黑翅土白蚁菌圃对采食工蚁的引诱活性
2017-11-07王亚召嵇保中刘曙雯徐立军
王亚召,嵇保中*,刘曙雯,丁 芳,徐立军
(1. 南京林业大学南方现代林业协同创新中心,南京林业大学林学院,南京 210037;2. 南京中山陵园管理局,南京 210014)
黑翅土白蚁菌圃对采食工蚁的引诱活性
王亚召1,嵇保中1*,刘曙雯2*,丁 芳1,徐立军1
(1. 南京林业大学南方现代林业协同创新中心,南京林业大学林学院,南京 210037;2. 南京中山陵园管理局,南京 210014)
黑翅土白蚁是一种重要的培菌白蚁,采食工蚁取食菌圃物质,采集树皮等植物材料作为菌圃的培养基。本文研究了菌圃、林间采集材料(杂交鹅掌楸树干外表皮)对采食工蚁的引诱活性,并与常用的白蚁饵料基质密粘褶菌松木粉培养物进行了比较。利用Y形嗅觉仪分别测定上、中、下层主巢菌圃、副巢菌圃、杂交鹅掌楸树干外表皮、密粘褶菌松木粉培养物对采食工蚁的引诱活性;比较了主巢下层菌圃与杂交鹅掌楸树干外表皮、主巢下层菌圃与密粘褶菌松木粉培养物、副巢菌圃与杂交鹅掌楸树干外表皮、副巢菌圃与密粘褶菌松木粉培养物、杂交鹅掌楸树干外表皮与密粘褶菌松木粉培养物的引诱活性。结果显示,(1)空白对照条件下,不同测定材料趋性反应率大小顺序:密粘褶菌松木粉培养物>副巢菌圃>主巢下层菌圃>主巢中层菌圃>主巢上层菌圃>杂交鹅掌楸树干外表皮。不同材料的趋性反应率显著性检验:密粘褶菌松木粉培养物与副巢菌圃、副巢菌圃与主巢下层菌圃、主巢下层菌圃与主巢中层菌圃、主巢中层菌圃与主巢上层菌圃、主巢上层菌圃与杂交鹅掌楸树干外表皮之间差异不显著,其余各组差异显著或极显著。(2)不同测定材料趋势反应率两两比较测定:密粘褶菌松木粉培养物高于杂交鹅掌楸树干外表皮(P<0.01);副巢菌圃高于杂交鹅掌楸树干外表皮(P<0.01);主巢下层菌圃高于杂交鹅掌楸树干外表皮(P<0.05);副巢菌圃与密粘褶菌松木粉培养物、主巢下层菌圃与密粘褶菌松木粉培养物差异不显著。(3)黑翅土白蚁共生菌圃对采食工蚁具有明显的引诱活性,引诱活性强度与菌圃的类型、部位有关。(4)主巢下层菌圃、副巢菌圃有较高的引诱活性,引诱活性强度与密粘褶菌松木粉培养物相近。相比之下,主巢中层菌圃、主巢上层菌圃以及杂交鹅掌楸树干外表皮引诱活性较低。菌圃物质对采食工蚁的引诱活性比林间采集材料的引诱活性更高。共生菌圃中引诱活性组分的理化性质、组成特点、来源等尚需进一步研究。
黑翅土白蚁;采食工蚁;菌圃;引诱活性
在长期演化过程中,有些白蚁与真菌形成了较为紧密的食物联系,如白蚁对一些木腐菌侵染物具有嗜食习性,两者之间形成相互促进的关系,成为森林生态系统中颇具特色的物质降解因子(Silvaetal., 2007;李姝江和朱天辉,2011)。在澳大利亚桉树林中,木腐菌通过森林火灾等形成的树干伤口侵入树体,不仅对树的木质纤维素进行分解,其菌丝还提高了腐朽物中的氮素营养,同时真菌感染降低了树木质部的树脂含量,对白蚁取食具有解毒作用,使树木降低了对白蚁的抗性。真菌腐朽和白蚁取食成为桉树林内常见的关联现象,且白蚁取食范围仅限于真菌腐朽部位(Perryetal., 1985;丁芳等,2015a)。这种现象在巴西亚马逊热带雨林也普遍存在,涉及的白蚁有乳白蚁Coptotermessp.、球白蚁Glossotermesoculatus、厚唇白蚁Labiotermesleptothrix,树种包括橡胶树Heveabrasiliensis、普通正玉蕊木Lecythisusitata、油桃木Caryocarpallidum、亚马逊安尼樟Anibaduckei等(Apolinário and Martius,2004)。在上述木腐菌与白蚁形成复合侵染取食的过程中,白蚁通过构筑泥被等行为,提高了木材腐朽部位的湿度,形成了有利于真菌发育的环境(Hennonetal., 2007;Kirkeretal., 2012)。除木腐菌外,褐腐菌以及蓝变真菌感染的木材对乳白蚁Coptotermessp.、散白蚁Reticulitermesspp.