基于rbcL序列试论鸢尾属部分物种间的系统发育关系
2017-11-10蒋喻林刘宇婧冯艺玫蒋天仪余小芳
蒋喻林 刘宇婧 冯艺玫 谭 淼 蒋天仪 余小芳*
(1.四川农业大学风景园林学院,成都 611130; 2.成都农业科学技术学院,成都 611130)
基于rbcL序列试论鸢尾属部分物种间的系统发育关系
蒋喻林1刘宇婧1冯艺玫1谭 淼1蒋天仪2余小芳1*
(1.四川农业大学风景园林学院,成都 611130;2.成都农业科学技术学院,成都 611130)
选用叶绿体rbcL基因对鸢尾属植物的分类和系统发育进行了分析。以雄黄兰为外类群,对21份鸢尾属材料和GenBank下载的共49条序列采用最大简约法构建系统进化树,探讨了鸢尾属的分类系统。针对有争议的物种,聚类结果表明:马蔺与白花马蔺关系较远;扁竹兰和蝴蝶花亲缘关系较扇形鸢尾近;细锐果鸢尾和锐果鸢尾亲缘关系近,大锐果鸢尾较之远;四川鸢尾和薄叶鸢尾聚类近;西伯利亚鸢尾系内物种间聚类复杂,亲缘关系不清楚。
鸢尾属;rbcL序列;系统发育
鸢尾属(IrisL.)是鸢尾科(Iridaceae)中最大的一个属,属内全部物种均为多年生草本植物,具有较高的利用价值[1~2]。据不完全统计,全世界鸢尾属植物有300余种,中国约有60种13变种4变型,分布于全国各省区,尤以西南和西北最多[3]。鸢尾属由赵毓棠在1985年提出并进行分类修订[3~4],嗣后诸多学者不断从形态学、解剖学、细胞学、孢粉学和分子生物学等学科对该属做了系统分类的研究[5~16]。然而,由于各学者依据不同的形态特征和不同学科的分类证据,从而导致属下许多物种的具体分类位置仍存在分歧与争议[4,17]。
白花马蔺(IrislacteaPall.)和马蔺(Irislacteavar.chinensisFisch)因花色不同定为种和变种的关系[3],但高宝莼和赵毓棠认为两者属于一个物种[4,18]。锐果鸢尾(IrisgoniocarpaBaker)、大锐果鸢尾(IrisgoniocarpaBaker var.grossaY.T.Zhao)和细锐果鸢尾(IrisgoniocarpaBaker var.tenellaY.T.Zhao)三者形态相似,定为一个种和两个变种的关系[3],后来赵毓棠根据形态学的研究取消了细锐果鸢尾的分类地位,把它作为锐果鸢尾的异名,并将大锐果鸢尾提升为独立种[4]。高宝莼认为四川鸢尾(IrissichuanensisY.T.Zhao)和薄叶鸢尾(IrisleptophyllaLingelsh.)是同一个物种[19],得到了赵毓棠的支持[4],余小芳根据形态学、孢粉学和ITS测序的研究也发现两者亲缘关系很近[7,20]。扁竹兰(IrisconfusaSealy)、蝴蝶花(IrisjaponicaThunb.)和扇形鸢尾(IriswattiiBaker)三者亲缘关系很近[14,21],诸多学者通过形态学[22]、孢粉学[7]、细胞学[23~24]、分子生物学[25]等研究仍不能准确地对三者进行定义和划分。西伯利亚鸢尾系(Seriessiberica(Diels) Lawrence)包括云南鸢尾(IrisforrestiiDykes)、黄花鸢尾(IriswilsoniiC.H.Wright)、金脉鸢尾(IrischrysographesDykes)、西南鸢尾(IrisbulleyanaDykes)和长葶鸢尾(IrisdelavayiMicheli)[26]5个种,赵毓棠发现根据植物志中的描述很难区分这5个种,这些描述主要来自栽培种或者腊叶标本,缺乏野生种的形态考证[4],后来仲轶通过多基因组研究和杂交实验也发现这些物种的分类地位比较混乱[21]。
