大小也会影响适宜度曲线的准确性。Riedl & Peter (2013)推荐在构建适宜度曲线时的样本数量应大于 100尾[38], 本文的样本数量满足了这一要求。此外, 样本采集时也覆盖了采样区域所有生境类型(图3和图4)。尽管如此, 由于圆口铜鱼栖息地与产卵场分布的广泛性[12]以及鱼类对不同环境适应所导致的适宜度的可变性[39], 因此本文的结果很可能仅适用于金沙江下游干流区域。

图5 金沙江下游圆口铜鱼亲鱼对水温、水深、流水和底质类型的适宜度曲线(LG: 大型砾石; SS: 小型卵石; B: 巨岩; R: 基岩)Fig.5 Water temperature, water depth, velocity and substratum type suitability for the mature C.guichenoti of the lower Jinsha River (LG: large gravel; SS: small stones; B: boulder; R: rock)
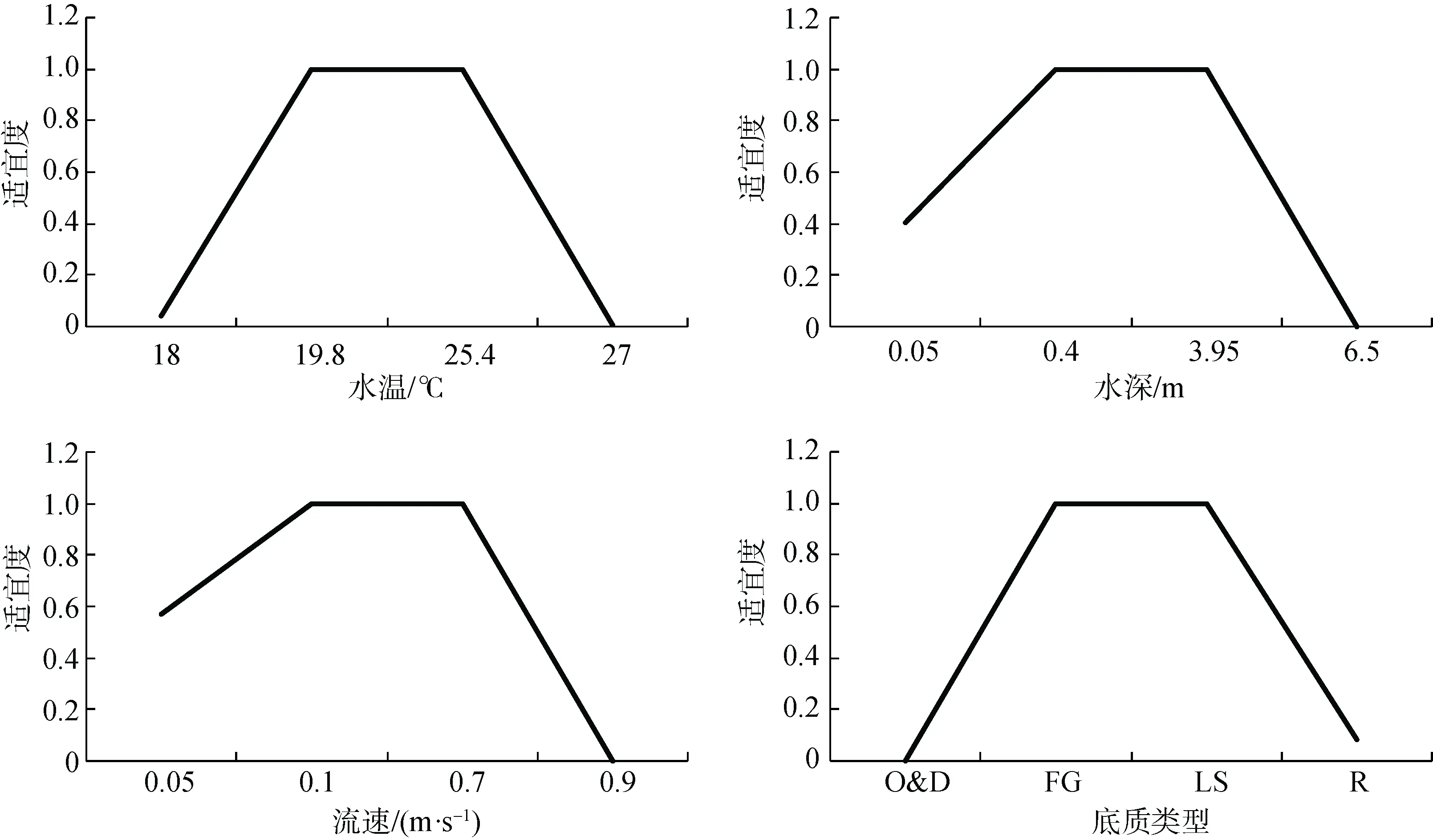
图6 金沙江下游圆口铜鱼幼鱼对水温、水深、流水和底质类型的适宜度曲线(O&D: 有机质及碎屑; FG: 细小砾石; LS: 大型卵石; R: 基岩)Fig.6 Water temperature, water depth, velocity and substratum type suitability for the juvenile C.guichenoti of the lower Jinsha River
2012—2014年间圆口铜鱼在金沙江下游产卵时的最低水温为 18.2 ℃, 然而刘乐和等(1990)[36]根据1981—1986年在宜昌至屏山的早期资源调查结果,认为两种铜鱼产卵最低水温为17 ℃, 低于17 ℃时未见产卵活动发生, 因此图 5中水温的适合度曲线左端未闭合, 显示在 18.2℃以下时圆口铜鱼仍很可能完成自然繁殖活动。同理, 流速在1.7 m·s-1以上时圆口铜鱼仍可能有一定适宜度, 这是因为在产卵季节, 金沙江下游圆口铜鱼产卵场中流速大于 1.7 m·s-1的区域分布较多, 但是在这些区域采样危险性很高, 本文中的采样实际未涉及到这些区域(图3)。圆口铜鱼亲鱼栖息的最适水深较大, 且分布范围较宽(1.2—11.5 m), 显示圆口铜鱼对不同水深的高度适应性。已有研究表明, 更大规格个体的鱼类倾向于栖息在更深的水体之中[40]。此外, 圆口铜鱼亲鱼喜栖息在卵石和巨石之上, 这很可能与其食性有关。圆口铜鱼喜食淡水壳菜等软体动物[13], 而这些动物通常吸附在卵石和巨石上面。
金沙江下游圆口铜鱼幼鱼对环境条件的适宜度曲线与亲鱼对环境条件的适宜度曲线存在一定差异,其最适水深、流速范围明显小于亲鱼, 而最适水温、底质类型范围大于亲鱼(图5和图6), 显示幼鱼群体更喜欢生活在岸边流速较缓区域, 且这些幼鱼个体对水温、底质类型的要求不高, 反映了幼鱼对不同生境较强的环境适应力。已有研究也表明, 在三峡库区淤泥底质的浅水缓流区域采用罾网就能够捕捞大量的圆口铜鱼幼鱼个体[14],进一步证实了图6结果的可靠性。
