磷浓度与氮磷比对蛋白核小球藻氮磷吸收效应的影响
2017-11-10曹煜成李卓佳胡晓娟徐煜孙志伟李奕雯文国樑
曹煜成, 李卓佳, 胡晓娟, 徐煜, 孙志伟, 李奕雯, 文国樑,*
1.中国水产科学研究院南海水产研究所, 农业部南海渔业资源开发利用重点实验室, 广东省渔业生态环境重点实验室, 广州510300 2.广东海洋与渔业出版传媒有限公司, 广州 510220
磷浓度与氮磷比对蛋白核小球藻氮磷吸收效应的影响
曹煜成1, 李卓佳1, 胡晓娟1, 徐煜1, 孙志伟1, 李奕雯2, 文国樑1,*
1.中国水产科学研究院南海水产研究所, 农业部南海渔业资源开发利用重点实验室, 广东省渔业生态环境重点实验室, 广州510300 2.广东海洋与渔业出版传媒有限公司, 广州 510220
为明确在不同磷浓度及氮磷比(N/P)的协同影响下蛋白核小球藻(Chlorella pyrenoidosa) 的生长和氮磷吸收特性, 文章将微藻培养液中的磷酸盐浓度设为 0.50 mg·L–1、0.25 mg·L–1、0.05 mg·L–1, 并将 N/P 分别设为 32、16、8, 每2 d取样测定藻细胞数量和氮磷指标, 实验设9个组, 每组3平行, 周期12 d。结果显示, 在低氮磷浓度组藻细胞数量明显低于高氮磷组(P<0.05), 氮磷浓度对藻细胞生长的影响远大于N/P(P<0.05); 在高氮磷浓度组, 当N/P低于8时, 藻细胞生长受到限制, 当其高于16时, N/P不再成为限制性因子; 不同N/P下初始氮磷浓度的影响大于N/P。由藻细胞与氮磷指标的多元回归分析发现, 藻细胞数量与氮磷浓度, 以及不同取样时间的氮磷浓度等多个因子存在显著线性关系, 其中初始氮磷浓度影响较大。上述结果表明, 蛋白核小球藻数量与氮磷的消耗呈正相关, 以期仅通过调整 N/P的方式影响其对水环境中氮磷的吸收, 恐难达到良好效果。
磷; 氮磷比; 蛋白核小球藻; 吸收效应
1 前言
蛋白核小球藻(Chlorella pyrenoidosa)是养殖池塘微藻群落中的常见优势种[1-4], 它不仅可作为养殖生物幼体和浮游动物的天然饵料, 还能起到稳定和优化水体微藻群落结构, 快速吸收转化水体氨氮、亚硝酸盐等有毒有害物质[5,6], 促进养殖生物健康生长的良好效果[7]。通常养殖水体中的氮磷富营养程度较高[5,8], 微藻作为水体生态系统中的生产者, 可将之进行高效吸收利用, 实现养殖水质的净化[9]。有研究表明, 不同种类的微藻对氮磷的营养要求存在较大差异[10-18], 并且不同的光照和温度条件下其氮磷需求量也差别巨大[19,20]。其实, 氮磷浓度固然与微藻生长密切相关, 氮磷间的数量比例对微藻亦同样重要[21], 以往的研究大多或关注微藻的氮磷数量需求[22,23], 或是仅关注氮磷比对微藻生长的影响[24],而自然环境下以上两种因素均客观存在, 并对微藻生长共同产生影响, 仅进行单因子实验无法科学阐释究竟何种氮磷因子对微藻自然状态的影响贡献更大, 因此, 有必要将氮磷营养浓度和比例相结合,并以统计学分析方法评估两者的协同影响效应。