rbcL基因编码1,5-二磷酸核酮糖羧化酶/氧化酶(Rubisco,光合作用酶)的大亚基,现今已被广泛用于被子植物科、亚纲乃至整个种子植物主要类群间系统进化关系的研究[27~30]。Reeves等[31]通过多基因组分析了鸢尾科不同属间的关系,发现rbcL拓扑结构与基因组综合拓朴树差异较大,尤其针对某些属如Watsonia和Lapeirousia,推测rbcL序列在鸢尾科的系统分析较其它基因(trnL-F,rps4)具有更精确的信息位点。仲轶[21]的研究发现鸢尾属植物的rbcL信息位点差异在组及亚属间很少,主要存在于个体间。据此,本研究对鸢尾属14种3变种21份材料进行rbcL测序,并结合前人发表的共49条rbcL序列来探讨该属部分争议物种的系统分类位置,旨在为整个鸢尾属亲缘关系及系统分类研究提供理论依据。
1 材料与方法
本试验所用材料来自野外活体植株或四川农业大学种质资源圃(表1)。经四川农业大学周永红教授和余小芳副教授鉴定,凭证标本保存于四川农业大学小麦研究所植物标本室(SAUTI)。每个居群,随机选取5个单株,每单株取等量幼叶作为一混合样品,放入-80℃冰柜存用。
本研究DNA提取采用DN32050-植物基因组DNA提取试剂盒(成都百菲特科技有限公司)。序列选自rbcL基因的一段,长度在700 bp左右[21],引物(Forward:P1 5′-ATGTCACCACAAACAGAGACTAAAGC-3′;Reverse:P2 5′-CTTCTGCTACAAATAAGAATCGATCTC-3′)由北京擎科公司成都分部合成。
PCR扩增体系为50 μL体系:Buffer 5 μL;Mg2+4 μL;dNTP底物4 μL;模板DNA 4 μL;ex-Taq酶0.5 μL;引物1,2(10 μmol·L-1)1 μL;双蒸水(ddH2O)30.5 μL。扩增反应在Mastercycle(ProS,Eppendorf)扩增仪上进行,程序为:94℃预变性4 min,94℃变性30 sec,52℃退火30 sec,72℃延伸2 min,循环30次,72℃保温10 min,4℃保存1 h。1.5%琼脂糖凝胶电泳检测扩增产物。
DNA凝胶回收试剂盒(BioTeke,成都精博生物技术有限公司)回收,将回收产物连接到pMD19-T载体(Takara)上,16℃水浴过夜。连接产物转入大肠杆菌感受态细胞(DH5a,成都百思博生物技术有限公司)热击进行转化。菌落PCR方法筛选阳性克隆,利用2×Taq Master Mix(Dye plus,奥克生物有限公司)进行PCR,阳筛程序沿用扩增程序,用1%琼脂糖凝胶进行电泳检测。选取8个阳性克隆到生工生物工程(上海)股份有限公司测序。
对测序得到的结果用DNAman软件在NCBI网址上进行BLAST(http://blast.ncbi.nlm.nih.gov/Blast.cgi),检测以便排除微生物或其它污染物的目的片段干扰,选取测序正确的基因片段,并上传序列至数据库。用Clustal W进行序列比对[32],然后用DAMBE进行手工校正和转换格式[33],最后用MEGA 6.0和PAUP* Version 4.0进行系统发育分析[34]。采用最大简约法(Maximum Parsimony),对排好的序列中所有的碱基同等加权(equal weighted)并作为无序(unordered)性状,空位(gap)作为缺失(missing)处理[57],Bootstrap Method添加方式循环1 000次,TBR(tree bisection-reconnection)枝长交换,进行启发式搜索(Heuristic Search),拓扑结构的可靠性用1 000次重复的自展检验来完成[35]。
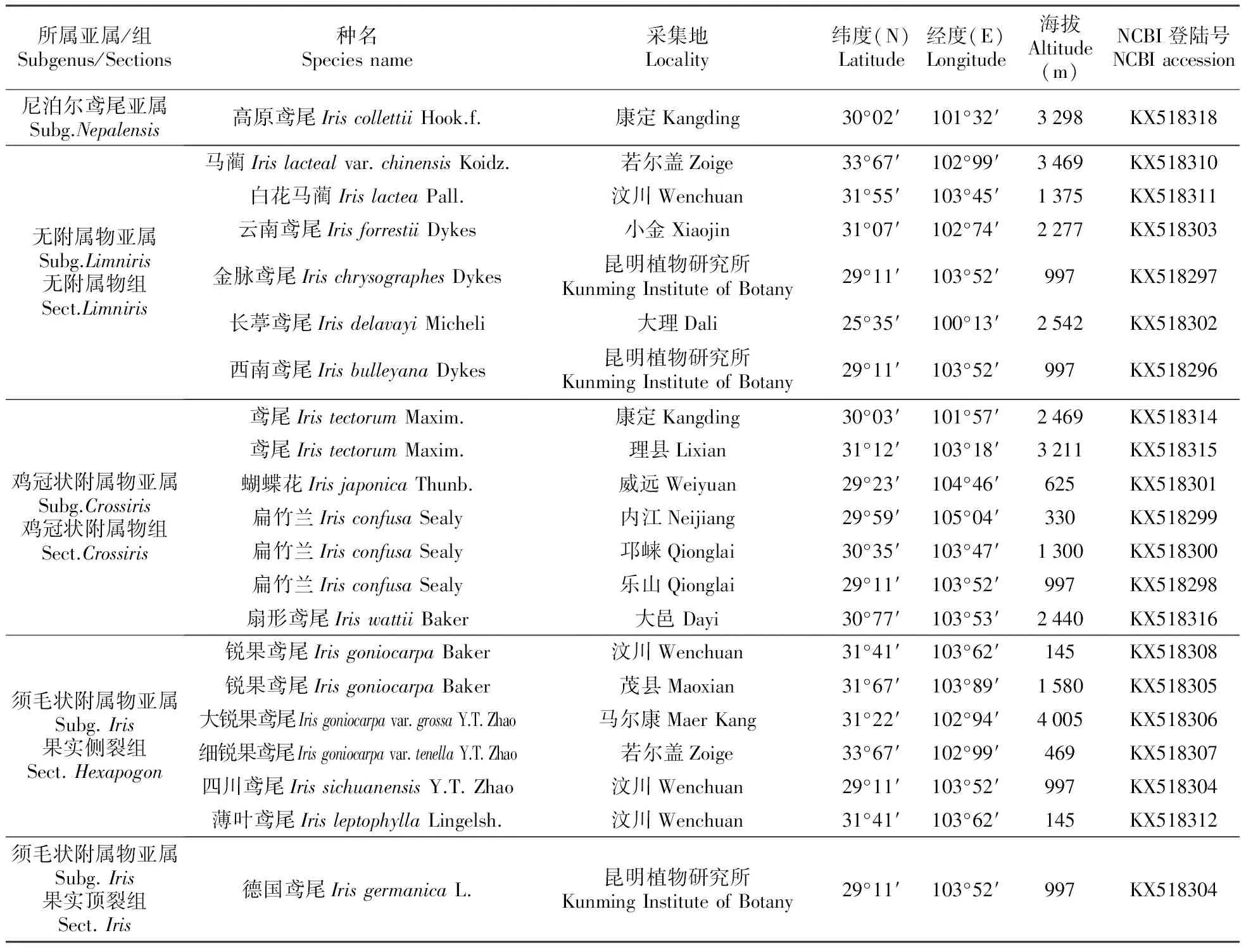
表1 鸢尾属植物材料来源及序列编号
2 实验结果
经PCR扩增、克隆测序及序列检测选择后,共获得鸢尾属21份材料的rbcL序列。选择雄黄兰(Crocosmiacrocosmiflora(Nichols.) N.E.Br.)作为外类群,结合前人发表的27条rbcL序列(表2)共同进行聚类分析。
2.1 序列信息