[1]ZHU Di, CHANG Jianbo.Annual variations of biotic integrity in the upper Yangtze River using an adapted index of biotic integrity (IBI)[J].Ecological Indicators, 2008, 8(5):564–572.
[2]HOLMLUND C M, HAMMER M.Ecosystem services generated by fish populations[J].Ecological Economics,1999, 29(2): 253–268.
[3]DEJALON D G, SANCHEZ P.Downstream effects of a new hydropower impoundment on macrophyte, macroinvertebrate and fish communities[J].Regulated Rivers: Research and Management, 1994, 9(4): 253–261.
[4]LAU J K, LAUER T E, WEINMAN M L.Impacts of channelization on stream habitats and associated fish assemblages in east Central Indiana[J].American Midland Naturalist, 2006, 156(2): 319–330.
[5]曹文宣.长江鱼类资源的现状与保护对策[J].江西水产科技, 2011, 2: 1–4.
[6]KIME D E.The effects of pollution on reproduction in fish [J].Reviews in Fish Biology & Fisheries, 1995, 5(1): 52–95.
[7]黄亮亮, 李建华, 鹿野雄一, 等.东苕溪中下游内河航运对鱼类群落的影响[J].水生态学杂志, 2013, 34(5):30–37.
[8]CHENG Fei, LI Wei, CASTELLO L, et al.Potential effects of dam cascade on fish: lessons from the Yangtze River[J].Reviews in Fish Biology & Fisheries, 2015, 25(3): 1–17.
[9]高少波, 唐会元, 乔晔, 等.金沙江下游干流鱼类资源现状研究[J].水生态学杂志, 2013, 34(1): 44–49.
[10]杨志, 唐会元, 朱迪, 等.金沙江干流攀枝花江段鱼类种类组成和群落结构研究[J].水生态学杂志, 2014, 35(5):43–51.
[11]唐会元, 杨志, 高少波, 等.金沙江中游圆口铜鱼早期资源现状[J].四川动物, 2012, 31(3): 416–421.
[12]张轶超.大坝建设对长江上游圆口铜鱼和长鳍吻鮈自然繁殖的影响[D].武汉: 中国科学院水生生物研究所,2009: 21–52.