该实验根据对虾养殖池塘磷浓度水平的动态特征[25],选取三个磷浓度水平为基础, 明确不同氮磷浓度和氮磷比条件下, 蛋白核小球藻的生长及其对氮磷的吸收效应, 并以多元回归分析法研究藻细胞数量(YCell)与初始氮浓度(XN0)、初始磷浓度(XP0)、初始氮磷比(XN/P0)、生长时间(XT), 以及不同取样时间的氮浓度(XNv)、磷浓度(XPv)、氮磷比(XN/Pv)之间的相关性, 建立回归方程。以期为今后进一步探索构建适宜养殖池塘有益微藻群落的氮磷营养调控策略提供基础参考依据。
2 材料与方法
2.1 材料
蛋白核小球藻(Chlorella pyrenoidosa)分离自广东省汕尾市红海湾的对虾集约化养殖池塘。小球藻的基础培养基为 BG11液体培养基[10,11], 根据实验需求利用分析纯的硝酸铵(NH4NO3)、磷酸二氢钾(KH2PO4)调节培养基中的氮磷浓度水平和氮磷比。
2.2 试验设计
根据集约化对虾养殖池塘生产过程中水体活性磷酸盐的 0.01 mg·L–1—0.50 mg·L–1的变化范围[25],分别将培养基中的磷酸盐浓度设为 0.05 mg·L–1、0.25 mg·L–1、0.50 mg·L–1, 并以磷酸盐浓度为基础按质量比的关系设置3个氮磷比(N/P)水平, 即32、16、8。由此将实验组分为 9个组, 分别为:C1(P 0.50 mg·L–1, N/P 32)、C2(P 0.50 mg·L–1, N/P 16)、C3(P 0.50 mg·L–1, N/P 8)、C4(P 0.25 mg·L–1, N/P 32)、C5(P 0.25 mg·L–1, N/P 16)、C6(P 0.25 mg·L–1, N/P 8)、C7(P 0.05 mg·L–1, N/P 32)、C8(P 0.05 mg·L–1, N/P 16)、C9(P 0.05 mg·L–1, N/P 8)。每组均设三个平行,实验周期12 d。
2.3 小球藻的培养
小球藻藻种经过扩大培养后, 藻液于6300 g、4℃下离心6 min, 藻沉淀用无菌双蒸水反复洗涤 3次,最后以蒸馏水重悬藻沉淀, 取500 µL在显微镜下检查藻细胞情况并准确计数。根据藻原液计数结果,按 2.4×105cell·mL–1浓度将藻细胞接种至不同实验组的培养瓶, 培养液盐度为6。将培养瓶置于光照培养箱中恒温培养, 温度25℃, 光暗比12 h :12 h, 光照强度4000—5000 lx。每6 h对各培养瓶进行手摇振荡, 避免藻细胞贴壁生长。
2.4 样品采集与检测