基因序列的碱基频率检测显示T占28.7%,C占20.2%,A占28.0%,G占23.1%,两个基因碱基A+T平均含量56.6%高于G+C的平均含量43.4%。经比对排序,序列总共647个排列位点,长度变异在617~647 bp。利用MEGA 6.0进行了排列矩阵数据分析,其中rbcL保守位点(Conserved Sites)575个、简约信息位点(Parsimony-informative Sites,PI)36个、变异位点(Variable Sites)63个、自裔位点(Singleton Sites)27个,变异位点占总序列长度的10%。
rbcL基因的两两序列间碱基替换统计显示:碱基转换对(Transitional Pairs,si)有20个;碱基颠换对(Transversional Pairs,sv)有10个;转换对与颠换对的比值(R=si/sv)为2。该统计表明转换频率显著高于颠换频率,序列可用于系统发育分析。用MEGA 6.0软件进行序列间碱基组成偏倚差异估算,结果显示全数据组的平均值P=0.011 4,结果显著(P<0.05),所以拒绝零假设(null hypothesis,即序列以相同的替换模式进化),因此各分类单元间不存在碱基组成偏向性。
2.2 中性检测
为鉴定DNA序列在长期进化过程中是否遵循中性进化模型,利用MEGA 6.0软件进行Tajima Test(Tajima’s D)中性统计检验[36]。序列变化总位点N(total number of sites)有609个,多态性位点数目S(Number of segregating sites)有62个,多态性比率Ps(ps=S/N)为0.101 806和核苷酸变异π(nucleotide diversity)值为0.013 272。rbcL基因序列Tajima’s D值为-1.481 759(P<0.05),明显不遵循中性进化模型。

表2 NCBI下载的rbcL序列
2.3 聚类分析结果
总共49条序列构建的系统进化树见图1,序列代表了中国鸢尾属的5个不同亚属(不包含琴瓣鸢尾亚属SubgenusXyridionSpach)。外类群雄黄兰以100%的支持率聚到鸢尾属外面,鸢尾属形成两个大支Clade 1(支持率Popularity rating,PP=100%)和Clade 2(PP=100%)。Clade 1代表种类最多的无附属物亚属,该亚属又形成5个小的聚类支(图1,黑色箭头)。Clade 2代表有附属物的其他3个亚属(尼泊尔鸢尾亚属、须毛状附属物亚属、鸡冠状附属物亚属)和野鸢尾亚属(PP=100%)。鸡冠状附属物亚属形成两个聚类支(图1,黑色箭头),其中一支(PP=92%)与尼泊尔鸢尾亚属聚到一起,但该亚属的小花鸢尾(Irisspeculatrix)作为独立支(PP=100%)和无附属物亚属聚到一起。
对于有争议物种的聚类情况为:白花马蔺(I.lactea)的3个类群聚成一小支(PP=100%),其变种马蔺(I.lacteavar.chinensis)却镶嵌到了无附属物亚属的其他聚类支(PP=100%);蝴蝶花(I.japonica)和扁竹兰(I.confusa)居群聚到一起(PP=100%),扇形鸢尾(I.wattii)和鸢尾(I.tectorum)聚到一起(PP=100%);锐果鸢尾(I.goniocarpa)和细锐果鸢尾(I.goniocarpavar.tenella)聚到一起(PP=100%),大锐果鸢尾(I.goniocarpavar.grossa)聚到另外一小支(PP=96%);四川鸢尾(I.sichuanensis)和薄叶鸢尾(I.leptophylla)聚到一起;西伯利亚鸢尾系形成两个小支,支持率分别为100%和58%(图1,竖线)。

图1 基于rbcL基因系列构建的MP树 黑色粗体字母代表本研究所涉及的类群产地;粗体数字代表支持率,图中已经舍去低于50%的支持率。Fig.3 MP tree based on the rbcL gene Species Locality of this study was indicated by Bold Font;Bold type Numbers at nodes indicate bootstrap values 50%.