等的取食也有引诱活性(Kirkeretal., 2012;Littleetal., 2012a, 2012b, 2013)。密粘褶菌Gloeophyllumtrabeum在白蚁嗜食真菌腐朽物方面研究较为深入,该菌感染木材后引起褐色腐朽,其感染形成的松木腐朽物作为白蚁诱饵剂的基本材料,加入长效慢性杀虫剂制成商品毒饵,成为白蚁诱杀和种群监测的重要制剂,在美国、澳大利亚、加拿大等国得到广泛应用(Kirkeretal., 2012;丁芳等,2015a)。与白蚁有重要食物联系的另一类真菌为蚁巢伞属Termitomyces,这类真菌与白蚁科Termitidae大白蚁亚科Macrotemitinae的一些种类形成共生关系。这类白蚁有培植菌圃习性,称为培菌白蚁(fungus-growing termites)。采食工蚁出巢采集植物材料,巢内工蚁取食菌圃菌丝、分生孢子以及植物材料并在菌圃表面排粪,粪便中含有初步消化并清除杂菌的植物材料和菌圃真菌分生孢子,粪便中分生孢子萌发完成新一代菌圃的接种。采食工蚁则取食菌圃下层物质,上述菌圃构建过程、不同品级和发育阶段个体取食菌圃物质的部位以及摄食途径等最先在一些大白蚁如:可可大白蚁Macrotermesbellicosus;Macrotermessp.中得到阐明(Hinzeetal., 2002;Aanenetal., 2007;Nobreetal., 2011;王鹏飞等,2012)。黑翅土白蚁Odontotermesformosanus不仅是危害树木、堤坝、建筑等的重要害虫,同时也是一种重要的培菌白蚁;其菌圃培植与利用与前述大白蚁属的类似,采食工蚁出巢收集用于培植菌圃的植物材料,是林间的危害虫态,同时,工蚁也有取食菌圃物质的习性(李鸿杰,2014;Lietal., 2015;张新慰,2015),蚁巢内共生的真菌菌圃与培菌白蚁间的食性联系是长期演化的结果。本研究以黑翅土白蚁采食工蚁为对象,研究菌圃、林间采集材料(杂交鹅掌楸树干外表皮)对采食工蚁的引诱活性,分析其食物的组成成分及引诱活性物质的可能来源;同时,通过与白蚁饵料基质常用成分密粘褶菌松木粉培养物的引诱活性进行比较,分析菌圃物质引诱活性的强度。本研究可为进一步揭示培菌白蚁食物采集、加工、分配及其营养生理机制提供参考,也可为开发针对培菌白蚁的高效食性饵剂提供依据。
1 材料和方法
1.1 材料
1.1.1供试昆虫
2015年10月于南京林业大学北大山树木园杂交鹅掌楸Liriodendronchinense×L.tulipifera成熟林(平均胸径约25 cm),用镊子轻轻剥除覆盖在树干表面的泥被,用底部衬有湿滤纸的培养皿收集泥被破损处出现的黑翅土白蚁采食工蚁,培养皿置于遮光处理的塑料桶中带回实验室后,将工蚁置于底部衬有湿滤纸的培养皿内并用保鲜膜包裹保湿,黑暗环境(25℃±1℃、相对湿度75%±5%)静置12 h以上,去除伤残及死亡个体,根据体型大小和体色,选择第5龄采食工蚁静置待用(Soleymaninejadianetal., 2014),环境条件同前。实验前,每5头工蚁一组,放入侧面开孔(孔直径0.6 cm)、底部衬有湿滤纸的塑料培养皿内(直径5.7 cm),分组数量根据不同试验设定。实验时,打开培养皿侧孔,使之与测定装置连接。
1.1.2供试菌圃
林间挖掘黑翅土白蚁蚁巢获得,挖巢时间、地点同1.1.1。本实验采得的蚁巢位于地下约30 cm处,主巢中位,4个副巢紧密排列在其周围。主巢菌圃直径约30 cm,主巢菌圃按上、中、下三个部分分别采集。副巢菌圃(直径约15 cm)为单层结构,采集时未分层次。林间采集的菌圃随即装入标记好的可封口干净塑料袋中,带回实验室后,将菌圃分割成小碎块,用毛笔清除菌圃上的白蚁个体,清除干净的菌圃重新装袋后,-80℃冷冻保存待用。
1.1.3密粘褶菌松木粉培养物
