五大连池矮曲林土壤微生物量碳、氮对季节变化的动态响应特征
2017-11-10姜明月张文天魏晓雪
姜明月, 张文天, 魏晓雪
1.黑龙江省科学院火山与矿泉研究所, 哈尔滨 150080 2.黑龙江省科学院高技术研究院, 哈尔滨 150001
五大连池矮曲林土壤微生物量碳、氮对季节变化的动态响应特征
姜明月1,2,*, 张文天1,2, 魏晓雪1,2
1.黑龙江省科学院火山与矿泉研究所, 哈尔滨 150080 2.黑龙江省科学院高技术研究院, 哈尔滨 150001
为今后更好了解矮曲林对五大连池风景区生态系统的影响, 比较老黑山主要矮曲树种山杨、香杨和白桦的土壤微生物量的差异特征, 揭示其对微生物生物量碳(MBC)、微生物生物量氮(MBN)的影响, 选取了老黑山脚下熔岩台地15个样点进行研究。结果表明: 矮曲林三种主要林型的土壤微生物生物量碳氮季节变化规律相似, 全年整体含量为微生物量碳香杨<白桦<山杨, 微生物量氮则是山杨<白桦<香杨。白桦矮曲林土壤微生物量碳与其生物量氮呈显著相关性(p<0.05), 与土壤全氮呈现显著相关性(p<0.05)。山杨矮曲林土壤微生物量碳与其土样有机碳和全氮呈高度负相关性(p<0.05)。香杨矮曲林土壤微生物生物量碳与其土样有机碳呈高度负相关性(p<0.05), 与土壤全氮呈显著负相关性(p<0.05)。矮曲林土壤含水率四季变化不大, 涵养水源能力差, 微生物生物量呈现季节性动态变化, 由此可见, 不同植被型对土壤微生物量碳和氮有一定影响, 而含水率对其影响不大, 温度变化对其有一定影响。
五大连池; 火山; 矮曲林; 土壤微生物生物量
1 前言
土壤微生物是联系凋落物分解、养分元素C和N等的矿化及土壤与大气C和N的交换等过程的关键, 其生长活性对森林生态过程具有重要的调控作用[1]。它是森林生态系统中的重要分解者改善土壤健康状况, 利于植物生长和发育[2-4]。土壤微生物量可反映土壤同化和矿化能力的大小, 其参与生态系统养分循环、有机质分解等生态过程, 在陆地生态系统碳氮循环中发挥着重要作用[5-6]。土壤微生物数量和生物量受气候、土壤和植被因子的显著影响,即使在相同的气候条件和土壤类型下, 不同植被下的土壤微生物仍然存在较大差异[7]。另外, 土壤微生物量的季节变化与环境因子以及植物生理生态特性息息相关[8]。水热条件的季节变化影响土壤微生物量的周转、养分的有效性以及植物对养分的利用[9]。国内目前对土壤微生物研究集中在土壤微生物量碳氮含量和有机质动态关系[10-11]、不同生态系统土壤微生物生物量比较[12]等方面。水热要素变化对土壤微生物生物量影响也有研究报道[13-14], 但都以室内培养土壤或通过季节差异比较分析来确定温湿度变化对土壤微生物生物量的影响, 田间条件下温度和降水量变化对微生物量关系方面的研究却不多。
五大连池-世界著名地质公园, 位于中国东北部,气候条件恶劣, 地质变化丰富。老黑山为本地新期火山的典型代表, 岩浆流经形成的熔岩台地土壤十分贫瘠, 故而植物只有改变自身形态才能在这种特殊的生境中生存, 矮曲林是本地区植物进化的突出范例[15], 尤其以香杨、山杨和白桦的矮曲程度最为显著。矮曲林作为老黑山生态系统重要组成部分,影响着火山生态恢复的进程, 而矮曲林土壤微生物方面的研究当前还是罕见报导的。本文选择对矮曲林表层土壤微生物进行研究, 发现不同生境下生物量碳、氮含量及其季节变化规律, 探讨影响土壤微生物生物量碳氮季节变化的主要环境因子, 为探索本地区熔岩台地这一特殊的生态系统变化提供理论支撑, 为本地区环境保护提供依据。