3 讨论
3.1 序列分析
rbcL基因相对叶绿体基因间隔区序列较保守[29],变异位点占总序列长度的10%。由于rbcL基因是在光合作用及光呼吸中起关键作用的编码基因,进化过程中受到功能限制,故其序列相对保守[27]。这些变异位点较均匀地分布于rbcL基因所选区段上,这和前人研究结果一致[30]。本研究中该序列信息分析显示碱基转换频率高于碱基颠换频率,这符合进化过程中近缘物种之间或种内其编码区DNA序列容易发生同义突变的规律[35]。中性检测发现rbcL的D值为负,推测鸢尾属在演化过程中经历过种群扩张或瓶颈事件,但平行演化事件少,说明该系统树较为真实[36]。整个聚类树中,鸢尾属不同亚属及组的聚类存在相互渗透的现象,组及亚属之间的划分不清晰,rbcL基因还表现出个体间差异较明显的特点,这和前人的研究结果相似[21,31]。
3.2 鸢尾属的分类系统
笔者采用赵毓棠的分类系统(表3),本研究聚类结果大体支持该分类系统。该系统属下分为6个亚属,其中无附属物亚属(Subg.Limniris(Tausch) Spach)的物种数量最多,分布范围最广,亚属下再分为3个组:紫苞鸢尾组(Section.Ioniris)、无附属物组(Sect.Limniris)和单苞鸢尾组(Sect.Ophioris)。本研究中紫苞鸢尾组和单苞鸢尾组都镶嵌到无附属物组里,推测这两个组起源于无附属物组,而后独立分化出去。但紫苞鸢尾组中的变种窄叶单花鸢尾(I.unifloravar.caricina)聚到长尾鸢尾和玉蝉花的小支里(PP=80%),推测该组内物种在进化过程中存在分化现象。谢航[37]曾认为紫苞鸢尾组应归为无附属物组中紫苞鸢尾系(SeriesRuthenicae),但王玲[38]根据种子表面微形态和种苗形态结果认为该组应独立存在,本研究支持后者。无附属物亚属分类群是一个极不自然的类群[14],相较于其他亚属起源较早[25],本研究中该属属下各物种聚类较为分散,推测无附属物亚属可能是多起源的,且各物种的起源和亲缘关系比较复杂。

表3 赵毓棠的分类系统
须毛状附属物亚属(Subg.Iris)聚成3个小支,包括长白鸢尾和粗根鸢尾聚成的一小支(PP=74%),膜苞鸢尾和德国鸢尾聚成的一小支(PP=100%),及包含四川鸢尾和薄叶鸢尾的锐果鸢尾类群支(PP=96%),这3个小支分布较为分散,表现出较远的亲缘关系,说明该亚属的遗传多样性较高。长白鸢尾(I.mandshurica)一直是有争议的物种,王玲认为其种苗形态和花粉形态都与无附属物亚属物种相似,并推测其为无附属物亚属与须毛状附属物亚属之间的过渡类型,在本研究中,长白鸢尾和须毛状附属物亚属中的物种聚到一起,这和王玲的研究结果不同[39]。果实顶裂组(Sect.Iris)的德国鸢尾(I.germanica)和果实侧裂组(Sect.Hexapogon)的膜苞鸢尾(I.scariosa)聚在一小支,表现出该亚属组间的划分不清晰,这与Wilson的研究结果一致[40],推测须毛状附属物亚属内各物种之间的分化程度与时间可能存在较大差异,建议对该亚属的组下级别做进一步研究。
鸡冠状附属物亚属(Subg.Crossiris)包含鸡冠状附属物组(Sect.Crossiris)和小鸢尾组(Sect.Lophiris)。本研究中该亚属的鸡冠状附属物组分裂成为关系较远的两支,且该组中的小花鸢尾单独和无附属物亚属聚到一起,这与Guo和Wilson的结果一致[1]。早前Rodionenko认为小花鸢尾拥有外源血缘,暂时把它归类到该亚属[41],目前还鲜有该种的文献报道,本文建议针对该种做进一步研究。Mathew曾将鸡冠状附属物亚属归类到无附属物亚属[42],Rodionenko和Wilson将其提升为亚属[25,41],仲轶[21]和Wilson[40]通过花粉形态、同工酶、数量性状等研究发现该亚属的位置存在较大争议,本研究结果支持该观点,建议针对该亚属做深入研究。
尼泊尔鸢尾亚属(Subg.Nepalensis)包含尼泊尔鸢尾和高原鸢尾[3],该亚属区别于其他亚属的主要特点为:拥有纺锤状的肉质根。本研究中,两个种聚到一起,赵毓棠认为尼泊尔鸢尾亚属是一个比较自然的分类群[4],本研究同意其观点。野鸢尾亚属(Subg.Pardanthopsis)的野鸢尾和射干属(GenusBelamcanda)的射干聚到一起(PP=100%)。早前Rodionenko把野鸢尾放到亚属的位置[41],随后Mathew把它提升为独立的属[42]。很多学者系统发育分析发现野鸢尾都聚类到鸢尾属中[3,43~44],故本研究也支持将其作为亚属处理。射干曾被发现聚到鸢尾属中[17,40],Goldblatt已经把射干属(Belamcanda)归类到了鸢尾属里[45],本研究支持这一处理。