参考丁芳等(2015b)的方法。密粘褶菌菌种购自中国科学院普通微生物菌种保藏管理中心,使用前经PDA平板活化后,接种于PDA平板培养基上,26℃培养14 d,挑取菌丝约0.75 g,无菌水混合至300 mL,将菌丝溶液加入经过灭菌处理的松木粉培养袋(松木粉150 g、12 cm×28 cm聚丙烯塑料袋),25℃培养6周。培养物4℃冷藏备用。
1.1.4白蚁野外采集的植物材料
采集时间、地点同1.1.1。该林分内的黑翅土白蚁主要采集杂交鹅掌楸树干外表皮,林间采集外表皮后,装入干净封口塑料袋,4℃保存备用。使用前风干粉碎。
1.2 方法
1.2.1Y形嗅觉仪结构
玻璃管内径1.0 cm,两臂长15.0 cm,基端长10.0 cm;两臂间夹角45°,两臂与基端夹角157.5°。管道内壁下侧及交叉处为宽0.4 cm的毛玻璃面,便于工蚁正常爬行。两臂端通过三通管与味源瓶相连,三通管出气接口用硅胶管连接并用止水夹封闭。以空气泵驱动的气流经活性炭过滤器和蒸馏水瓶净化后进入味源瓶,携带气味物质进入两臂端,然后经基端进入等候室,各部分之间用聚四氟乙烯管连接。等候室为侧面开孔(直径0.6 cm)、底部衬有湿滤纸的塑料培养皿(直径5.7 cm,高1.5 cm)。孔口与Y形管基端相连(图1)。
1.2.2引诱活性的测定
1.2.2.1 不同材料的引诱活性
测定材料包括主巢菌圃上、中、下层;副巢菌圃;杂交鹅掌楸树干外表皮;密粘褶菌松木粉培养物共6种。各待测材料经室温风干后,适当破碎成小颗粒,称取约3 g装入味源瓶,设自然气流为空白对照。暗室、红光照明、25℃±1℃环境下检测。实验时,调节气体流量使侧臂气流流速300 mL/min,工蚁由等候室出口进入Y形管基端,在感知气味后,开始沿底部的毛玻璃带向臂端方向前行,到达分支角时,白蚁开始晃动触角探测左右两臂气味的区别,选择后沿某一臂继续前进。以工蚁爬过基端5 cm为计时起点,时长5 min。5 min后仍不做出选择则结束观察,记为不选择。工蚁爬过某侧臂5 cm距离以上,视为对该臂味源做出选择。试验共4组,每组5头工蚁,记录处理与对照侧臂中工蚁数量,每测定一次更换一洁净的Y形管,每5次调换味源的位置。每种材料测定试验重复3次,涉及12组60头采食工蚁,6类材料共计72组试验、360头工蚁。每种材料测定完成后,Y形管和味源瓶用95%乙醇进行超声波震荡清洗,150℃高温烘干,晾至常温后重新使用。
1.2.2.2 不同材料引诱活性比较
参考1.2.2.1的结果,选择主巢下层菌圃、副巢菌圃、杂交鹅掌楸树干外表皮、密粘褶菌松木粉培养物为供试材料,分别比较主巢下层菌圃与杂交鹅掌楸树干外表皮、主巢下层菌圃与密粘褶菌松木粉培养物、副巢菌圃与杂交鹅掌楸树干外表皮、副巢菌圃与密粘褶菌松木粉培养物、杂交鹅掌楸树干外表皮与密粘褶菌松木粉培养物的引诱活性。材料来源、处理、引诱活性测定方法同1.2.2.1,每组比较重复3次,共进行60组试验,包括300头工蚁。
1.3 数据统计与分析
按下式计算其趋性反应率和选择系数(张红兵等,2005;丁芳等,2015b)。

选择系数大于0,表示工蚁对该试验材料有趋性,数值越大,表示趋性越强,最大值为1;选择系数若小于0,则表示工蚁对该试验材料有负趋性。活性测定数据结果采用SPSS 19.0和Microsoft Excel 2010统计软件处理分析,采用最小显著差数(LSD)法和新复极差(SSR)法进行单因素方差分析的显著性检验。

图1 Y形嗅觉仪的结构Fig.1 The structure of Y-tube olfactometer注:a,空气泵;b,气流分支件;c,气体流量计;d,空气净化柱;e,加湿瓶;f,样品瓶;g,三通管;h,Y形嗅觉管;i,等候室;j,聚四氟乙烯管;k,硅胶管。Note:a,Air pump;b,Airflow branches;c,Gas flowmeter;d,Air purifier;e,Humidifier;f,Sample bottle;g, Three-way tube;h,Y-tube olfactometer;i,Waiting room;j,PTFE tube;k,Silicon tube.