2 研究区概况与研究方法
2.1 研究地点概况
五大连池风景区是中国最典型和最完整的近代火山喷发遗迹, 已获得“世界地质公园”、“世界生物圈保护区”、“中国矿泉城”等称号, 位于黑龙江省北部地区, 地理坐标为北纬 48°30′—48°51′、东经126°00′—126°25′, 属于寒温带大陆性季风气候, 低温冷湿, 年平均气温0 ℃左右, 年平均降水量在500毫米, 无霜期119天。
五大连池风景区最近一次火山爆发而形成的老黑山和火烧山, 喷发于 1719—1721年, 距今仅 270年, 地质地貌保存完好, 熔岩流动景象清晰, 地貌复杂多样, 形态各异, 面积达 64平方公里, 奇丽壮观。熔岩台地生境特殊, 近年来受人为因素影响越来越大, 了解此处微生物生物量的变化, 有利于预警此处的生态变化、保护环境。
2.2 研究方法
2.2.1 样品的采集
本研究的样地设置在老黑山脚下, 以熔岩台地山杨、香杨和白桦三种主要林型为研究对象, 每种森林类型设置5块固定样地, 每块样地间相隔10 m以上。本次试验采样时间是2015年3、6、9、11月,采样时, 先去除地表面凋落物, 采得0—10 cm土层,然后将每种林型采集的 5个样进行混和后, 及时装入自封袋带回实验室, 取部分放入冰箱4 ℃下保存,供土壤微生物生物量的测定, 剩余部分风干后过 2 mm筛, 用于土壤指标测试。
2.2.2 土壤有机质及全氮的测定
土样基本理化性质的测定用常规分析方法[16]:土壤有机质测定采用重铬酸钾容量法—外加热法,土壤全氮测定采用半微量凯氏法。
土壤有机质(g·kg-1)=C·5·(V0-V) ·10-3·3.0·1.1·103·(m·V0)-1式中:C为0.8 mol·L-1(1/6 K2Cr2O7)标准液的浓度; 5为重铬酸钾标准液加入的体积(mL);V0为滴定空白所用FeSO4体积(mL);V为滴定样品所用FeSO4体积(mL);3.0为四分之一碳原子的摩尔质量(g·mol-1); 10-3将mL转化为L; 1.1为氧化校正系数;m为风干土质量。
土壤全氮(g·kg-1)=C·14.0·(V0-V) ·10-3·103·m-1式中:C为0.01 mol·L-1(1/2 H2SO4)标准溶液的浓度;V0为滴定空白所用酸体积(mL);V为滴定样品所用酸体积(mL); 14.0为氮原子的摩尔质量(g·mol-1); 10-3将mL转化为L;m为风干土质量。
2.2.3 土壤微生物生物量碳和氮的测定
土壤微生物量C、N采用氯仿熏蒸浸提法测定[17]。提取液中C的测定用重铬酸钾-硫酸消煮, 硫酸亚铁滴定法, N的测定采用凯氏消煮法。微生物量C、N的计算, 见公式(1)、(2)。

式中:EC、EN分别表示熏蒸与未熏蒸土壤提取液中所测C、N含量之差,KEC、KEN分别表示微生物量C、N浸提测定的比例, 数值分别为0.38、0.54。
微生物生物量商(%)用公式: qMB=MBC·SOC-1
式中,MBC为土壤微生物量碳,SOC为土壤有机碳。
2.2.4 统计分析
试验数据的处理和运算采用 Excel 2010 软件与SPSS 19.0统计软件, 相关性分析用Pearson 相关分析,检验相同条件下土壤微生物碳氮生物量的相关性。
3 结果与分析
3.1 三种林型土壤理化性质比较