3.3 有争议物种的分类地位
3.3.1 马蔺和白花马蔺的系统发育关系
马蔺与白花马蔺的植株形态极其相似,主要分化特点为花色变异,且同一居群里会出现不同花色的植株,高宝莼和赵毓棠认为两者为同一物种[4,18]。余小芳通过孢粉学研究发现白花马蔺和马蔺花粉大小相似,形状、萌发孔、外壁纹饰和网脊形态相同,但网脊宽度不同,通过细胞学研究发现两者的核型不同,认为马蔺作为白花马蔺的变种是可行的[20]。
本研究中,3个不同来源的白花马蔺聚到一起,其变种马蔺聚到另外一支,两者表现出较远的亲缘关系,支持将马蔺作为白花马蔺的变种处理。马蔺类群分布范围很广,牟少华[46]对不同种源的马蔺进行了AFLP研究,认为马蔺非单起源,且种内遗传多样性高,其分化具有地理变异性,其中海拔和纬度是重要的影响因子。本研究中白花马蔺来自四川汶川(海拔1 375 m),马蔺来自四川若尔盖(海拔3 469 m),两居群间生境存在明显差异,推测这是导致两者聚类较远的原因。
3.3.2 扁竹兰、蝴蝶花和扇形鸢尾的系统发育关系
扁竹兰、蝴蝶花和扇形鸢尾都有相似的形态特征,难以区分[1],三者之间的亲缘关系一直存在争议[7,22~25]。本研究中蝴蝶花和扁竹兰聚到一起(PP=100%),扇形鸢尾和鸢尾聚到一起(PP=100%),且两分类支相距较远,支持扁竹兰与蝴蝶花的亲缘关系较扇形鸢尾更近[7]。研究还发现扁竹兰聚到蝴蝶花类群里,说明后者拥有更高的遗传多样性且更为原始,这和张敏等的研究结果相似[16]。Waddick认为蝴蝶花和扁竹兰来自同一个物种[47]。笔者在形态观察中发现扁竹兰和蝴蝶花的花茎形状不同,前者花茎呈“之”字形,后者花茎呈直立状,从花茎上可以将二者区分开来。
3.3.3锐果鸢尾、大锐果鸢尾和细锐果鸢尾的系统发育关系
锐果鸢尾的不同居群间存在显著的形态差异,种内遗传多样性较高[21]。赵毓棠基于形态将大锐果鸢尾提升为独立种,并更名为IriscuniculiformisNoltie & K.Y.Guan,同时取消了细锐果鸢尾的分类地位[4]。本研究中,锐果鸢尾的两个居群和细锐果鸢尾聚到一起,大锐果鸢尾聚到另外一小支,说明锐果鸢尾和细锐果鸢尾关系很近,大锐果鸢尾与两者关系较远。形态观察发现:大锐果鸢尾和锐果鸢尾确实存在明显的差异,如大锐果鸢尾叶片较厚且顶端圆盾,花色深紫,花被片上深紫色斑点更为明显;锐果鸢尾和细锐果鸢尾形态相似,结合聚类分析本实验结果支持赵毓棠的分类处理。
3.3.4 四川鸢尾和薄叶鸢尾的系统发育关系
本研究中四川鸢尾(I.sichuanensis)和薄叶鸢尾(I.leptophylla)聚在一起,表明两者亲缘关系很近。前人发现两者形态相似[11,38,44],赵毓棠和高宝莼认为两者是同一物种[4,19],余小芳通过核型分析建议两者应为独立种[20]。形态观测发现:薄叶鸢尾植株较小,叶片质地柔软;四川鸢尾叶片较厚略披白粉,叶脉更加明显,花色略红,两者可通过形态进行区分。如何进一步对它们分类处理,还需进行更多方面的资料综合分析后方能得出。
3.3.5 西伯利亚鸢尾系系统发育关系
本研究中,西伯利亚鸢尾系的四个物种聚到不同的小支,表现出较复杂的亲缘关系。不同来源的金脉鸢尾并未聚到一起,西南鸢尾与其变种白花西南鸢尾也未聚到一起,但金脉鸢尾、西南鸢尾和长葶鸢尾表现出较近的亲缘关系,三者与云南鸢尾关系较远。笔者发现长葶鸢尾的植株高大,不呈丛状,叶片宽厚披白粉,花茎高于植株,花大、花色深紫;云南鸢尾丛状,叶黄绿色,花黄色,这两种能明显被区分。但金脉鸢尾和西南鸢尾植株形态与花形态都很相似[4],两者在形态上极易混淆[21],故本文建议对两者进行更深入的研究。
1.Guo J Y,Wilson C A.Molecular phylogeny of crestedIrisbased on five plastid markers(Iridaceae)[J].Systematic Botany,2013,38(4):987-995.
2.Raven P H,Axelrod D I.Angiosperm biogeography and past continental movements[J].Annals of the Missouri Botanical Garden,1974,61(3):539-673.
3.赵毓棠.中国植物志[M].北京:科学出版社,1985:120-198.
Zhao Y T.Flora of China[M].Beijing:Science Press,1985:120-198.