2 结果与分析
2.1 不同材料对采食工蚁的引诱活性
空白对照时,不同材料对黑翅土白蚁采食工蚁引诱活性如表1。结果表明:副巢菌圃的选择系数最高(>90%),不同材料间选择系数差异显著性检验:杂交鹅掌楸树干外表皮与主巢上层菌圃、主巢下层菌圃差异不显著,其余各材料间差异显著,但均未达极显著水平;各测定材料的选择系数都超过了50%,说明菌圃物质、密粘褶菌松木粉培养物、杂交鹅掌楸树干外表皮对黑翅土白蚁采食工蚁均具有一定的取食引诱活性;不同测定材料趋性反应率大小顺序:密粘褶菌松木粉培养物>副巢菌圃>主巢下层菌圃>主巢中层菌圃>主巢上层菌圃>杂交鹅掌楸树干外表皮。
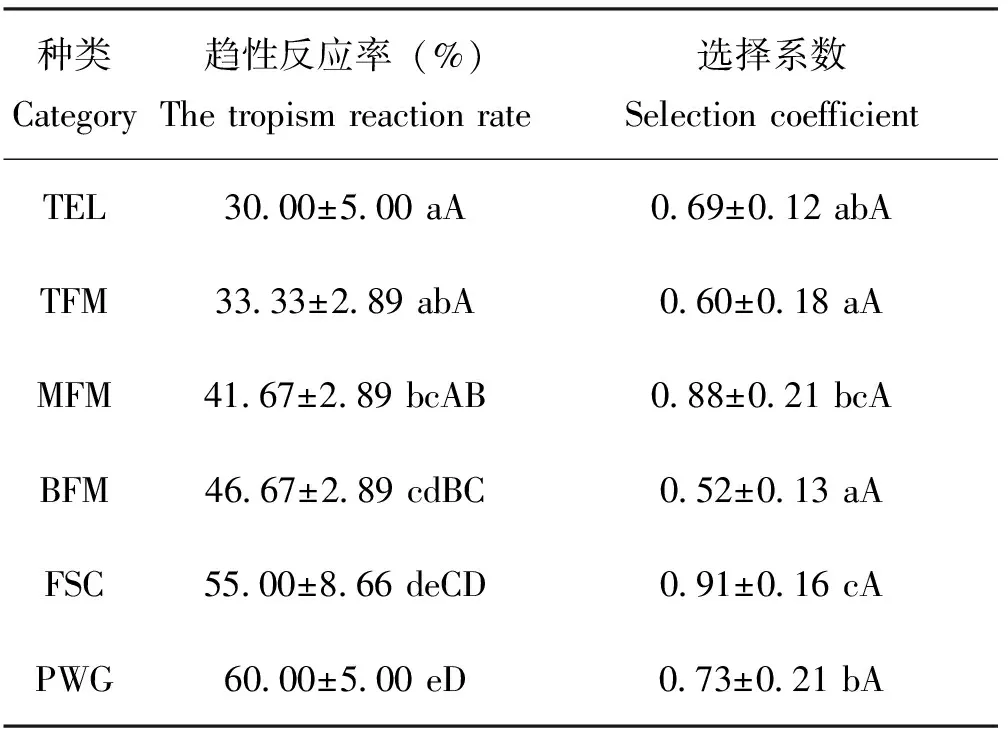
表1 不同材料对黑翅土白蚁采食工蚁的引诱活性
注:(1)表中数据为平均值±标准差,同列数据后标有不同字母表示差异显著,小写字母表示0.05水平上的差异显著水平,大写字母表示0.01水平上的差异显著。(2)TEL,杂交鹅掌楸树干外表皮;TEM,主巢上层菌圃;MFM,主巢中层菌圃;BFM,主巢下层菌圃;FSC,副巢菌圃;PWG,密粘褶菌松木粉培养物。Note: (1) The data in the table are mean±SD and followed by the different small or capital letters in the same column show significant difference at the 0.05 or 0.01 level, respectively. (2) TEL, Trunk’s outer epidermis ofL.chinense×L.tulipifera; TFM, Top layer of fungus garden in main chamber; MFM, Middle layer of fungus garden in main chamber; BFM, Bottom layer of fungus garden in main chamber ; FSC, Fungus garden in satellite chamber; PWG, Pine wood powder infected byG.trabeum.
不同测定材料趋性反应率显著性检验结果较为复杂:密粘褶菌松木粉培养物与副巢菌圃、副巢菌圃与主巢下层菌圃、主巢下层菌圃与主巢中层菌圃、主巢中层菌圃与主巢上层菌圃、主巢上层菌圃与杂交鹅掌楸树干外表皮5个处理组之间差异不显著,其余各组差异显著或极显著。
2.2 不同材料对采食工蚁的引诱活性比较
不同材料引诱活性分组比较结果如表2。5组两两比较测定,各组趋性反应率比较:密粘褶菌松木粉培养物引诱活性高于杂交鹅掌楸树干外表皮(P<0.01);副巢菌圃高于杂交鹅掌楸树干外表皮(P<0.01);主巢下层菌圃高于杂交鹅掌楸树干外表皮(P<0.05);副巢菌圃与密粘褶菌松木粉培养物、主巢下层菌圃与密粘褶菌松木粉培养物2组各自成分间差异均不显著。

表2 不同材料对黑翅土白蚁采食工蚁引诱活性的比较
注:(1)处理组测定材料简写表达同表1。表中数据为平均值±标准差,同一处理标有不同字母表示差异显著,小写字母表示0.05水平上的差异显著水平,大写字母表示0.01水平上的差异显著。(2)数据对应的测定材料种类见数据后括号。Note: (1) Logogram of tested materials are the same as in Table 1. The data in the table are mean±SD and followed by the different small or capital letters in the same treatment show significant difference at the 0.05 or 0.01 level, respectively. (2)The capital letters in parenthesis are homologous to the category of tested materials.