实验过程中每2 d取样一次进行小球藻数量和氮磷水质指标的测定。藻样加甲醛固定, 在显微镜下用血球计数板准确计数小球藻细胞数量; 水样按照国家标准方法[26], 以萘乙二胺分光光度法测定亚硝酸盐(NO2–-N), 紫外分光光度法测定硝酸盐(NO3–-N), 次溴酸盐法测定氨氮(NH3-N), 磷钼蓝比色法测定磷酸盐(PO43–-P)。考虑到无机氮在水体环境中具有亚硝酸盐氮(NO3–-N)、硝酸盐(NO2–-N)、氨氮(NH3-N)等多种存在形式, 因此实验过程中以溶解态总无机氮(Total dissolved inorganic nitrogen,DIN)形式分析小球藻对氮的吸收效率。即DIN浓度为NO3–-N、NO2–-N、NH3-N三者浓度之和。
2.5 数据处理
小球藻的比生长速率按公式μ= (lnNi+1- lnNi) /(t i+1-ti) 计算, 其中,t i+1和ti表示两次连续取样的时刻点,N i+1和N i分别是微藻在t i+1和ti时刻点的藻细胞数量。氮磷的吸收速率按公式△C=(C i+1-Ci)/ (T i+1-Ti) 计算, 其中,t i+1和ti表示两次连续取样的时刻点,C i+1表示T i+1时刻点的氮磷营养盐浓度,Ci表示Ti时刻点的营养盐浓度。所得数据以单因素和双因素方差分析法(ANOVA)检验不同实验组之间的显著性差异, 显著性水平设置为P<0.05。
将小球藻细胞数量作为因变量(YCell); 初始氮浓度(XN0)、初始磷浓度(XP0)、初始氮磷比(XN/P0)、生长时间(XT), 以及不同取样时间的氮浓度(XNv)、磷浓度(XPv)、氮磷比(XN/Pv)等作为自变量, 进行多元回归分析, 建立回归方程。
3 结果与分析
3.1 蛋白核小球藻的生长
C1和C2组的蛋白核小球藻数量呈持续升高的趋势, 实验结束时平均达到 1.19×107cell·mL–1, 显著高于其他各组(P<0.05); C3、C4、C5组在第6—8 d时达到最高值, 平均为 4.80×106cell·mL–1数量水平, 随后降低至 3.80×106cell·mL–1; C7、C8、C9 三组的生长曲线形态相似, 数量增长幅度较小, 平均仅为 1.52×106cell·mL–1, 远低于C1 和 C2 组(图 1)。就各组的小球藻日平均增长率对比而言, C1和C2组亦显著高于其他各组(P<0.05), 而C8和C9组仍为最低(表1)。
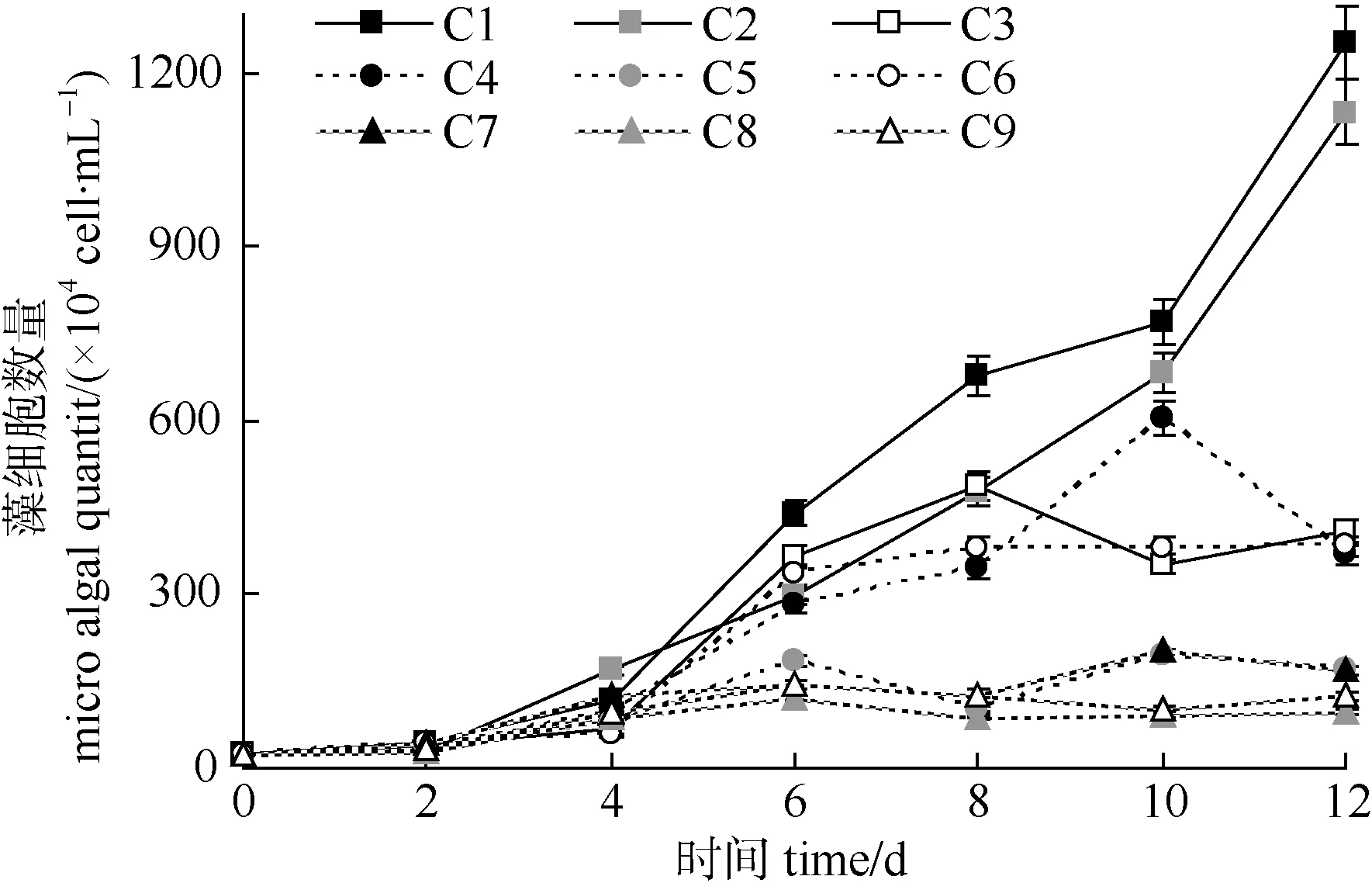
图1 蛋白核小球藻的数量变化Fig.1 The quantity variation of Chlorella pyrenoidosa
3.2 培养液中无机氮的变化
培养液中的DIN浓度总体呈现不断降低的趋势,除C1组外其余各组在第4—6 d时DIN基本耗尽, C1组到12 d时DIN才降低至0 mg·L–1左右。其中初始氮营养浓度越低的试验组其氮耗尽的时间越短,C9组在第2 d时DIN即降至0 mg·L–1, C5组至C8组所需时间则为4 d(图2)。其次, 各组硝酸盐、氨氮和 DIN的最大日吸收量存在一定的差异, 总体而言, 初始氮营养浓度越高, 蛋白核小球藻对硝酸盐、氨氮和 DIN的最大日吸收量也就越高, 反之亦然(图3)。而从培养液中氨氮和硝酸盐完全消耗的速率分析, C1组的氨氮和硝酸盐分别于第8 d和12 d时降至0 mg·L–1, C2组分别为第4 d和12 d,C4组第6 d时两者均降至0 mg·L–1, C3组、C5组、C6组、C7组所需时间为4 d, C9组为2 d。由此可见, 该研究中就小球藻对氮营养吸收的影响因子而言, 氮浓度的影响最大, 其次为氮的化学形式,最后为氮磷比。