[13]程鹏.长江上游圆口铜鱼的生物学研究[D].武汉: 华中农业大学, 2008: 12–21.
[14]杨少荣, 马宝珊, 孔焰, 等.三峡库区木洞江段圆口铜鱼幼鱼的生长特征及资源保护[J].长江流域资源与环境,19(Z2): 52–57.
[15]李晓东, 危兆盖, 黄照, 等.长江珍稀特有鱼类圆口铜鱼人工驯养繁殖成功[J].水产科技情报, 2014(5): 268–269.
[16]李斌, 王志坚, 杨洁萍, 等.三峡库区干流鱼类食物网动态及季节性变化[J].水产学报, 2013, 37(7): 1015–1022.
[17]易雨君, 程曦, 周静.栖息地适宜度评价方法研究进展[J].生态环境学报, 2013, 22(5): 887–893.
[18]SCHNEIDER M, NOACK M, GEBLER T, et al.Handbook for the Habitat Simulation Model.Casimir.Module Casimir-Fish.Base version[M].Institut fur Wasserbau, Universitat Stuttgart, 2010: 12.
[19]COON T G.Development and evaluation of alternative habitat suitability criteria for brook trout[J].Transactions of the American Fisheries Society, 1997, 126(1): 65–76.
[20]FUKUDA S, BAETS B D, WAEGEMAN W, et al.Habitat prediction and knowledge extraction for spawning European grayling (Thymallus thymallusL.) using a broad range of species distribution models[J].Environmental Modelling & Software, 2013, 47(2): 1–6.
[21]MUNOZ-MAS R, MARTINEZ-CAPEL F, SCHNEIDER M, et al.Assessment of brown trout habitat suitability in the Jucar River Basin (SPAIN): Comparison of data-driven approaches with fuzzy-logic models and univariate suitability curves[J].Science of the Total Environment,2012, 440(3): 123–131.
[22]李建, 夏自强, 戴会超, 等.三峡初期蓄水对典型鱼类栖息地适宜性的影响[J].水利学报, 2013, 39(8): 892–900.
[23]康鑫, 张远, 张楠, 等.太子河洛氏鱥幼鱼栖息地适宜度评估[J].生态毒理学报, 2011, 6(3): 310–320.
[24]欧晓昆, 金振洲.金沙江干热河谷植物区系和生态多样性的初步研究[J].植物科学学报, 1996, 14(4): 318–322.
[25]曹文宣, 常剑波, 乔晔, 等.长江鱼类早期资源[M].北京: 中国水利水电出版社, 2007.
[26]余志堂, 周春生, 邓中粦, 等.葛洲坝水利枢纽工程程截流的长江四大家鱼产卵场[C]// 中国鱼类学会, 鱼类学论文集(第四辑)[M].北京: 科学出版社, 1985: 1–11.
[27]陶江平, 乔晔, 杨志, 等.葛洲坝产卵场中华鲟繁殖群体数量与繁殖规模估算及其变动趋势分析[J].水生态学杂志, 2009, 2(2): 37–43.
[28]VAYGHAN A H, POORBAGHER H, SHAHRAIYNI H T,et al.Suitability indices and habitat suitability index model of Caspian kutum (Rutilus frisii kutum) in the southern Caspian Sea [J].Aquatic Ecology, 2013, 47(4): 441–451.
[29]WANG Fei, LIN Binliang.Modelling habitat suitability for fish in the fluvial and lacustrine regions of a new Eco-City[J].Ecological Modelling, 2013, 267: 115–126.
[30]文良印, 谭玉钧.水温对草鱼鱼种摄食, 生长和死亡的影响[J].水产学报, 1998, 22(4): 371–374.
[31]WANG Junna, LI Chong, DUAN Xinbin, et al.Variation in the significant environmental factors affecting larval abundance of four major Chinese carp species: fish spawning response to the Three Gorges Dam[J].Freshwater Biology,2014, 59(7): 1343–1360.
[32]HIGHTOWER J E, RAABE J K, DREW C A, et al.A Bayesian spawning habitat suitability model for American shad in southeastern United States rivers[J].Journal of Fish& Wildlife Management, 2012, 3(2): 184–198.
[33]GREGORY E S, CHRIS J J, SHRIMPTON J.M.Temperature and flow effects on migration timing of Chinook salmon smolts[J].Transactions of the American Fisheries Society, 2009, 138(6): 1252–1265.
[34]殷名称.鱼类生态学[M].北京: 中国农业出版社, 1995:180–185.
[35]OSMUNDSON D B, RYEL R J, LAMARRA V L, et al.Flow-sediment-biota relations: implications for river regulation effects on native fish abundance[J].Ecological Applications, 2002, 12(6): 1719–1739.