综合表1和表2的结果:(1)主巢下层菌圃、副巢菌圃、密粘褶菌松木粉培养物对采食工蚁有较高的引诱活性,主巢中层菌圃、主巢上层菌圃以及杂交鹅掌楸树干外表皮对采食工蚁的引诱活性相对较低;(2)主巢下层菌圃、副巢菌圃以及密粘褶菌松木粉培养物对采食工蚁的引诱活性相近,没有显著差异,但密粘褶菌松木粉培养物较高,副巢菌圃次之。(3)菌圃物质对采食工蚁的引诱活性比林间采集的食物材料(杂交鹅掌楸树干外表皮)的引诱活性更高。
3 结论与讨论
本文研究表明,黑翅土白蚁共生菌圃对采食工蚁具有明显的引诱活性,根据我们对国内外常用文献数据库检索,以前尚未发现类似报道。引诱活性与菌圃类型、部位有关,主巢下层菌圃和副巢菌圃引诱活性最高,主巢中层菌圃次之,主巢上层菌圃最弱。主巢下层菌圃与副巢菌圃、主巢下层菌圃与主巢中层菌圃、主巢中层菌圃与主巢上层菌圃差异不显著,副巢菌圃和主巢中层菌圃、副巢菌圃与主巢上层菌圃、主巢下层菌圃与主巢上层菌圃之间的差异都达到了极显著水平。从菌圃构建的成分来源和白蚁活动场所特点看,引诱活性物质可能来源于白蚁、共生真菌、植物材料发酵产物,从上述引诱活性分布特征看,食物材料进一步发酵形成的可能性较大。从主巢菌圃引诱活性从上到下逐步增强分析,活性物质含量还与菌圃物质发酵程度有关。本实验的黑翅土白蚁林间采食材料为杂交鹅掌楸树干外表皮,其对采食工蚁也有一定的引诱活性,这似乎为引诱活性物质的食物来源提供了一定支持。黑翅土白蚁菌圃的构建和利用已基本了解,巢内工蚁取食菌丝、分生孢子以及采食工蚁采集的植物材料,排出粪便(初级)作为菌圃培养基(Lietal., 2015;张新慰,2015)。从这一过程看,引诱活性物质的来源也不能排除共生真菌等的影响。确切来源尚待进一步研究。
黑翅土白蚁蚁巢从初建、成长到衰亡的过程中,其结构经历了多种形式的发展过程。3年以上的蚁巢会形成副巢,即“卫星巢”,其内的菌圃也称为“卫星菌圃”,其上有工蚁、兵蚁、幼蚁活动,且有卵存在。空腔巢首先出现在6年以上的“下位寡腔巢”内,其中没有菌圃存在,有报道认为长翅生殖蚁在分飞前大量取食菌圃物质是出现“空腔巢”的重要原因。(蔡邦华等,1965;Chengetal., 2007)。黄翅大白蚁Macrotermesbarneyi相关研究认为采食工蚁取食是“空腔巢”出现的原因(Wangetal., 2009),本文研究表明黑翅土白蚁“空腔巢”形成缘于采食工蚁取食的可能性不能排除。此外,主、副巢菌圃培养基质相同、功能相似,副巢菌圃引诱活性物质含量较高的原因也有待查明。
毒饵诱杀技术因成本低、针对性强、诱杀效果显著等优点已成为白蚁监测和防治的重要措施。从相关报道看,以密粘褶菌松木腐朽物为主要引诱成分的食饵对低等白蚁引诱效果更好,但诱杀高等培菌白蚁效果较差(Ngeeetal., 2004;Kasseneyetal., 2011;Littleetal., 2013;丁芳,2015a)。这可能与培菌白蚁采食巢内菌圃习性有关,其在巢外收集的材料主要用于共生真菌的培养,巢内菌圃较为丰盛时,出巢采集的任务并不迫切,至少不是即采即食的关系。采食工蚁对采集的植物材料、气候以及其他环境因子的变化可能更为敏感,一些细微的变化可能导致出巢采集活动的停止(陈崇征等,2007;王治国和张秀江,2007)。因此,高等培菌对诱饵仿真性等方面的要求可能更高。而以巢内共生菌圃气味物质为基础,研究开发的饵剂,有可能提高对这类白蚁的引诱效果,从而为高等培菌白蚁高效引诱剂的研发提供新的思路。
段金廒等(1999)应用GC-MS从黑翅土白蚁菌圃(含5%白蚁)乙醚提取液中分离出32个成分,鉴定出其中12种的化学结构,分别为己酸、壬酸、N-甲基,N-O-十二烷基-氨基乙酸、十四烷酸、(反)9-十八烯酸、十一烷酸、十七碳烯酸、(反,反)9,1-十八碳二烯酸、十四烷醛、二十五烷、二十二烯酸、三十六烷。薛德钧等(2003)从黑翅土白蚁菌圃中分离出28种水溶性成分,鉴定出其中11种的化学结构,主要为9Z-十八碳烯酸(29.78%)、9,12(Z,Z)-十八碳二烯酸(18.98%)、棕榈酸(18.20%)、十八碳酸(8.68%)、胆固醇(5.79%),此外为微量成分己醛、乙酸正丁酯、2-三氯甲基-2-丙醇、9(11)-脱氢麦角甾醇苯甲酸酯、β-谷甾醇和谷甾-3,5-二烯-7-酮。Nyegue 等(2003)从喀麦隆一种大白蚁Macrotermesnatalensis共生蚁巢伞Termitomycesshimperi子实体中分离鉴定出24种挥发性化合物,主要为1-辛烯-3-醇(34.3%)、苯乙醇(19.5%)、正己醛(11.8%)、2-己醇(7.4%)、3-己醇(3.4%)、辛烷-3-酮(2.1%)、(E,E)2,4-壬二烯醛(1.3%)、1-辛烯-3酮(1.0%)、(E)-2-辛烯醛(0.8%),其他为微量成分。上述已报道黑翅土白蚁菌圃以及Termitomycesshimperi子实体中挥发性成分与菌圃挥发性成分的关系,及其与白蚁之间的信息联系机制值得进一步研究。