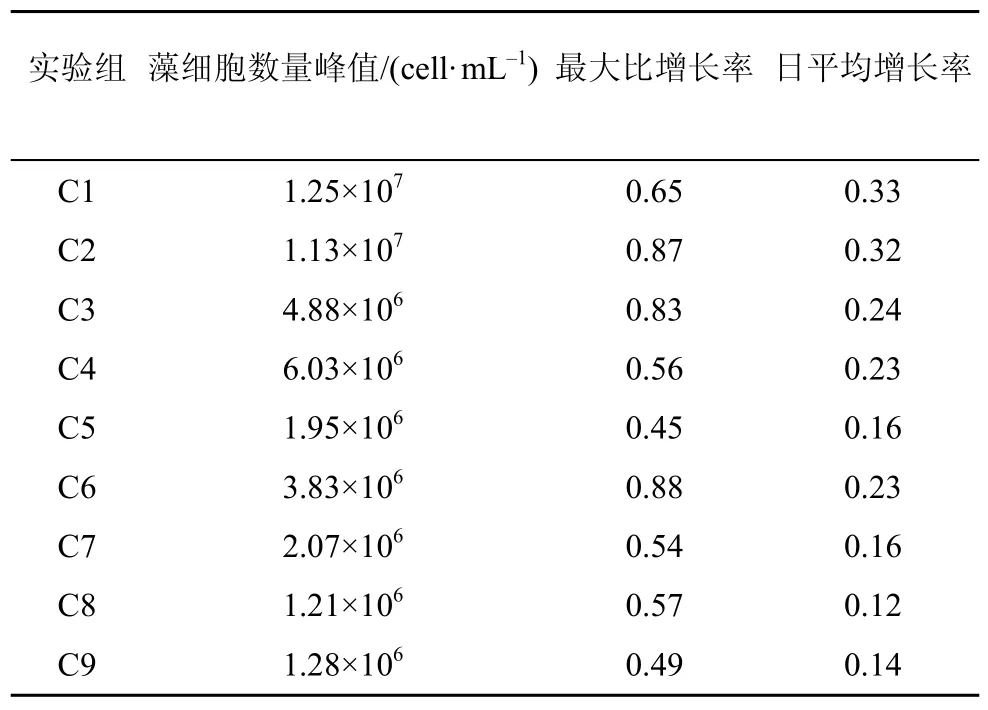
表1 蛋白核小球藻的藻细胞数量峰值、最大比增长率及平均增长率Tab.1 The maximum quantity, maximum growth rate and daily average growth rate of Chlorella pyrenoidosa
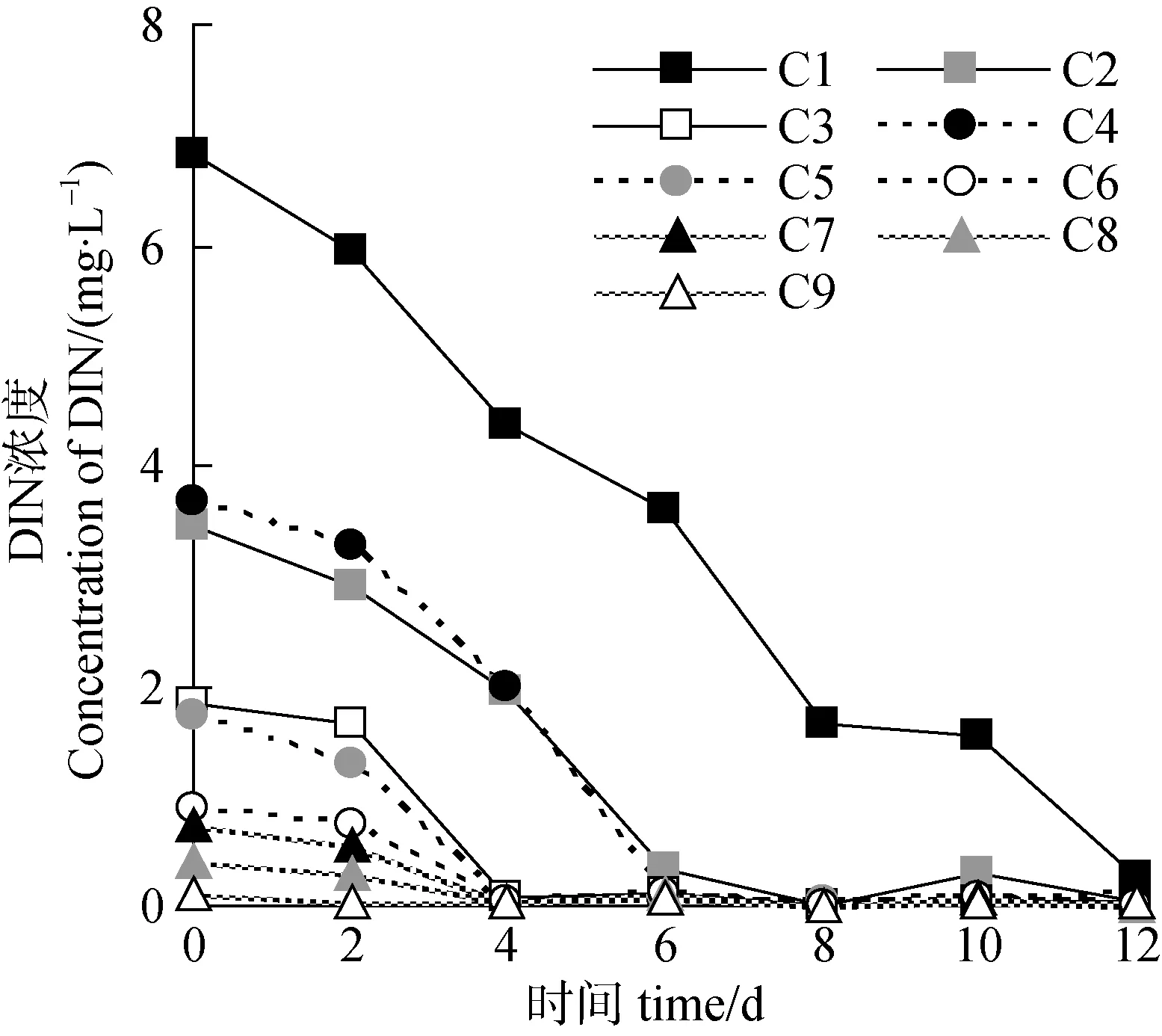
图2 总无机氮的浓度变化Fig.2 The variation of DIN concentration
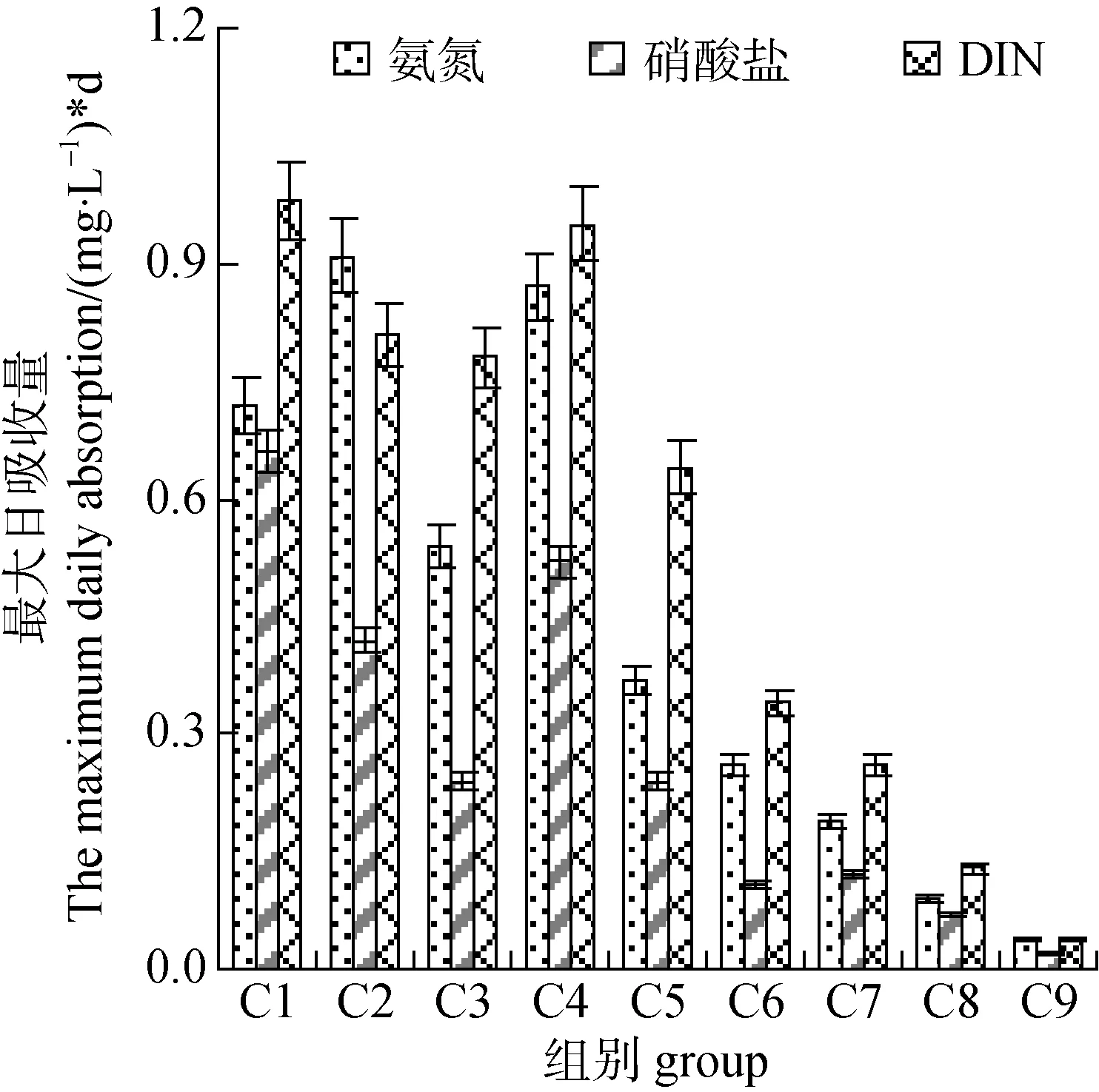
图3 不同氮素的最大日吸收量Fig.3 The maximum nitrogen daily absorption of cell
以上结果显示, 由于低氮磷营养的影响 C7、C8、C9三组的藻细胞生长均受到了明显的限制, 可见, 氮磷营养水平的高低对蛋白核小球藻生长的影响程度远大于N/P; 再者, C1和C2组的藻细胞数量并无显著性差异(P>0.05), C3组的藻细胞数量在实验结束时明显低于C1和C2组(P<0.05), 这表明即使在氮磷富营养条件下, N/P低于8时小球藻的生长仍会受到一定程度的限制, 而当其高于16时N/P不再成为限制性因子。
3.3 培养液中磷的变化
培养液中的磷酸盐浓度总体呈现不断降低的趋势, 第2—8 d时各组的磷酸盐多降低至0 mg·L–1左右(图 4)。按起始磷浓度 0.5 mg·L–1、0.25 mg·L–1、0.05 mg·L–1三个水平分析, 磷的变化曲线基本也分为3个聚类, 其中, 磷浓度较高的C1、C2、C3组的变化趋势基本一致, 培养液中的磷于第8 d时降至 0 mg·L–1左右; 另外 C4、C5、C6 组为一类, C7、C8、C9组为一类, 前者磷被基本完全消耗的时间为4 d, 后者为2 d(图4)。可见, 水体环境中起始磷浓度对微藻的营养吸收速率影响较大。而从其最大日吸收量分析, C1—C6组之间无显著性差异(P>0.05), C7、C8、C9三组则显著低于前六组(P<0.05), 但 C7、C8、C9三组之间无明显差异(P>0.05)。综合以上结果, 该研究中就小球藻对磷酸盐吸收的影响而言, 磷浓度的影响大于氮磷比。
3.4 小球藻生长与氮磷营养指标的相关性分析