如表1所示, 老黑山熔岩台地的白桦矮曲林土壤有机碳呈现连续上升趋势, 而土壤全氮含量第一季度呈现最高值, 在第二季度急剧下降, 三、四季度呈现缓慢上升趋势。山杨矮曲林土壤有机碳及全氮变化趋势相似, 均在第二季度出现一个峰值, 不同的是土壤全氮的含量在第四季度超出前三个季度。香杨矮曲林土壤有机碳及全氮第一季度值为最低,第二季度出现峰值, 第三季度为转折, 继续在第四季度上升。三处林地含水率在第二季度达到最高,全年平均在1%—2%之间。
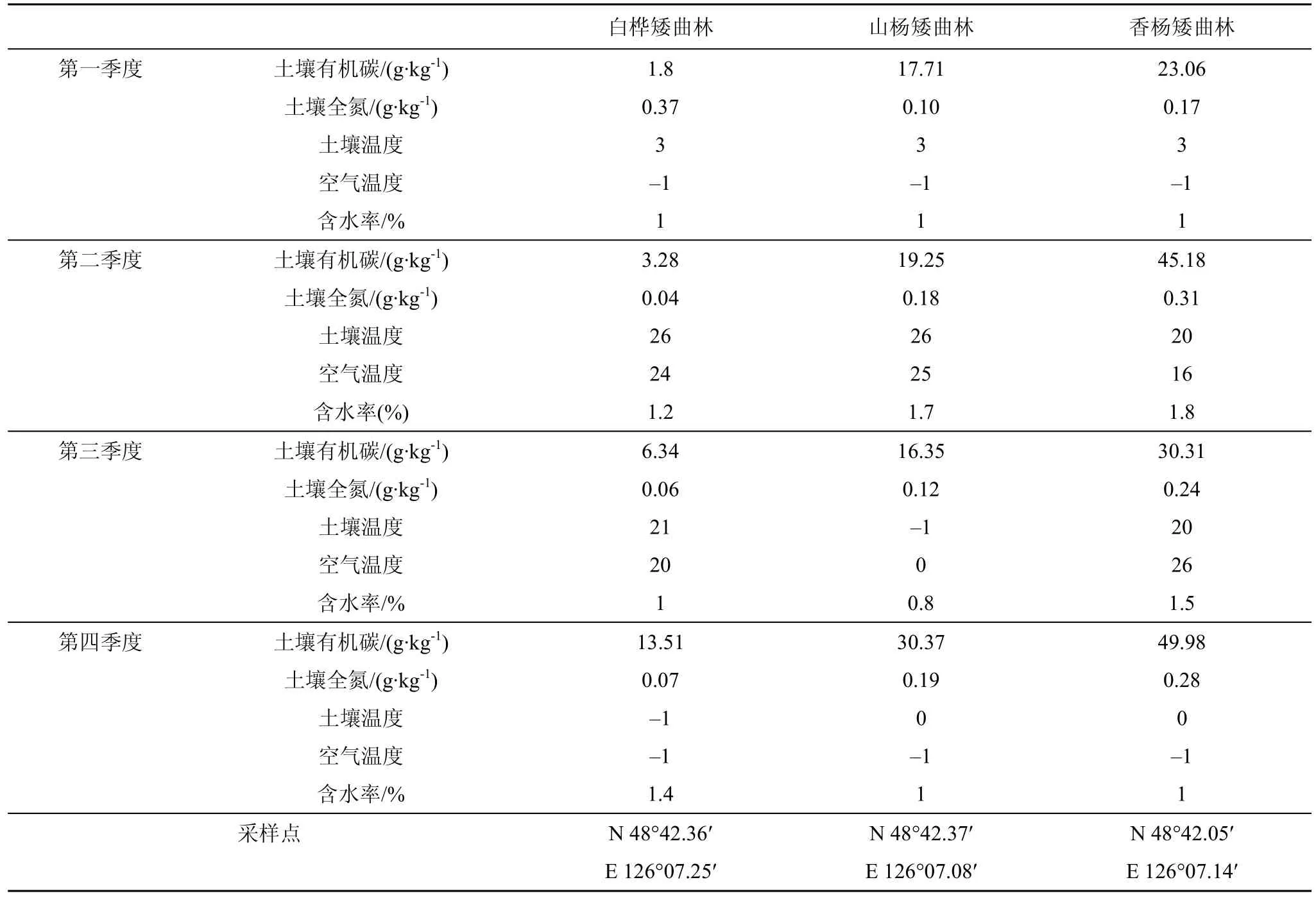
表1 矮曲林土壤理化性质季节变化Tab.1 The physical and chemical properties of elfin forest soil at different seasons
在我们的研究过程中发现, 矮曲林地土壤有机碳及全氮整体含量低于本地区未经过火山喷发后熔岩流经的普通林地, 三种林地矮曲林土壤有机碳全年均值依次为 6.23g·kg-1、20.92g·kg-1和 37.13g·kg-1,而对应的、未经过火山喷发的林地土壤有机碳全年均值依次为 66.21g·kg-1、35.20g·kg-1和 51.01g·kg-1。相关研究表明, 黑土地土壤有机碳含量为 50—100 g·kg-1, 本地区非矮曲林的土壤有机碳基本达到这一范围, 而熔岩台地的矮曲林则普遍低于这一范围很多。本研究中, 三种矮曲林的土壤全氮全年均值分别为 0.135g·kg-1, 0.148g·kg-1和 0.25g·kg-1, 对应的、未经过火山喷发的林地土壤全氮全年均值分别为2.48g·kg-1, 0.88g·kg-1和 1.85g·kg-1, 几乎是矮曲林地土壤全氮的二倍。老黑山约在300年前喷发, 熔岩流经的地区生境恢复缓慢, 土壤含量低, 多数是火山灰与土壤的混合体, 涵养水源能力差, 全年土壤含水率均低于2%, 供给生物生长的营养成分相对较少。
有学者对黑龙江帽儿山、北京东灵山等十二处有地域特点的森林土壤进行研究发现: 不同地点表层土壤有机碳平均含量为 38.79 g·kg-1, 其中樟木镇有机碳平均含量最大为 94.7 g·kg-1, 泰和含量最低为11.25 g·kg-1; 不同地点间土壤全氮含量有明显差异, 其中樟木镇最高为6.72 g·kg-1, 黑龙江帽儿山为 4.02 g·kg-1, 其他地点的全氮含量在 2.0 g·kg-1以下, 泰和的土壤全氮含量最低为0.86 g·kg-1[18]。然而,老黑山白桦矮曲林的土壤有机碳含量比含量最低的江西泰和还要少近二分之一, 而西藏樟木镇土壤有机碳含量是香杨矮曲林的近三倍左右; 三种主要矮曲林土壤全氮整体要较其余地点的含量相差较大,含量最低的泰和土壤全氮是含量最高的香杨矮曲林的近3.5倍。如此差别可能是因为火山喷发后, 熔岩流经处土壤不存, 经过三百年的恢复, 成土还是相当的少, 养分含量较低, 熔岩台地上的树林成长形态矮曲, 与此有很大关系。
3.2 三种林型土壤微生物生物量碳、氮动态研究
3.2.1 三种林型土壤微生物生物量碳、氮季节规律
如图1所示, 矮曲林土壤微生物生物量碳季节变化规律相似, 三种主要矮曲林均在第二季度含量呈现波谷值, 分别是白桦矮曲林 74.74 μg·kg-1、山杨矮曲林 779.53 μg·kg-1和香杨矮曲林 207.87 μg·kg-1,第三季度呈现峰值, 分别是白桦矮曲林616.58 μg·kg-1、山杨矮曲林1556.61 μg·kg-1和香杨矮曲林445.08 μg·kg-1。香杨矮曲林土壤微生物生物量碳含量整体明显较山杨矮曲林和白桦矮曲林土壤微生物生物量碳低, 山杨矮曲林土壤微生物生物量碳整体较其他两种林型高。三种矮曲林第一、三季度生物量碳含量高于第二、四季度, 除第二季度外, 微生物量碳的水平为山杨矮曲林>白桦矮曲林>香杨矮曲林; 全年水平看,山杨矮曲林土壤微生物生物量碳浮动比其他两种林型剧烈, 这与林地所处坡向、不同林型提供的底物、土壤理化性质和植被生长状况均有关系。