韩美贞和严峰(1980)曾比较家白蚁有翅成虫、巢片、密粘褶菌、茯苓、杂色云芝的染菌腐木粗提液的踪迹和引诱活性,证明引诱活性和踪迹活性间存在较大差异。研究较为系统的是密粘褶菌腐朽木引诱白蚁取食的机制,Smythe 等(1967)从密粘褶菌腐木提取物中发现了与白蚁腹板腺相同性质的踪迹活性物质。Ohmura 等(1995)鉴定这种踪迹活性物质是(Z,Z,E)-3,6,8-十二碳三烯-1-醇。丁芳等(2015b)研究了密粘褶菌松木粉培养物对黑翅土白蚁采食工蚁的引诱活性和踪迹活性,结果表明除踪迹活性外,密粘褶菌松木粉培养物对黑翅土白蚁采食工蚁还具有较高的引诱活性,乙酸乙酯和正己烷均有较高的引诱活性组分提取效果,从而为常用密粘褶菌饵剂化学增效提供了思路。本文的研究虽然证实共生菌圃中含有较高的引诱活性组分,但组分的理化性质、组成特点、来源等尚需要进一步研究查明。此外,共生菌圃对采食工蚁是否具有踪迹活性,也需要进一步研究查明。
References)
Aanen DK, Ros V, de Fine Licht H,etal. Patterns of interaction specificity of fungus-growing termites andTermitomycessymbionts in South Africa [J].BMCEvolutionaryBiology, 2007, 7 (1): 115.
Apolinário FE, Martius C. Ecological role of termites (Insecta, Isoptera) in tree trunks in central Amazonian rain forests [J].ForestEcologyandManagement, 2004, 194 (1-3): 23-28.
Cai BH, Chen NS, Chen AG,etal. Architecture and development of the termitarium ofOdontotermesformosanus(Shiraki) [J].ActaEntomologicaSinica, 1965, 14 (1): 53-70. [蔡邦华, 陈宁生, 陈安国, 等. 黑翅土白蚁Odontotermesformosanus(Shiraki) 的蚁巢结构及其发展[J]. 昆虫学报, 1965, 14 (1): 53-70]
Chen CZ, Jiang XJ, Deng Y. Effects of environmental factors on termites harmfulness toEucalyptus[J].GuangxiForestryScience, 2007, 36 (3): 121-123. [陈崇征, 蒋学建, 邓艳. 桉树白蚁危害与环境因子的关系研究 [J]. 广西林业科学, 2007, 36(3): 121-123]
Cheng ML, Mo JC, Deng TF,etal. Biology and ecology ofOdontotermesformosanusin China (Isoptera: Termitidae) [J].Sociobiology, 2007, 50 (1): 45-461.
Ding F, Ji BZ, Liu SW,etal. Food selection of termites [J].ChineseAgriculturalScienceBulletin, 2015a, 31 (2):166-173. [丁芳, 嵇保中, 刘曙雯, 等. 白蚁的食物选择[J]. 中国农学通报, 2015a, 31 (2):166-173]
Ding F, Ji BZ, Liu SW,etal. The biological activity of pine wood powder infected withGloeophyllumtrabeumonOdontotermesformosanus(Shiraki) [J].ChineseJournalofAppliedEntomology, 2015b, 52 (3): 736-742. [丁芳, 嵇保中, 刘曙雯, 等. 密粘褶菌松木粉培养物对黑翅土白蚁的生物活性[J]. 应用昆虫学报, 2015b, 52 (3): 736-742]
Duan JA, Huang WZ, Yang D,etal. Analysis of ethereal extracts fromOdondotermesformosanusand fungus garden [J].ZhongYaoCai, 1999, 22 (9): 437-438. [段金廒, 黄文哲, 杨东, 等. 黑翅土白蚁及其菌圃的醚溶性成分[J]. 中药材, 1999, 22 (9): 437-438]
Han MZ, Yan F. A preliminary report on the comparative tests of termite trail-following pheromone analogues from fungus-infected wood [J].ActaEntomologicaSinica, 1980, 23 (3): 260-264. [韩美贞, 严峰. 白蚁跟踪信息素及其类似物的活性比较试验初报[J]. 昆虫学报, 1980, 23 (3): 260-264]
Hennon P, Woodward B, Lebow P. Deterioration of wood from live and dead Alaska yellow-cedar in contact with soil [J].ForestProductsJournal, 2007, 57 (6): 23-30.