将藻细胞数量作为因变量(YCell); 初始氮浓度(XN0)、初始磷浓度(XP0)、初始氮磷比(XN/P0)、生长时间(XT), 以及不同取样时间的氮浓度(XNv)、磷浓度(XPv)、氮磷比(XN/Pv)等作为因变量, 进行多元回归分析, 获得以下回归方程。该方程的相关系数R=0.884,判断系数R2=0.781, 统计量F=27.964, 相伴概率值P<0.001, 表明方程中的多个自变量与因变量藻细胞数之间存在线性回归关系。
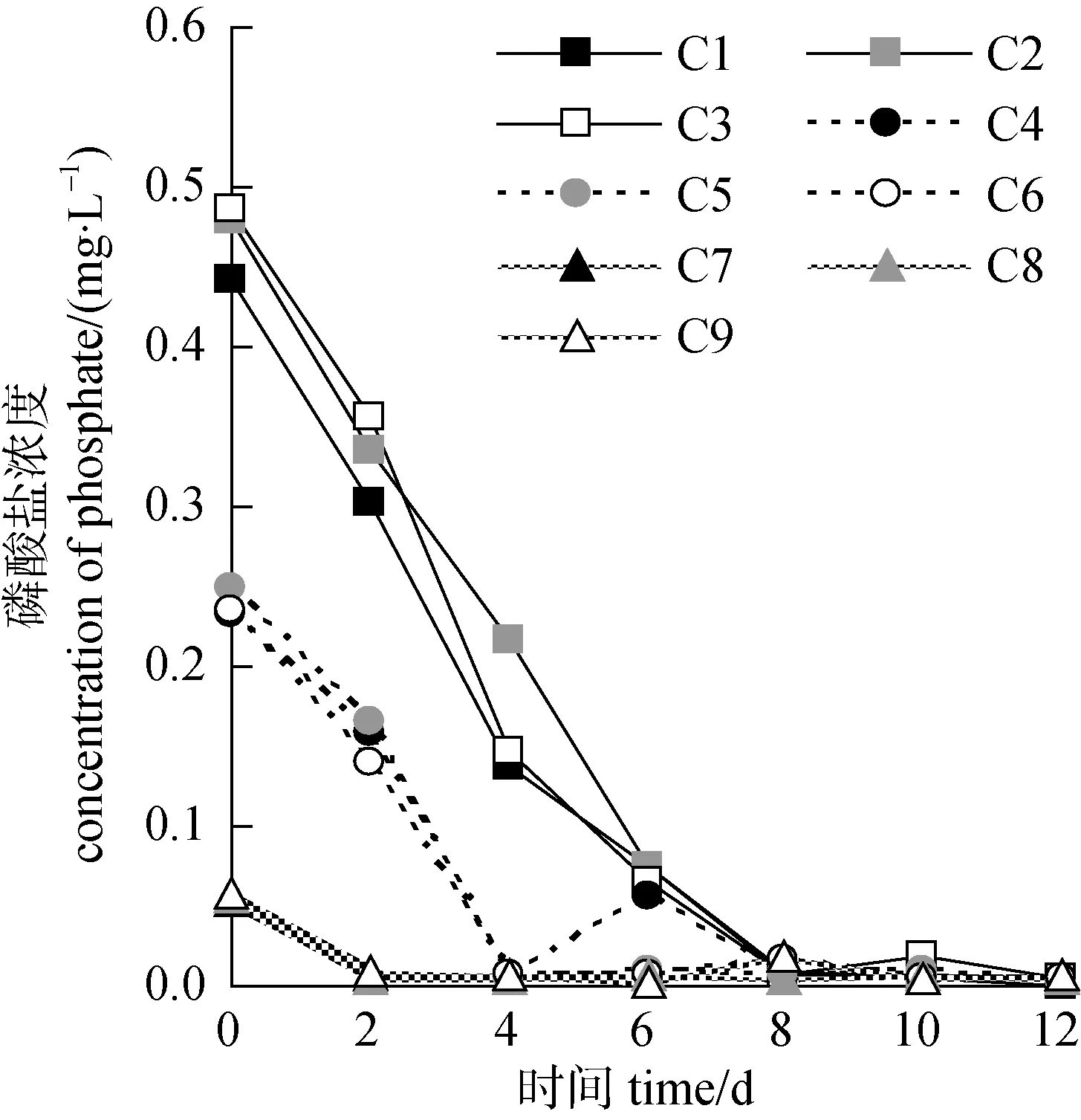
图4 磷酸盐的浓度变化Fig.4 The variation of phosphate concentration
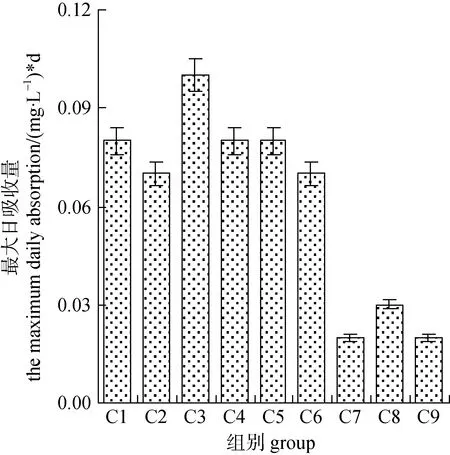
图5 磷酸盐的最大日吸收量Fig.5 The maximum phosphate daily absorption of cell
YCell=138.801XN0+356.516XP0-2.433XN/P0+25.397XT–165.980XNv-119.233XPv+0.649XN/PV-46.692
此外, 结果还显示, 就藻细胞数量与各因子的相关系数而言, 生长时间(XT)的相关性最大, 然后依次为初始氮浓度(XN0)、初始磷浓度(XP0)、不同取样时间的氮磷比(XN/Pv), 其相关系数分别为 0.618、0.462、0.448、0.443。
4 讨论
有学者指出当河流水体的磷高于 0.2 mg·L–1容易造成微藻大量生长甚至造成水华[27]。养殖池塘水体的氮磷富营养化更为明显, 总体随养殖时间的延长, 呈现不同程度的富营养化状况, 有报道显示对虾养殖水体环境中的氮磷总输入为87.8%和 97.9%,而通过养殖生物的氮磷总输出为23.4%和10.5%, 底泥沉积输出 50.5%和 80%[28], 其中大部分的氮磷营养积存于环境中。在集约化对虾高密度养殖池塘水体的无机氮浓度变化范围为 0.375—4.261 mg·L–1,磷酸盐为 0.033—0.514 mg·L–1, 氮磷比为 2—45[28]。而且无机氮中的氨氮和亚硝酸盐对养殖生物也具有较强的毒害作用, 诱发养殖病害导致严重的经济损失[29]。可见, 如何有效运用菌藻生物技术, 促进水环境中的物质循环, 实现水体生态系统的高效自我净化, 具有重要的现实意义[9]。本研究将水体环境中的初始氮磷浓度范围分别设置为0.18—7.20 mg·L–1和0.05—0.50 mg·L–1, N/P 设为 8—32, 基本可反映自然水体的氮磷氮磷营养状况, 具有良好的代表性。