如图2所示, 三种矮曲林土壤土壤微生物生物量氮季节变化整体呈“对号”趋势。均在第二季度出现波谷值, 然后随时间的持续而缓慢递增, 第四季度达到含量最高值, 白桦矮曲林与香杨矮曲林第一高于第三季度生物量氮。香杨矮曲林土壤微生物生物量氮全年波动最为明显、剧烈, 在第二季度迅速下降到1.52 μg·kg-1, 而第四季度急剧上升至17.47 μg·kg-1, 均是三种矮曲林微生物量氮的最值。白桦矮曲林土壤微生物生物量氮整体含量比其他两种高,含量在 4.95 μg·kg-1—10.11 μg·kg-1之间浮动。山杨矮曲林土壤微生物生物量氮整体含量最低, 含量范围是 2.33 μg·kg-1—4.54 μg·kg-1。
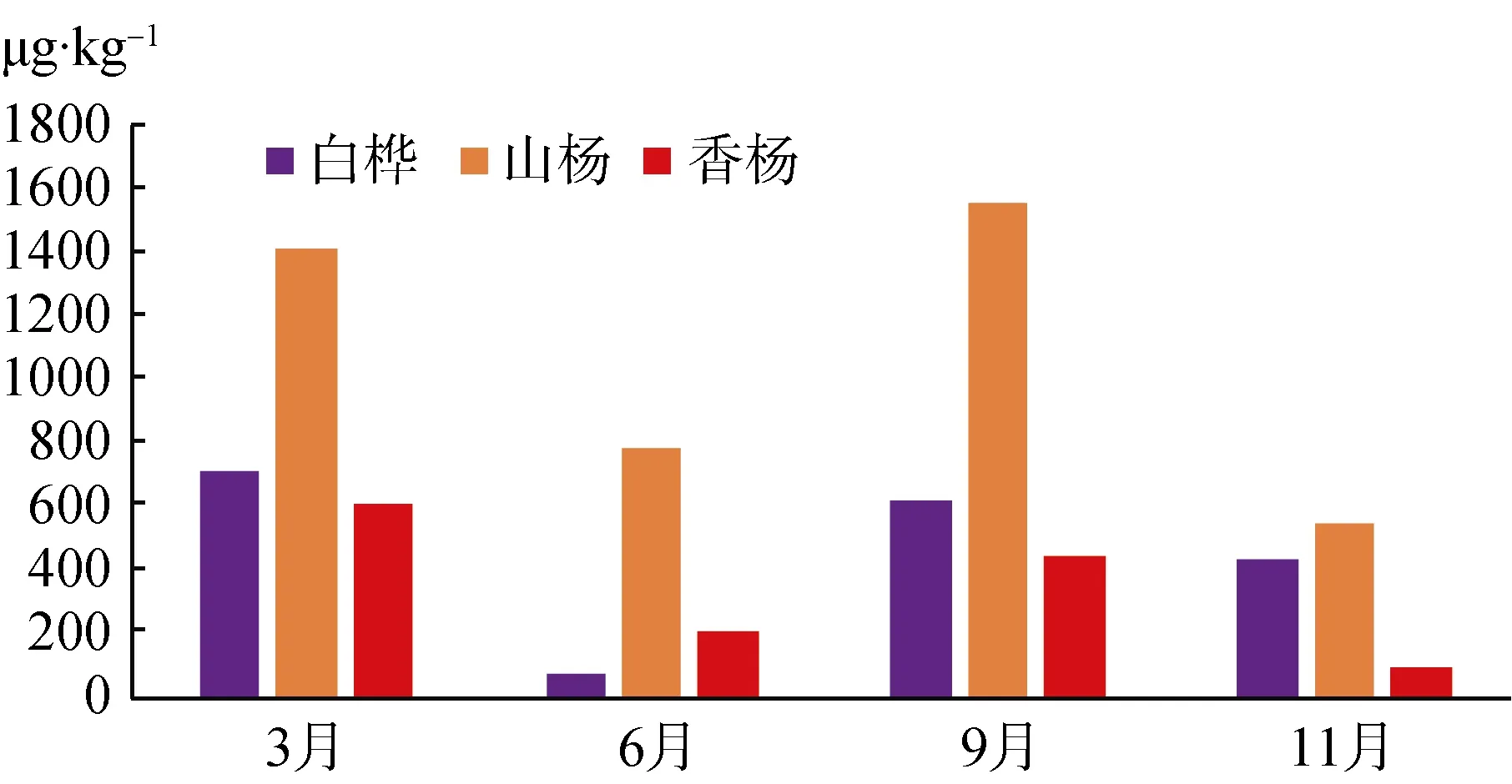
图1 五大连池风景区矮曲林土壤微生物生物量碳季节变化Fig.1 The seasonal variation of soil microbial biomass carbon in the elfin forest of Wudalianchi Scenic
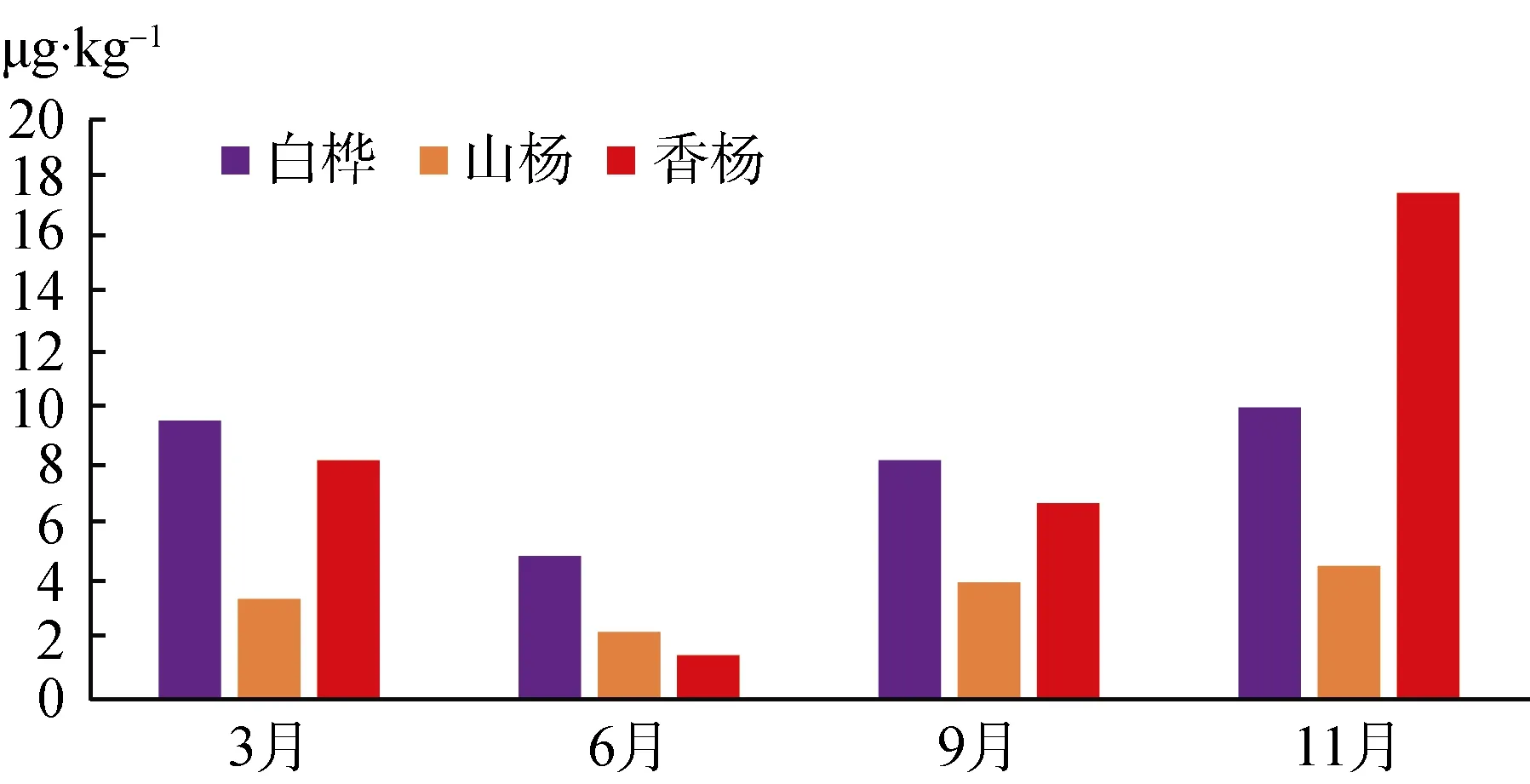
图2 五大连池风景区矮曲林土壤微生物生物量氮季节变化Fig.2 The seasonal variation of soil microbial biomass nitrogen in the elfin forest of Wudalianchi Scenic
白桦矮曲林土壤有机碳含量呈现递增状态, 而微生物量碳与其趋势明显不同, 呈现第二和四季度下降、第三季度上升; 土壤全氮和微生物量氮均为第二季度下降, 三四季度连续上升趋势。山杨和香杨矮曲林的土壤有机碳和全氮呈现第二四季度上升,三季度下降, 然而与之对应的微生物量碳趋势相反,微生物量氮则呈现三四季度连续上升趋势。白桦矮曲林明显与山杨和香杨矮曲林微生物量碳氮和土壤有机碳及全氮的季度变化规律不同, 这可能与不同植被的对土壤利用的方式不同有关。
3.2.2 土壤微生物量商
矮曲林土壤微生物量碳含量十分微小, 几乎是其他森林土壤微生物量碳的千分之一, 因而土壤微生物量商也十分低。图3是土壤微生物量商的季节变化, 与土壤微生物量碳趋势相似, 白桦矮曲林的变化相对剧烈, 香杨矮曲林季节波动较为平缓, 三种林型土壤微生物量商在0.00019—0.0396156之间变化, 整体含量为香杨<山杨<白桦的趋势。
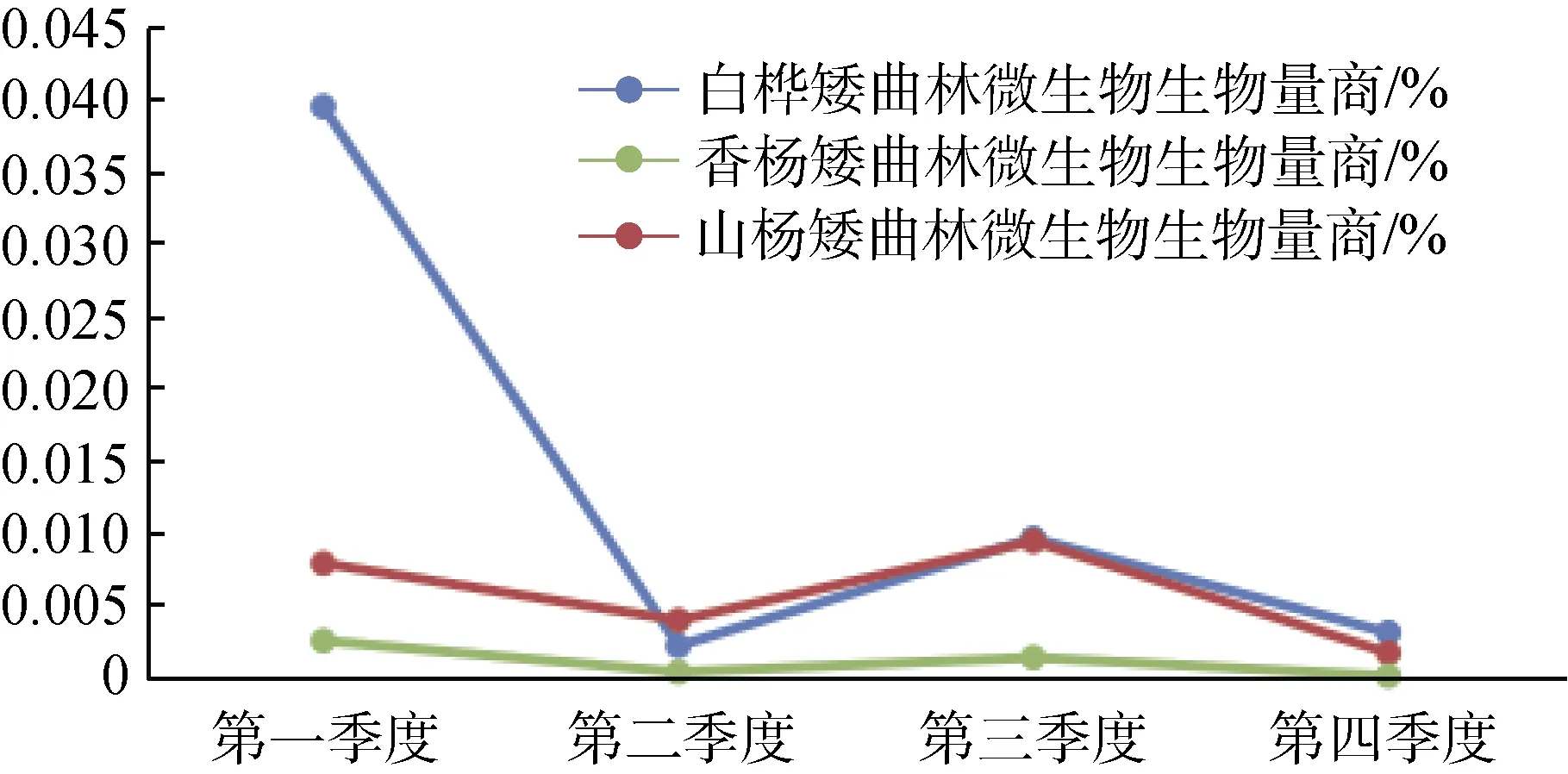
图3 五大连池风景区矮曲林土壤微生物量商季节变化Fig.3 The seasonal variation of soil microbial biomass and metabolic quotient in the elfin forest of Wudalianchi Scenic
3.3 三种林型土壤微生物生物量碳氮及土壤有机碳、全氮之间的关系