[36]刘乐和, 吴国犀, 王志玲.葛洲坝水利枢纽兴建后长江干流铜鱼和圆口铜鱼的繁殖生态[J].水生生物学报,1990, 14 (3): 205–215.
[37]DAUFRESNE M, VESLOT J, CAPRA H, et al.Fish community dynamics (1985–2010) in multiple reaches of a large river subjected to flow restoration and other environmental changes[J].Freshwater Biology, 2015, 60(6):1176–1191.
[38]RIEDL C, PETER A.Timing of brown trout spawning in Alpine rivers with special consideration of egg burial depth[J].Ecology of Freshwater Fish, 2013, 22(3): 384–397.
[39]MARY C F, ZACHARY H.B, JOHNIE H C.Transferability of habitat suitability criteria for fishes in warm water streams[J].North American Journal of Fisheries Management, 1997, 17(1): 20–31.
[40]HARVEY B C, STEWART A J.Fish size and habitat depth relationships in headwater streams[J].Oecologia, 1991,87(3): 336–342.
The formation of habitat suitability curves for Coreius guichenoti (Sauvage &Dabry de Thiersant, 1874) of the lower Jinsha River
YANG Zhi1,2,*, ZHANG Peng1,2, TANG Huiyuan1,2, GONG Yun1,2, DONG Chun1,2, CHEN Xiaojuan1,2, ZHAO Na1,2
1.Institute of Hydroecology,Ministry of Water Resources and Chinese Academy of Sciences,Wuhan430079,China2.Key Laboratory of Ecological Impacts of Hydraulic-projects and Restoration of Aquatic Ecosystem of Ministry of Water Resources,Wuhan430079,China
Coreius guichenotiis an important endemic fish species in the lower reaches of the Jinsha River.Due to the cascade hydropower development, its nature habitats suffered serious impacts.It is urgent to protect its habitat and restore its population resources.In order to obtain habitat restoration goals, obtaining the key environmental factors and forming the suitability curves for the different life history stages ofC.guichenotiare very important.In this study, we sampled the eggs in the Yibin, Qiaojia and Jiaopingdu sections from May to July in 2012-2014, and collected filed individual and surveyed the habitat in the lower reaches of the Jinsha River (from Panzhihua to Jinyang)during the period from May to July in 2016.Based on these data, we set up the suitability curves of water temperature, water depth, velocity and substratum type for the parent individuals ofC.guichenotiand its juveniles.The results showed that for adults (1) the range of the optimum breeding water temperature was 20-25.2 ℃, (2) the range of the most suitable habitat depth was 1.2—11.5 m, (3) the range of the most suitable velocity was 0.2-1.3 m·s-1, and (4) the most suitable substratum types were small stones, large stones and boulders.For the juveniles (1) the optimum temperature range was 19.8-25.4 ℃, (2)the optimum depth range was 0.4—3.95 m, (3) the optimal velocity range was 0.1—0.7 m·s-1, and (4) the most suitable types of substratum were fine gravel, medium gravel, large gravel, small stones and large stones.This study not only can provide basic data for the habitat quality assessment of the critical fish species under the influences of hydropower projects, but also can provide a scientific basis for the policy formulation of fish habitat protection and restoration.
the lower Jinsha River;Coreius guichenoti; habitat assessment; habitat restoration; spawning temperature
10.14108/j.cnki.1008-8873.2017.05.017
S932.4, X176
A
1008-8873(2017)05-129-09
杨志, 张鹏, 唐会元, 等.金沙江下游圆口铜鱼生境适宜度曲线的构建[J].生态科学, 2017, 36(5): 129-137.
YANG Zhi, ZHANG Peng, TANG Huiyuan, et al.The formation of habitat suitability curves forCoreius guichenoti(Sauvage & Dabry de Thiersant, 1874) of the lower Jinsha River [J].Ecological Science, 2017, 36(5): 129-137.
2016-08-26;
2016-11-03
梯级开发背景下生境大小及其分隔对鱼类物种迁徙—灭绝动态研究(51379134); 西南河流典型鱼类对梯级水电开发响应与增殖放流技术(2016YFC0502206); 武汉大学测绘遥感信息工程国家重点实验室开放基金(15ZD04); Jinsha River Basin Integrated Water Resources and Risk Management under Changing Climate (425988607)
杨志(1982—), 男, 湖南芷江人, 硕士, 助理研究员, 主要从事鱼类生态学研究, E-mail: yangzhi4626@163.com
*通信作者:杨志, 男, 硕士, 助理研究员, 主要从事鱼类生态学研究, E-mail: yangzhi4626@163.com