水体环境中氮磷营养的浓度、化学存在形式、不同元素营养的数量比例均会对浮游微藻的数量和优势种组成产生巨大影响[30]。当氮素营养供给不足时会影响微藻细胞氨基酸的合成, 使之生长停滞于静止期[31,32]; 磷缺乏会降低微藻的光合效率和速率, 影响其光合作用的生理机能, 抑制藻细胞的生长与繁殖[33]。Plesnicar指出当水体中的氮达到 2.0 mg·L–1、磷 0.3 mg·L–1时, 有利于延长微藻的生长高峰期[34]; 黄翔鹄认为微绿球藻在氮磷浓度分别达到28.30 mg·L–1和2.076 mg·L–1时, 有利于藻细胞的生长[35]。本研究表明氮磷初始浓度与小球藻的生长关系密切, 高氮磷组的藻细胞增殖数量显著高于其他组, 随着氮磷的消耗藻细胞不断升高,这与孟顺龙等提出的在小球藻和鱼腥藻纯培养体系最大藻细胞数量均随氮磷质量浓度的增加而增加一致[24]。而江怀真等则认为氮磷对小球藻的影响有所区别, 增加氮浓度能提高小球藻生长速率, 磷却无影响[36], 该结果可能与其采用单因子实验设计, 未综合考虑N/P的影响有关。通过科学地施加氮磷混合肥, 增加水体氮磷浓度可有效促进各种微藻的生长繁殖[37,38], 但同时也应考虑 N/P的影响。Alonso指出一般养殖水体的 N/P变动幅度较大, 其中集约化养殖池塘水体的N/P变动范围为5.5—67.0, 半集约化养殖水体为1.1—61.0[39], 一般N/P值较高时有利于绿藻、硅藻的生长, 反之则利于促进蓝藻的增殖。本研究结果表明在氮磷富营养条件下, N/P低于8时小球藻的生长仍会受到一定程度的限制, 而当其高于16时N/P不再产生影响, 这与上述观点基本相同。对此, 有学者指出通过选择性施肥, 定向调节水体氮磷比可有效控制蓝藻的生长[23]。而 Wang等报道氮磷比对纯培养的小球藻的生长没有影响[40],这可能与其将磷浓度设置在0.025 mg·L–1的低水平,仅通过调节氮浓度设定不同氮磷比的设计有关。本研究结果显示, 蛋白核小球藻与氮磷浓度, 以及不同取样时间的氮磷浓度等多个因子均存在明显线性关系, 且氮磷浓度对藻细胞的影响明显大于N/P(P<0.05)。所以, 在定向调控水体氮磷营养时既需要考虑氮磷的浓度高低情况, 还需适当控制其数量比例关系。基于本研究结果而言, 建议在养殖过程中定期监测水体溶解性氮磷营养盐的动态变化,通过科学的人为干预将水体N/P设定在大于16的数量水平; 同时, 考虑到池塘中磷的输出主要以底泥沉积吸附的方式为主, 水中磷酸盐的浓度相对缺乏,而饲料中的氮输入相对丰盈, 故而在养殖过程中可根据微藻生长状况, 每7到12天定期使用解磷菌或适量添加水溶性磷肥提高水体磷营养的有效供给,从而为维持水体中微藻的良好生长提供有力的氮磷营养条件。
总体而言, 以往的报道多把氮磷浓度、氮磷比对微藻的影响进行独立式研究, 以期通过单因子的营养需求分析阐释究竟何种氮磷因子对微藻的影响贡献更大, 这在一定程度上忽略了自然水体环境下,氮磷的浓度与比例其实均客观影响着微藻的生长、群落结构、优势种演替动态以及群落的生态功能等。所以, 研究过程中应充分考虑两者的协同效应, 同时, 将氮磷营养消耗的动态变化特性亦纳入分析过程, 才能获得更为科学的结果。
[1]刘孝竹, 曹煜成, 李卓佳, 等.高位虾池养殖后期浮游微藻群落结构特征[J].渔业科学进展, 2011, 32(3): 84–91.