4.Zhao Y T,Noltie H J,Mathew B.Iridaceae[M].//Wu Z Y,Raven P H.Flora of China.Beijing,China,St.Louis,Missouri,USA:Science Press,Missouri Botanical Garden Press,2000:297-313.
5.Wilson C A.A Cladistic analysis ofIrisseriesCalifornicaebased on morphological data[J].Systematic Botany,1998,23(1):73-88.
6.齐耀斌,赵毓棠.中国鸢尾属花粉形态研究[J].植物分类学报,1987,25(6):430-436.
Qi Y B,Zhao Y T.Studies on the pollen morphology of the genusIrisin China[J].Journal of Systematics and Evolution,1987,25(6):430-436.
7.余小芳,张海琴,何雪梅,等.鸢尾属12种(变种)植物花粉形态及其系统学意义[J].园艺学报,2010,37(7):1175-1182.
Yu X F,Zhang H Q,He X M,et al.Pollen morphology of 12 species ofIrisL.and it’s systematic significations[J].Acta Horticulturae Sinica,2010,37(7):1175-1182.
8.Mitió B,Halbritter H,oštaric R,et al.Pollen morphology of the genusIrisL.(Iridaceae) from Croatia and surrounding area:taxonomic and phylogenetic implications[J].Plant Systematics and Evolution,2013,299(1):271-288.
9.周永红,伍碧华,颜济,等.Irisjaponica×Irisconfusa种间杂种的细胞遗传学研究[J].云南植物研究,2003,25(4):497-502.
Zhou Y H,Wu B H,Yan J,et al.Cytogenetic study on the interspecific hybrid betweenIrisjaponicaandIrisconfusa(Irisaceae)[J].Acta Botanica Yunnanica,2003,25(4):497-502.
10.Ikinci N,Hall T,Lledó M D,et al.Molecular phylogenetics of the JunoIrises,IrissubgenusScorpiris(Iridaceae),based on six plastid markers[J].Botanical Journal of the Linnean Society,2011,167(3):281-300.
11.沈云光,王仲朗,管开云.国产13种鸢尾属植物的核型研究[J].植物分类学报,2007,45(5):601-618.
Shen Y G,Wang Z L,Guan K Y.Karyotypical studies on thirteenIrisplants from China[J].Acta Phytotaxonomica Sinica,2007,45(5):601-618.
12.Wilson C A.Phylogenetic relationships inIrisseriesCalifornicaebased on ITS sequences of nuclear ribosomal DNA[J].Systematic Botany,2003,28(1):39-46.
13.Wheeler A S,Wilson C A.Exploring phylogenetic relationships within a broadly distributed northern hemisphere group of semi-aquaticIrisspecies(Iridaceae)[J].Systematic Botany,2014,39(3):759-766.
14.牟少华,彭镇华,郄光发,等.部分鸢尾属植物的AFLP标记[J].东北林业大学学报,2011,39(1):124-126.