表2给出的 Pearson相关性系数可知(P<0.05),显著相关性, 与山杨、香杨矮曲林土壤微生物量碳和白桦、山杨矮曲林土壤微生物量氮呈现显著相关性, 与香杨矮曲林土壤微生物量氮相关性低; 山杨矮曲林土壤微生物量碳与香杨矮曲林土壤微生物量碳高度相关, 与其他因子相关性低; 白桦矮曲林土壤微生物量氮与山杨、香杨矮曲林土壤微生物量氮呈现高度相关性; 山杨、香杨矮曲林土壤微生物量氮呈现高度相关性。
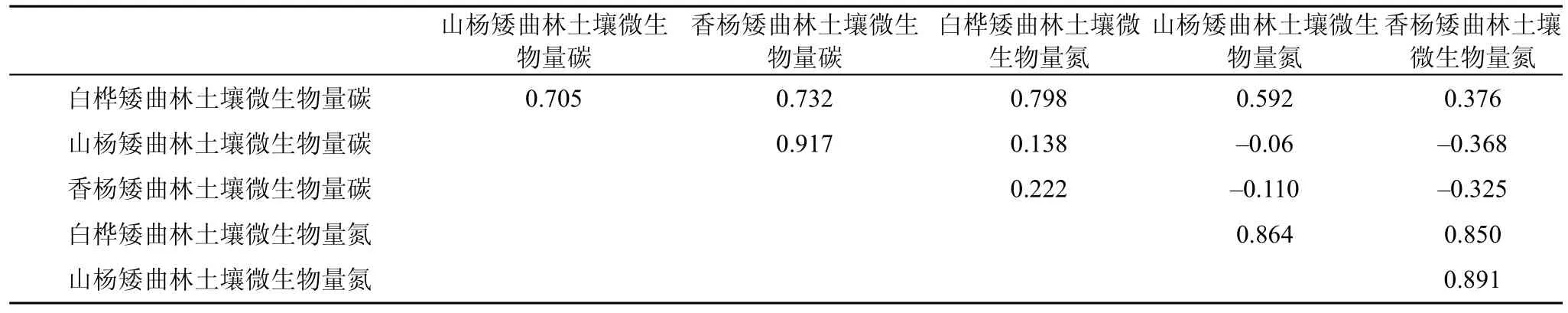
表2 三种矮曲林微生物生物量之间Pearson相关系数Tab.2 The Pearson correlation coefficient among three kinds of microbial biomass from elfin forest
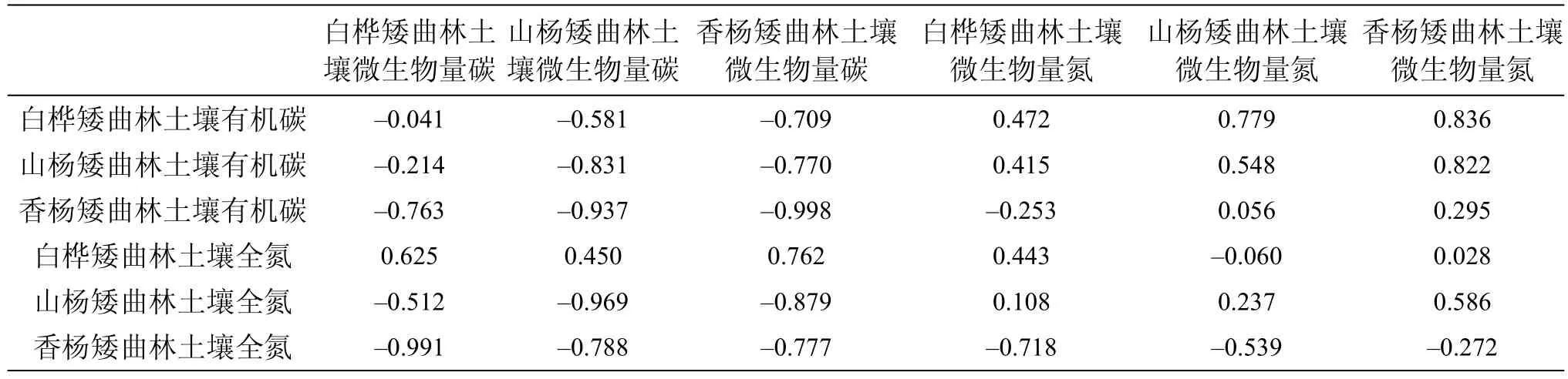
表3 三种矮曲林微生物生物量与土壤有机碳、全氮之间Pearson相关系数Tab.3 The Pearson correlation coefficient of microbial biomass to soil organic carbon and total nitrogen among three kinds of elfin forest
表3显示(P<0.05), 白桦矮曲林土壤微生物量碳与香杨矮曲林土壤全氮高度相关; 山杨矮曲林土壤白桦矮曲林土壤微生物量碳与土壤微生物量氮呈现微生物量碳与白桦矮曲林土壤有机碳、香杨矮曲林土壤全氮呈显著相关, 与香杨矮曲林土壤有机碳和山杨矮曲林土壤全氮呈负的高度相关; 香杨矮曲林土壤微生物量碳除了与香杨矮曲林土壤有机碳和山杨矮曲林土壤全氮呈现高度相关性外, 与其他因素均呈现显著相关性; 白桦矮曲林土壤微生物量氮与香杨矮曲林土壤全氮呈负的显著相关; 山杨矮曲林土壤微生物量氮与白桦、山杨矮曲林土壤有机碳和香杨矮曲林土壤全氮展现显著相关性; 香杨矮曲林土壤微生物量氮与白桦、山杨矮曲林土壤有机碳和高度相关, 与山杨矮曲林土壤全氮显著相关。
4 讨论
有研究表明, 不同生态系统土壤微生物生物量的季节变化不同[19-20]而在同一生态系统, 即使气候条件相同、不同植被下土壤微生物生物量的季节变化也不同[21], 本研究的结果显示三种矮曲林的微生物量变化趋势虽然一致, 数量的差异确很大。第一季度时, 土壤微生物量碳氮含量较高, 这是因为本地区此时处于冬春交替时期, 土壤温度开始回升,土壤水分和养分开始供应, 土壤微生物开始活动,因而微生物生物量碳氮开始增加; 进入第二季度,植物迅速萌发进入生长期, 需要从土壤中摄取大量营养物质, 在一定程度上形成了与微生物间的营养需求竞争, 致使微生物的营养可获得量降低, 限制了微生物的生长和繁殖[22], 因而此时土壤微生物量碳氮含量均急剧降低; 第三季度, 由于地表植物相继进入衰亡期, 大量凋落物的输入, 根系衰老以及碳水化合物由地上向地下的不断转移, 致使土壤中生物可利用的营养增多, 土壤微生物量碳氮含量呈现增加趋势; 最后, 在第四季度时, 由于土壤温度迅速下降, 凋落物分解速率下降, 能源供应不足,加之微生物活性迅速减弱, 微生物生物量碳呈下降状态, 这与尉海东等人[23]对鲁东南杨树人工林土壤微生物量碳的时空动态的研究、邸雪颖等[24]对兴安落叶松林土壤微生物量碳的研究、刘爽[25]对黑龙江五种温带森林土壤微生物生物量碳和王国兵[26]对北亚热带次生栎林和火炬松人工林土壤微生物生物量碳的研究结果相似。