[2]彭聪聪, 李卓佳, 曹煜成, 等.凡纳滨对虾半集约化养殖池塘浮游微藻优势种变动规律及其对养殖环境的影响[J].海洋环境科学, 2011, 30(2): 193–198.
[3]彭聪聪, 李卓佳, 曹煜成, 等.珠江河口区罗非鱼养殖池塘中浮游微藻的群落特征[J].农业环境科学学报, 2012,1(9): 2019–2030.
[4]刘孝竹, 李卓佳, 曹煜成, 等.低盐度养殖池塘常见浮游微藻的种类组成、数量及优势种群变动[J].南方水产,2009, 5(1): 9–16.
[5]李卓佳, 梁伟峰, 陈素文, 等.虾池常见微藻的光照强度、温度和盐度适应性[J].生态学杂志, 2008, 27(3): 397–400.
[6]朱建新, 曲克明, 刘慧, 等.小球藻和光合细菌在大菱鲆育苗中对水质调节作用的研究[J].海洋水产研究, 2008,29(6): 116–121.
[7]CAO YC, WEN GL, LI ZJ, et al.The effects of dominant microalgae species and bacterial quantity on shrimp production in final culture season[J].Journal of Applied Phycology, 2014, 26(4): 1749–1757.
[8]查广才, 周昌清, 黄建容, 等.凡纳对虾淡化养殖虾池微型浮游生物群落及多样性[J].生态学报, 2004, 24(8):1752–1759.
[9]曹煜成, 李卓佳, 杨莺莺, 等.浮游微藻生态调控技术在对虾养殖应用中的研究进展[J].南方水产, 2007, 3(4):70–73.
[10]JI Y, HU W, LI X, et al.Mixotrophic growth and biochemical analysis ofChlorella vulgariscultivated with diluted monosodium glutamate wastewater [J].Bioresource Technology, 2014, 152: 471–476.
[11]HUANG Y, HONG A, ZHANG D, et al.Comparison of cell rupturing by ozonation and ultrasonication for algal lipid extraction fromChlorella vulgaris[J].Environmental Technology, 2014, 35(8): 931–937.
[12]LANANAN F, JUSOH A, ALI N, et al.Effect of Conway Medium and f/2 Medium on the growth of six genera of South China Sea marine microalgae [J].Bioresource Technology, 2013, 141(4): 75–82.
[13]SHIH CH, CHEN HY, LEE HC, et al.Purple chromoprotein gene serves as a new selection marker for transgenesis of the microalgaNannochloropsis oculata[J].PLoS One, 2015, 10(3): e0120780.
[14]ROOPNARAIN A, GRAY VM, SYM SD.Phosphorus limitation and starvation effects on cell growth and lipid accumulation inIsochrysis galbanaU4 for biodiesel production[J].Bioresource Technology, 2014, 156(2):408–411.
[15]VILLAY A, LAROCHE C, RORIZ D, et al.Optimisation of culture parameters for exopolysaccharides production by the microalgaRhodellaviolacea[J]. Bioresource Technology, 2013, 146(10): 732–735.
[16]RHEE GY.Effects of N: P atomic ratios and nitrate limitation on aglae growth, cell composition and nitrate uptake[J].Limnology and Oceanography, 1978, 23(1):10–25.
[17]TERRY KL, LAWS EA, BURNS DJ.Growth rate variation in the N: P requirement ratio of phytoplankton[J].Journal of Phycology, 1985, 21(2): 323–329.
[18]张青田, 张兆琪, 董双林.氮、磷对金藻的增殖效应[J].海洋湖沼通报, 2002(2): 45–51.
[19]李艳和, 管远亮, 叶应旺, 等.2种培养液培养蛋白核小球藻的效果[J].安徽农业科学, 2003, 31(2): 262–263.
[20]黄翔鹄, 李长玲, 刘楚吾, 等.微绿球藻对氮和磷营养盐需求的研究[J].海洋科学, 2002 , 26(8): 13–17.
[21]梁伟峰, 陈素文, 李卓佳, 等.虾池常见微藻种群温度、盐度、和氮、磷含量生态位[J].应用生态学报, 2009, 20(1):223–227.
[22]王丽卿, 黄旭雄.不同营养盐浓度下微绿球藻的生长及水体中氮磷的变化[J].上海水产大学学报, 2002, 11(3):215–218.
[23]万蕾, 朱伟, 赵联芳.氮磷对微囊藻和栅藻生长及竞争的影响[J].环境科学, 2007, 28(6): 1230–1235.
[24]孟顺龙, 王菁, 裘丽萍, 等.氮磷质量浓度对普通小球藻和鱼腥藻生长竞争的影响[J].生态环境学报.2015, 24(4):658–664.
[25]胡晓娟, 李卓佳, 曹煜成, 等.养殖池塘生态系统中磷的收支及解磷微生物的研究进展[J].安全与环境学报,2010, 10(1): 7–11.
[26]GB 17378.4–2007, 海洋监测规范[S].北京: 中国标准出版社, 2008.