Mu S H,Peng Z H,Qie G F,et al.Genetic diversity ofIrisdetermined by AFLP marker[J].Journal of Northeast Forestry University,2011,39(1):124-126.
15.张敏,黄苏珍.鸢尾属部分植物种质资源的RAPD分析[J].西北植物学报,2008,28(5):933-939.
Zhang M,Huang S Z.RAPD analysis on some germplasm ofIrisL.plants[J].Acta Botanica Boreali-Occidentalia Sinica,2008,28(5):933-939.
16.张敏,黄苏珍,仇硕,等.鸢尾属植物遗传多样性的RAPD和ISSR分析[J].植物资源与环境学报,2007,16(2):6-11.
Zhang M,Huang S Z,Qiu S,et al.Analysis of RAPD and ISSR on genetic diversity ofIrisL.plants[J].Journal of Plant Resources and Environment,2007,16(2):6-11.
17.Wilson C A.Phylogeny ofIrisbased on chloroplastmatK Gene andtrnK intron sequence data[J].Molecular Phylogenetics and Evolution,2004,33(2):402-412.
18.高宝莼,谭仲明.四川植物志:第9卷[M].成都:四川民族出版社,1989:320-370.
Gao B C,Tan Z M.Flora Sichuanica:vol.9[M].Chengdu:Sichuan National Publishing House,1989:320-370.
19.高宝莼.四川鸢尾属的研究[J].西南师范学院学报,1985(3):11-15.
Gao B C.Notes on the genusIrisof Sichuan[J].Journal of the Southeast Teachers Collage,1985(3):11-15.
20.余小芳.四川鸢尾属植物的系统学及种子休眠与萌发特性研究[D].雅安:四川农业大学,2009.
Yu X F.Systematic study and seed dormancy and germination characters ofIrisspieces in Sichuan[D].Ya’an:Sichuan Agricultural University,2009.
21.仲轶.基于多基因组的鸢尾属部分种的系统分类研究[D].哈尔滨:东北林业大学,2010.
Zhong Y.The system classification research base on multi-genome of someIrisspecies[D].Harbin:Northeast Forestry University,2010.
22.Colasante M,Sauer W.The genusIrisL.in Latium(central Italy):general remarks on the morphological and karyological differentiation as well as on the ecological adaptation and flowering periods[J].Linzer Biologische Beiträge,1993,25(2):1189-1204.
23.舒焕麟,颜济,杨俊良.IrisconfusaSealy的细胞学研究[J].四川农业大学学报,1992,10(4):586-591.
Shu H L,Yan J,Yang J L.Cytological study onIrisconfusaSealy[J].Journal of Sichuan Agricultural University,1992,10(4):586-591.
24.毛节绮,薛详骥.浙江几种鸢尾科植物的染色体[J].浙江农业大学学报,1986,12(1):97-101.
Mao J Q,Xue X J.Chromosome numbers of thirteen iridaceous species from Zhejiang province[J].Journal of Zhejiang Agricultural University,1986,12(1):97-101.
25.Wilson C A.Phylogenetic relationships among the recognized series inIrissectionLimniris[J].Systematic Botany,2009,34(2):277-284.
26.Makarevitch I,Golovnina K,Scherbik S,et al.Phylogenetic relationships of the SiberianIrisspecies inferred from noncoding chloroplast DNA sequences[J].International Journal of Plant Sciences,2003,164(2):229-237.
27.Chase M W,Soltis D E,Olmstead R G,et al.Palmer,phylogenetics of seed plants:an analysis of nucleotide sequences from the plastid generbcL[J].Annals of the Missouri Botanical Garden,1993,80(3):528-580.
28.Soltis D E,Soltis P S,Chase M W,et al.Angiosperm phylogeny inferred from 18S rDNA,rbcL,andatpB sequences[J].Botanical Journal of the Linnean Society,2000,133(4):381-461.
29.Donoghue M J,Olmstead R G,Smith J F,et al.Phylogenetic relationships ofDipsacalesbased onrbcL sequences[J].Annals of the Missouri Botanical Garden,1992,79(2):333-345.
30.Chase M W,Bruijn A Y D,Cox A V,et al.Phylogenetics of asphodelaceae(asparagales):an analysis of plastidrbcL andtrnL-F DNA sequences[J].Annals of Botany,2000,86(5):935-951.
31.Reeves G,Chase M W,Goldblatt P,et al.Molecular systematics of iridaceae:evidence from four plastid DNA regions[J].American Journal of Botany,2001,88(11):2074-2087.
32.Thompson J D,Higgins D G,Gibson T J.CLUSTAL W:improving the sensitivity of progressive multiple sequence alignment through sequence weighting,position-specific gap penalties and weight matrix choice[J].Nucleic Acids Research,1994,22(22):4673-4680.