土壤微生物生物量碳、氮在土壤中的含量较大程度上代表着土壤活性有机碳和氮的储量, 而微生物生物量商(qMB)是微生物生物量碳与土壤有机碳之比, 在不同土壤进行比较时, qMB参数可以避免由于土壤有机质含量的差异而难以说明的缺点[27]。微生物商的变化可反映土壤中有机质的输入量、微生物碳的转化速率、土壤碳的流失以及土壤矿化部分的有机碳的稳定性, 它以一种稳定的形式变化,比微生物碳或有机碳单独应用有效, 可作为土壤质量变化的一个有用指标[28]。微生物生物量和商值可以反映土壤肥力状况。原因是微物生物量碳和氮是土壤碳库、氮库中活性最高的有机成分, 易矿化, 周转快[29]。矮曲林微生物量碳与商值、土壤有机碳也大致相似(白桦林除外), 这可能是由于矮曲林土壤含水量低, 土壤中的化学、生物过程不易完成, 导致土壤养分积累量少。
相关研究表明, 降水量直接影响土壤水分含量,土壤水分又直接影响土壤气体交换、微生物养分供应和温度[30], 而本研究发现, 矮曲林土壤含水率四季变化不大, 涵养水源能力差, 微生物量呈现季节性动态变化, 由此可见, 降水量变化对土壤有机碳和全氮含量影响不大, 温度变化对其有一定影响,这与衡涛[31]对高寒草甸土壤的研究结果一致, 与其他一些结果不符, 这可能是因为影响土壤微生物的是由多个环境因子综合作用产生的, 而且本研究仅一年按季度四次采样, 周期较短、跨度较大、测定的环境因素有限, 不能完全反映出期间的关系, 这需要进一步的研究。
5 结论
(1)老黑山矮曲林的林下土壤有机碳和全氮有一定的差异性, 整体含量为白桦<山杨<香杨的趋势。由于生态恢复是一个漫长的过程, 老黑山成土量较低, 土壤养分低、涵养水源能力差, 导致土壤微生物生物量也较其他地区的含量低。
(2)老黑山矮曲林土壤微生物生物量碳氮季节变化规律相似, 微生物量碳全年整体含量为香杨<白桦<山杨, 均在第二季度含量呈现波谷值, 第三季度呈现峰值。第一、三季度生物量碳含量高于第二、四季度, 除第二季度外, 全年水平看, 山杨矮曲林土壤微生物生物量碳浮动比其他两种林型剧烈, 这与林地所处坡向、不同林型提供的底物、土壤理化性质和植被生长状况均有关系。
微生物量氮则是山杨<白桦<香杨, 季节变化整体呈“对号”趋势。均在第二季度出现波谷值, 然后随时间的持续而缓慢递增, 第四季度达到含量最高值。香杨矮曲林土壤微生物生物量氮全年波动最为明显、剧烈, 均是三种矮曲林微生物量氮的最值。
微生物量商与微生物量碳的变化规律一致, 香杨<山杨<白桦。白桦矮曲林的波动较为剧烈, 这与植物营养型、土壤养分构成等环境因素有关。
(3)白桦矮曲林土壤微生物生物量碳与其对应的生物量氮呈显著相关性(p<0.05), 与其所在的土样有机碳基本无相关性, 与土壤全氮呈现显著相关性(p<0.05)。山杨矮曲林土壤微生物生物量碳与其对应的生物量氮几乎无相关性, 与其所在的土样有机碳和全氮呈高度负相关性(p<0.05)。香杨矮曲林土壤微生物生物量碳与其对应的生物量氮几乎无相关性,与其所在的土样有机碳呈高度负相关性(p<0.05),与土壤全氮呈显著负相关性(p<0.05)。矮曲林土壤含水率四季变化不大, 涵养水源能力差, 微生物量呈现季节性动态变化, 由此可见, 降水量变化对土壤有机碳和全氮含量影响不大, 温度变化对其有一定影响。