[27]黄翔鹄, 李长玲, 郑莲, 等.固定化微藻对虾池弧菌数量动态的影响[J].水生生物学报, 2005, 29(6): 684–688.
[28]李卓佳, 虞为, 朱长波, 等.对虾单养和对虾–罗非鱼混养试验围隔氮磷收支的研究[J].安全与环境学报, 2012,12(4): 50–55.
[29]张华军, 李卓佳, 张家松, 等.凡纳滨对虾免疫指标变化与其养殖环境理化因子的关系[J].大连海洋大学学报,2011, 26(4): 356–361.
[30]李卓佳, 李奕雯, 曹煜成, 等.对虾养殖环境中浮游微藻、细菌及水质的关系[J].广东海洋大学学报, 2009,29(4): 95–98.
[31]GRASKE J, VANDER MOLEN M.Effects of nutrient concentration on the growth of Tahitian Isochrysis[J].Austasia Aquaculture, 1992, 6: 43–47.
[32]RENAUD SM, PARRY DL, LUONG VAN T, et al.Effect of light intensity on the proximate biochemical and fatty acid composition ofIsochrysis sp.andNannochloropsis oculatafor use in tropical aquaculture [J].Journal of Applied Phycology, 1991, 3(1): 43–53.
[33]FABREGAS J, ABALDE J, HERRERO C.Biochemical conposition and growth of the marine microalgaDunaliella tertiolecta(Butcher) with different ammonium nitrogen concentrations as chloride, sulphate, nitrate and carbonate[J].Aquaculture, 1989, 83(3): 289–304.
[34]PLESNICAR M, KASTORI R, PETROVIC N, et al.Photosynthesis and chlorophyll fluorescence in sunflower(Helianthus annuusL.) leaves as affected by phosphorus nutrition[J].Journal of Experimental Botany, 1994, 45(7):919–924.
[35]王丽花, 曹煜成, 李卓佳.溶藻细菌控藻作用及其在对虾养殖池塘中的应用前景[J].南方水产科学, 2012, 8(4):76–82.
[36]江怀真, 张维, 刘天中, 等.氮、磷浓度对小球藻生长及油脂积累的影响[J].食品工业科技.2011, 32(6):204–207.
[37]王海雷, 赵文, 李晓东, 等.施肥对多盐水池塘浮游植物演替的影响[J].中国水产科学, 2005, 12(5): 608–613.
[38]彭聪聪, 李卓佳, 曹煜成, 等.虾池浮游微藻与养殖水环境调控的研究概况[J].南方水产, 2010, 6(5): 74–80.
[39]ALONSO R R, PAEZ O F.Nutrients, phytoplankton and harmful algal blooms in shrimp ponds: a review with special reference to the situation in the Gulf of California[J].Aquaculture, 2003, 219(1/4): 317–336.
[40]WANG J, QIU L P, MENG S L, et al.Influences of Nitrogen-phosphorus ratio on the growth and competition ofChlorella vulgaandAnabaena sp.strain PCC[J].Agricultural Science and Technology, 2015, 16(8):1757–1762.
Effects of phosphorus concentration and nitrogen-phosphorus ratio on absorption of nitrogen and phosphorus byChlorella pyrenoidosa
CAO Yucheng1, LI Zhuojia1, HU Xiaojuan1, XU Yu1, SUN Zhiwei1, LI Yiwen2, WEN Guoliang1,*
1.South China Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences, Key Laboratory of South China Sea Fishery Resources Exploitation and Utilization, Ministry of Agriculture, Key Laboratory of Fishery Ecology and Environment,Guangzhou510300,China2.Guangdong Ocean and Fishery Publishing Media Company Limited,Guangzhou510220,China
The study investigated the absorption characteristics of nitrogen (N) and phosphorus (P) byChlorella pyrenoidosaunder the synergistic effects of different nitrogen-phosphorus ratios (N/P) and concentrations of P. The P concentrations of microalgal culture media were 0.50 mg·L–1, 0.25 mg·L–1, 0.05 mg·L–1and the N/P ratios were 32, 16, 8, respectively. There were nine test groups with triplicate in each group, and the test was lasted for 12 days. The results showed that the quantity ofC. pyrenoidosawas significantly lower at low P and N concentrations than that at high concentrations (P<0.05), and the N/P ratio negatively affected the growth ofC. pyrenoidosaas it was lower than 8 at high P and N concentrations, while the effect would relieve as the ratio was higher than 16. The multiple regression analysis showed that there were significant linear relationships betweenC. pyrenoidosaquantity with the initial concentrations of N and P, the influence of which concentrations also were much higher than other factors (P<0.05). Accordingly, though the growth ofC. pyrenoidosapositively correlated to the assimilation of N and P, it is still difficult to improve the absorption efficiency of P and N byC.pyrenoidosajust through regulating the N/P ratios in aquaculture water.
phosphorus; nitrogen-phosphorus ratio;Chlorella pyrenoidosa; absorption
10.14108/j.cnki.1008-8873.2017.05.005
X172; S917.1
A
1008-8873(2017)05-034-07
曹煜成, 李卓佳, 胡晓娟, 等.磷浓度与氮磷比对蛋白核小球藻氮磷吸收效应的影响[J].生态科学, 2017, 36(5): 34-40.
CAO Yucheng, LI Zhuojia, HU Xiaojuan, et al.Effects of phosphorus concentration and nitrogen-phosphorus ratio on absorption of nitrogen and phosphorus byChlorella pyrenoidosa[J].Ecological Science, 2017, 36(5): 34-40.
2017-04-11;
2017-06-02
中国水产科学研究院基本科研业务费(2017HY-ZD0501); 现代农业(虾蟹)产业技术体系专项(CARS-48); 广东省海洋渔业科技与产业发展专项(A201401B03); 广东省科技计划项目(2016A040402034)
曹煜成(1979—), 男, 博士, 副研究员, 从事对虾健康养殖及池塘环境调控研究, E-mail:cyc_715@163.com
*通信作者:文国樑(1978—), 男, 副研究员, 主要从事养殖水环境调控与修复研究, E-mail:guowen66@163.com