33.Xia X H.DAMBE5:a comprehensive software package for data analysis in molecular biology and evolution[J].Molecular Biology and Evolution,2013,30(7):1720-1728.
34.Tamura K,Stecher G,Peterson D,et al.MEGA6:molecular evolutionary genetics analysis version 6.0[J].Molecular Biology and Evolution,2013,30(12):2725-2729.
35.Kimura M.A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences[J].Journal of Molecular Evolution,1980,16(2):111-120.
36.Tajima F.Statistical methods to test for nucleotide mutation hypothesis by DNA polymorphism[J].Genetics,1989,123:585-595.
37.谢航.中国鸢尾属有关分类群的讨论及属下分类系统的修订[D].长春:东北师范大学,1996.
Xie H.Discussion of some taxa and revision of the classification system of ChineseIris[D].Changchun:Northeast Normal University,1996.
38.王玲,卓丽环.鸢尾属部分种种子微形态特征与系统演化关系[J].植物研究,2006,26(3):286-290.
Wang L,Zhuo L H.The relationship between seed coat micro-morphology characteristics and systemic evolution of some species ofIris[J].Bulletin of Botanical Research,2006,26(3):286-290.
39.王玲,卓丽环.基于ITS序列的鸢尾属植物部分种的系统分类[J].东北林业大学学报,2006,34(4):54-56,75.
Wang L,Zhuo L H.Systematic classification of some species ofIrisbased on ITS sequences[J].Journal of Northeast Forestry University,2006,34(4):54-56,75.
40.Wilson C A.Subgeneric classification inIrisre-examined using chloroplast sequence data[J].Taxon,2011,60(1):27-35.
41.Rodionenko G I.The genusIrisL.[M].London:The British Iris Society,1987.
42.Mathew B.TheIris[M].London:B.T.Batsford,1989.
43.Wilson C A,Calvin C L.An origin of aerial branch parasitism in the mistletoe family,Loranthaceae[J].American Journal of Botany,2006,93(5):787-796.
44.牟少华,孙振元,彭镇华.基于叶绿体DNAtrnL-F序列研究部分鸢尾属的亲缘关系[J].华北农学报,2010,25(5):112-116.
Mu S H,Sun Z Y,Peng Z H.Relationship ofIrisinferred from the chloroplast DNAtrnL-F sequence variation[J].Acta Agriculturae Boreali-Sinica,2010,25(5):112-116.
45.Goldblatt P,Manning J C.TheIrisfamily:natural history and classification[M].Portland:Timber Press,2008.
46.牟少华,彭镇华,郄光发,等.马蔺种质资源AFLP标记遗传多样性分析[J].安徽农业大学学报,2008,35(1):95-98.
Mu S H,Peng Z H,Qie G F,et al.Genetic diversity ofIrislacteadetermined by AFLP marker[J].Journal of Anhui Agricultural University,2008,35(1):95-98.
47.Waddick J W.CollectingIrisesin western China[J].Bulletin of the American Iris Society,1994(3):74-92.
The Education Department of Sichuan Province(13ZB0295)
introduction:JIANG Yu-Lin(1990—),female,Master degree candidate,Engaged in phylogenetic classification research of plants.
date:2016-11-14
PhylogeneticRelationshipofSeveralSpeciesfromIrisL.BasedonrbcLSequences
JIANG Yu-Lin1LIU Yu-Jing1FENG Yi-Mei1TAN Miao1JIANG Tian-Yi1YU Xiao-Fang1*
(1.College of Landscape Architecture,Sichuan Agricultural University,Chengdu 611130;2.Chengdu Agricultural College,Chengdu 611130)
The phylogenesis ofIrisL. was analyzed using the chloroplast genes rbcL. Classification was performed with maximum parsimony method and the gene tree were constructed with Crocosmia crocosmiflora as outgroup, 21 wild populations ofIrisL. from China and download 49 sequences from GenBank. As for disputed species, clustering trees results showedIrislacteavar.chinensis(Fisch.) Koidz. had close relationship withIrislacteaPall..IrisconfusaSealy had more close relationship withIrisjaponicaThunb. than that withIriswattiiBaker.Irisgoniocarpavar.tenellaY.T. Zhao was very relevant toIrisgoniocarpaBaker, butIrisgoniocarpavar.grossaY.T. Zhao was distant.IrissichuanensisY.T. Zhao andIrisleptophyllaLingelsh. were closely related. Clustering relationships of Series Sibirica were very complexed, and genetic relationgships were not clear.
IrisL.;rbcL sequence;systematic classification
四川省教育厅项目(13ZB0295)资助
蒋喻林(1990—),女,硕士研究生,主要从事植物系统进化分类研究。
* 通信作者:E-mail:xiaofangyu@sicau.edu.cn
2016-11-14
* Corresponding author:E-mail:xiaofangyu@sicau.edu.cn
Q949.71+8.28
A
10.7525/j.issn.1673-5102.2017.03.005