[1]CHAPIN F S, MATSON P A, MOONEY H A.Principles of Terrestrial Ecosystem Ecology[M].New York: Springerverlag Berlin Heidelberg, 2002: 202–210.
[2]VAN B A H C, SEMENOV.A M.In search of biological indicators for soil health and disease suppression[J].Applied Soil Ecology, 2000, 15: 13–24.
[3]李延茂, 胡江春, 汪思龙, 等.森林生态系统中土壤微生物的作用与应用[J].应用生态学报, 2004, 15(10): 1943–1946.
[4]胡亚林, 汪思龙, 颜绍馗.影响土壤微生物活性与群落结构因素研究进展[J].土壤通报, 2006, 37(1): 170–176.
[5]LAURIE A T, MARY A A, RUTH D Y.Forest floor microbial biomass across a northern hardwood successional sequence[J].Soil Biology and Biochemistry, 1999, 31(3):431–439.
[6]SCHIMEL D, MELILLO J, Tian H Q, et al.Contribution of increasing CO2and climate to carbon storage by ecosystems in the United States[J].Science, 2000,287(5460): 2004–2006.
[7]WARDLE D A.A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil[J].Biological Reviews, 1992, 67(3): 321–358.
[8]MYERS R T, ZAK D R, WHITE D C, et al.Landscape-level patterns of microbial community composition and substrate use in upland forest ecosystems[J].Soil Science Society of America Journal, 2001, 65(2):359–367.
[9]SINGH J A, RAGHUBANSHI A S, SIGH S R, et al.Microbial biomass acts as a source of plant nutrients in dry tropical forest and savanna[J].Nature, 1989, 338(6215):499–500.
[10]刘恩科, 赵秉强, 李秀英, 等.不同施肥制度土壤微生物碳氮变化及细菌群落 16SrDNA V3 片段 PCR产物DGGE分析[J].生态学报, 2007, 27(3): 1082–1085.
[11]毕明丽, 宇万太, 姜子绍, 等.施肥和土壤管理对土壤微生物生物量碳、氮和群落结构的影响[J].生态学报, 2010,30(1): 0032–0042.
[12]吴建国, 艾丽.祁连山 3种典型生态系统土壤微生物活性和微生物量碳氮含量[J].植物生态学报, 2008, 32(2):465–476.