Hinze B, Crailsheim K, Leuthold RH. Polyethism in food processing and social organisation in the nest ofMacrotermesbellicosus(Isoptera, Termitidae) [J].InsectesSociaux, 2002, 49 (1): 31-37.
Kasseney BD, Deng T, Mo J. Effect of wood hardness and secondary compounds on feeding preference ofOdontotermesformosanus(Isoptera: Termitidae) [J].JournalofEconomicEntomology, 2011, 104 (3): 862-867.
Kirker GT, Wagner TL, Diehl SV. Relationship between wood-inhabiting fungi andReticulitermesspp. in four forest habitats of northeastern Mississippi[J].InternationalBiodeterioration&Biodegradation, 2012, 72: 18-25.
Li HJ. The Mechanism of Lignocellulose Degradation in the Mutualism System ofOdontotermesformosanusand Associated Microorganism [D]. Hangzhou: Zhejiang University, 2014. [李鸿杰. 黑翅土白蚁及其微生物协同降解木质纤维素机制研究[D]. 杭州: 浙江大学, 2014]
Li HJ, Yang MY, Chen YE,etal. Investigation of age polyethism in food processing of the fungus-growing termiteOdontotermesformosanus(Blattodea: Termitidae) using a laboratory artificial rearing system [J].JournalofEconomicEntomology, 2015, 108 (1): 266-273.
Li SJ, Zhu TH. Macro-fungi and standing tree rot[J].JoumalofSichuanForestryScienceandTechnology, 2011, 32 (1): 59-64. [李姝江, 朱天辉. 2011. 大型真菌与立木腐朽[J]. 四川林业科技, 2011, 32 (1): 59-64]
Little NS, Blount NA, Londo AJ,etal. Preference of formosan subterranean termites for blue-stained southern yellow pine sapwood [J].JournalofEconomicEntomology, 2012a, 105 (5): 1640-1644.
Little NS, Riggins JJ, Schultz TP,etal. Feeding preference of native subterranean termites (Isoptera: Rhinotermitidae:Reticulitermes) for wood containing bark beetle pheromones and blue-stain fungi [J].JournalofInsectBehavior, 2012b, 25 (2): 197-206.
Little NS, Schultz TP, Diehl SV,etal. Field evaluations of subterranean termite preference for sap-stain inoculated wood [J].JournalofInsectBehavior, 2013, 26 (5): 649-659.
Ngee PST, Yoshimura T, Lee CY. Foraging populations and control strategies of subterranean termites in the urban environment, with special reference to baiting [J].JapaneseJournalofEnvironmentalEntomologyandZoology, 2004, 15 (3): 197-215.
Nobre T, Rouland-Lefèvre C, Aanen DK. Comparative biology of fungus cultivation in termites and ants.In: Bignell DE, Roisin Y, Lo N. Biology of Termites: A Modern Synthesis [C]. Berlin: Springer Verlag Press, 2011: 193-210.
Nyegue M, Phazollo PHA, Bessière JM,etal. Volatile components of freshPleurotusostreatusandTermitomycesshimperifrom Cameroon [J].JournalofEssentialOil-bearingPlants, 2003, 6 (3):153-157.
Ohmura W, Tokoro M, Tsunoda K. Termite trail following substances produced by brown-rot fungi [J].MaterialandOrganismen, 1985, 29 (2): 133-146.
Perry DH, Lenz M, Watson JAL. Relationships between fire, fungal rots and termite damage in Australian forest trees [J].AustralianForestry, 1985, 48 (1): 46-53.
Silva CA, Monteirob MBB, Brazolin S,etal. Biodeterioration of brazilwoodCaesalpiniaechinataLam. (Leguminosae-Caesalpinioideae) by rot fungi and termites[J].InternationalBiodeteriorationandBiodegradation, 2007, 60 (4): 285-292.
Smythe RV, Oppel HC, Lipton SH,etal. Chemical studies of attractants associated withReticulitermesflavipesandR.Virginicus[J].JournalofEconomicEntomology, 1967, 60 (1): 228-233.
Soleymaninejadian E, Ji BZ, Liu SW,etal. Foraging polyethism inOdontotermesformosanusShiraki [J].JournalofAdvancedAgriculturalTechnologies, 2014, 1 (1): 32-42.
Wang PF, He J, Zhou W,etal. A survey on the studies ofTermitomyces[J].MicrobiologyChina, 2012, 39 (10): 1487-1498. [王鹏飞, 何隽, 周文, 等. 蚁巢伞属真菌研究进展[J]. 微生物学通报, 2012, 39 (10): 1487-1498]
Wang ZG, Zhang XJ. The Fauna Orthopteroidea of Henan[M]. Zhengzhou: Henan Science and Technology Press, 2007: 63. [王治国, 张秀江. 河南直翅类昆虫志[M]. 郑州: 河南科学技术出版社, 2007: 63]
Wang, ZY, Mo JC, Lu YJ. Biology and ecology ofMacrotermesbarneyi(Isoptera: Termitidae) [J].Sociobiology, 2009, 54 (3): 777-785.