[13]何容, 王国兵, 汪家社, 等.武夷山不同海拔植被土壤微生物生物量的季节动态及其主要影响因子[J].生态学杂志, 2009, 28(3): 394–399.
[14]杨凯, 朱教君, 张金鑫, 等.不同林龄落叶松人工林土壤微生物生物量碳氮的季节变化[J].生态学报, 2009,29(10): 5500–5507..
[15]周志强, 徐丽娇, 张玉红, 等.黑龙江五大连池的生态价值分析[J].生物多样性, 2011, 19(1): 63-70.
[16]鲍士旦.土壤农化分析[M].北京, 中国农业出版社,2000: 14–61.
[17]VANCE E D, BROOKES P C, JENKINSON D S.An extraction method for measuring soil microbial biomass[J].Soil Biology and Biochemistry, 1987, 19:703–707.
[18]黄琳琪.林土壤有机碳氮分布及矿化特征[D].咸阳: 西北农林科技大学, 2015.
[19]EDWARDS K A, MCCULLOCH J, KERSHAW G, et al.Soil microbial and nutrient dynamics in a wet Arctic sedge meadow in late winter and early spring[J].Soil Biology and Biochemistry, 2006, 38(9): 2843–2851.
[20]DEVI N B, YADAVA P S.Seasonal dynamics in soil microbial biomass C, N and P in a mixed oak forest ecosystem of Manipur, North-east India[J].Applied Soil Ecology, 2006, 31(3): 220–227.
[21]刘满强, 胡锋, 何园球, 等.退化红壤不同植被恢复下土壤微生物量季节动态及其指示意义[J].土壤学报, 2003,40(6): 937–944.
[22]LI G H, CHEN Q F, HUANG Y M, et al..Soil microbial biomass C, N, P and basal respiration in rhizosphere soil of typical plants on the Loess Plateau[J].Acta Ecologica Sinica, 2010, 30(4): 976–983.
[23]尉海东, 董彬.鲁东南杨树人工林土壤微生物生物量碳的时空动态[J].生态环境学报, 2013, 22(2): 233–238.
[24]邸雪颖, 耿莹莹, 孙龙, 等.兴安落叶松林土壤微生物生物量季节动态及影响因素[J].东北林业大学学报, 2012,40(5): 1–4.
[25]刘爽, 王传宽.五种温带森林土壤微生物量碳氮的时空格局[J].生态学报, 2010, 30(12): 3135–3143.
[26]王国兵, 阮宏华, 唐燕飞, 等.北亚热带次生栎林与火炬松人工林土壤微生物生物量碳的季节动态[J].应用生态学报, 2008, 19(1): 37–42.
[27]任天志, STEFABO G.持续农业中的土壤生物指标[J].中国农业科学, 2000, 33(1): 68–75.
[28]ANDERSON T H, DOMSEH K H.Ratio of micorbial biomass carbon to total organic carbon in arable soil[J].Soi1 Biology and Biochemisty, 1989, 21: 471–479.
[29]ALLISONl MF, KILLHAM K.Response of soil microbial biomass to straw incorporation[J].Journal of Soil Science,1988, 39: 237–242
[30]TATE R L.Soil Microbiology [M].New York: Jone Wiley,2000.
[31]衡涛, 吴建国, 谢世友, 等.高寒草甸土壤碳和氮及微生物生物量碳和氮对温度与降水量变化的响应[J].中国农学通报, 2011, 27(3): 425–430.
The dynamic response characteristics of microbial biomass carbon and nitrogen in elfin forest soil to seasonal variation in Wudalianchi
JIANG Mingyue1,2,*, ZHANG Wentian1,2, WEI Xiaoxue1,2
1.Volcanoes and Mineral Springs Institute of Heilongjiang Academy of Sciences,Haerbin150080,China2.High-tech Institute of Heilongjiang Academy of Sciences,Haerbin150001,China
In this paper, 15 sampling points on the lava platform which at the foot of the Laoheishan Volcano were selected to investigate the affect of main elfin forest in Laoheishan, includingBetula patyphylla,Populus davidianaandPopulus koreana, on the microbial biomass carbon and nitrogen (MBC and MBN), as well as to compare the difference of MBC and MBN among different seasons.The results suggest that the MBC and MBN of the three main forest types from elfin forest exhibit a similar dynamic response to season variation.In addition, the values of MBC and MBN in the three main forest soil were different from each other.For MBC, it was in the order ofPopulus koreanaelfin forests <Betula platyphyllaelfin forests <Populus davidianaelfin forests, and for MBN,Populus davidiana<Betula patyphylla<Populus koreana.The values of MBC inBetula patyphyllaelfin forests are positively correlated with those of MBC and soil total nitrogen (TN) respectively (p<0.05).The value of MBC inPopulus koreanaelfin forests was negatively correlated with that of soil organic carbon (SOC) and TN respectively(p<0.05).The value of MBC inPopulus koreanaelfin forests was negatively correlated with that of SOC and TN respectively (p<0.05).The soil moisture of elfin forest changed slightly during seasons.Meanwhile, the water holding capacity of the soils was weak and the microbial biomass varied obviously in different seasons.In short, the different vegetation types and temperature variation could affect MBC and MBN.In addition, the moisture content could affect MBC and MBN slightly.
Wudalianchi; volcano; elfin forest; soil microbial biomass
10.14108/j.cnki.1008-8873.2017.05.013
S154
A
1008-8873(2017)05-096-08
姜明月, 张文天, 魏晓雪.五大连池矮曲林土壤微生物量碳、氮对季节变化的动态响应特征[J].生态科学, 2017, 36(5): 96-103.
JIANG Mingyue, ZHANG Wentian, WEI Xiaoxue.The dynamic response characteristics of microbial biomass carbon and nitrogen in elfin forest soil to seasonal variation in Wudalianchi[J].Ecological Science, 2017, 36(5): 96-103.
2016-08-08;
2016-09-10
姜明月(1987-6-11), 女, 黑龙江哈尔滨人, 硕士, 助理研究员, 主要从事环境生态学研究, E-mail: 736035905@qq.com
*通信作者:姜明月, 女, 硕士, 助理研究员 主要从事环境学研究, E-mail: 736035905@qq.com