Xue DJ, Zhou H, Zhang M,etal. Study of water-soluble compounds from fungus garden ofOdontotermesformosanus[J].JouranlofChineseMedicinalMaterials,2005, 28 (10): 873-875. [薛德钧, 周慧, 张敏, 等. 白蚁菌圃的水溶性化学成分研究[J]. 中药材, 2005, 28 (10): 873-875]
Zhang HB, Li XY, Dai HG,etal. The olfactory responses ofCoptotermesformosanusandReticulitermesflavicepsto three baited insecticides [J].ChineseBulletinofEntomology, 2005, 42 (3): 298-301. [张红兵, 李小鹰, 戴华国, 等. 台湾乳白蚁和黄胸散白蚁对三种饵剂的嗅觉行为反应[J]. 昆虫知识, 2005, 42 (3): 298-301]
Zhang XW. Researches on Feeding Differentiation among Castes ofOdontotermesformosanus(Shiraki) [D]. Nanjing: Nanjing Forestry University, 2015. [张新慰. 黑翅土白蚁不同品级食性分化研究[D]. 南京: 南京林业大学, 2015]
AttractiveactivityoffungusgardentoOdontotermesformosanus(Shiraki) (Isoptera:Termitidae)foragers
WANG Ya-Zhao1, JI Bao-Zhong1*, LIU Shu-Wen2*, DING Fang1, XU Li-Jun1
(1. Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, College of Forestry, Nanjing Forestry University, Nanjing 210037, China; 2. The Administration Bureau of Dr. Sun Yat-sen’s Mausoleum, Nanjing 210014, China)
Odontotermesformosanusis an important fungus-growing termite. Their foragers feed on the material consisting of fungus garden. Outside the nest, foragers collect tree bark or other plant materials to use them as the culture medium of fungus garden. In this paper, we investigated the attractive activity of fungus garden and field-collected materials (trunk’s outer epidermis ofLiriodendronchinense×L.tulipifera, TEL) toO.formosanusforagers. Meanwhile, the pine wood powder infected byGloeophyllumtrabeum(PWG), a kind of frequently used termite bait material, was taken as comparation, and its attractive activity was also tested. Results in this paper may provide references for the further researches on the mechanism of feeding habits and nutritional physiology inO.formosanusor other fungus-growing termites. Attractive activity of six kinds of materials toO.formosanusforagers was tested respectively by using Y-tube olfactometer. Tested materials included the top layer of fungus garden in main chamber (TFM), the middle layer of fungus garden in main chamber (MFM), the bottom layer of fungus garden in main chamber (BFM), the fungus garden in satellite chamber (FSC), TEL and PWG. Afterwards, attractive activity was compared in pairwise comparison. The treatments were arranged as follows: BFM to TEL, BFM to PWG, FSC to TEL, FSC to PWG, and TEL to PWG. It was showed that:(1) In the condition of blank control, orientational response rates in different tested materials were ranked as following order: PWG>PSC>BFM>MFM>TFM>TEL. The significance test of orientational response rate in different tested materials showed that there were not significant differences between PSC and PWG, PSC and BFM, BFM and MFM, MFM and TFM, TFM and TEL, respectively. Meanwhile, there were significant or extremely significant differences in other treatments. (2) In the condition of pairwise comparison, the attractive activity of PWG was higher than that of TEL (P<0.01); FSC was higher than TEL (P<0.01); BFM was higher than TEL (P<0.05); No significant differences existed between FSC and PWG or BFM and PWG. (3) The fungus garden ofO.formosanushas obviously attractive activity to its foragers. The intensity of attractive activity related to the location of tested material in fungus garden or the type of fungus garden. (4) BFM and FSC ranked highly in attractive activity, they were quite similar with that of PWG. On the contrast, the MFM, TFM, TEL showed low attractive activity. The attractive activity of fungus garden substance was much higher than that of field-collected materials. Some subjects were put forward for further researches, such as the physicochemical property, composition, and origin of attractive components in the symbiotic fungus garden.
Odontotermesformosanus; foragers; fungus garden; attractive activity
王亚召,嵇保中,刘曙雯,等.黑翅土白蚁菌圃对采食工蚁的引诱活性[J].环境昆虫学报,2017,39(5):1161-1168.
Q968.1
A
1674-0858(2017)05-1161-08
南京林业大学南方现代林业协同创新中心资助项目;高等学校博士学科点专项科研基金项目(20123204110001);江苏省自然科学基金项目(BK2012816);南京市建委、中山陵园管理局资助课题(201409);江苏高校优势学科建设工程资助项目(164010641);江苏省普通高校学术学位研究生科研创新计划项目(KYLX150911)
王亚召,男,1991年生,河南汝州人,硕士研究生,研究方向为昆虫生理生化,E-mail: 775159143@qq.com
*通讯作者Corresponding author,E-mail: jbz9885@njfu.edu.cn; liushuwen2006@163.com
Received: 2016-05-16;接受日期Accepted: 2016-07-02
