内隐情绪加工对返回抑制效应的调节 *
2017-11-02吴晓刚李乐源潘发达
吴晓刚 施 亮 李乐源 潘发达
(1 南通大学教育科学学院, 南通 226007)
(2 苏州大学心理学系, 心理与行为科学研究中心, 苏州 215123)
内隐情绪加工对返回抑制效应的调节 *
吴晓刚1,2施 亮1李乐源1潘发达1
(1 南通大学教育科学学院, 南通 226007)
(2 苏州大学心理学系, 心理与行为科学研究中心, 苏州 215123)
为探讨内隐情绪加工是否会对返回抑制产生影响,采用线索靶子范式,以情绪性面孔图片作为目标,要求被试完成定位任务。结果发现,行为数据和脑电数据均显示了显著的返回抑制效应;N1成分上情绪效价和线索有效性的主效应及交互作用均显著,有效线索条件下情绪目标诱发的波幅显著大于中性目标;N170成分上线索有效性主效应显著,相较于N1成分,N170成分在有效线索上波幅更小,在无效线索上波幅更大。实验结果表明返回抑制效应受到内隐情绪加工的调节,提示该效应来源于早期的知觉抑制,支持知觉抑制理论。
返回抑制,内隐情绪加工,事件相关电位。
1 问题提出
返回抑制(inhibition of return, IOR)是指个体对先前注意过地方反应变慢的一种心理现象(Posner & Cohen, 1984)。自IOR被发现以来,一直是注意研究领域的热点(张阳, 彭春花, 孙洋, 张明, 2013)。通过对先前注意过的地方打上抑制标签,提高对未注意过地方的搜索效率,返回抑制这种抑制搜索无关位置的能力极大地促进了视觉空间搜索效率。这种具有认知加工灵活性和适应性的机制被称为“视觉搜索加速器”,能有效避免重复搜索从而提高搜索效率(Klein & MacInnes,1999; Klein, 1988)。根据当前研究,IOR被认为是人类进化而来的一种适应机制,通过减少对无关刺激的注意时间从而提高有机体对环境的搜索能力,并从其他空间位置侦查潜在的意义事件(Sapir, Soroker, Berger, & Henik, 1999)。这样的一种机制不仅能帮助提高视觉搜索,对于生态环境中的需求变化,也能为个体的决策和行为调节提供帮助(Ivanoff & Taylor, 2006)。
对于返回抑制是如何产生的这一问题,一直存在着两种不同的理论争执,即知觉抑制理论和反应抑制理论。知觉抑制理论认为,对先前注意过地方的抑制主要是通过对早期知觉加工施以影响,这一理论得到了很多研究的支持。Klein和Dick(2002)使用RSVP范式,这种范式很好的解决了返回抑制中知觉和反应的关系。先前关于返回抑制的研究都使用了快速侦查范式,而他们的实验任务则是让被试尽可能正确的作出反应而不要求速度,并假设如果返回抑制是一种反应抑制效应,那么在有效线索和无效线索上差异不显著。结果发现,被试对有效线索上的刺激有更多的错误反应,这说明返回抑制不是一种反应抑制现象。最近的一项研究发现,在外源性线索化下的MRT任务和POT任务中,相比无效线索,有效线索上出现了知觉合并,即在有效线索上的反应时显著慢于无效线索上的反应时(Krüger,MacInnes, & Hunt, 2014)。在认知神经科学领域,知觉抑制理论也得到了支持。Prime和Ward(2004)的一项ERP研究发现,IOR的产生是来自知觉过程的变化,和早期知觉加工有关,位于枕部的P1/N1成分在有效线索上比无效线索上诱发出了更小的ERP波幅,说明返回抑制的产生不是源于对个体反应过程的抑制。另外一项ERP研究也发现,P1和N1在有线线索上的波幅显著小于无效线索上的波幅,说明IOR在有效线索上反应变慢是因为最后的反应受到了前期知觉的影响(Prime &Jolicœur, 2009)。反应抑制理论则是认为返回抑制的产生不是通过对早期知觉加工过程的影响,而是抑制晚期的运动加工。Taylor和Klein(1998)认为,IOR的产生是眼球运动对线索的激活从而产生了一种机体反应偏向,表现为个体不愿对有效线索位置的刺激做出反应的结果。另外几项研究对知觉抑制理论提出了挑战,例如返回抑制的效应量大小会受到反应选择物的数量(Adam,O’Donnell, & Pratt, 2005)、置于键盘的非反应手(Ivanoff & Klein, 2001)以及刺激-反应的概率(Ivanoff & Klein, 2004; Prime & Jolicœur, 2009)的影响。因此返回抑制被认为是受到了机体加工和反应选择变化的影响,而不是受到知觉加工过程的影响。
返回抑制作为一种“视觉搜索加速器”被认为是发生在知觉层面的,属于知觉层面的抑制过程(Stoyanova, Pratt, & Anderson, 2007)。此外,随着认知神经科学的兴起,越来越多的研究关注生物学意义的刺激对IOR影响的神经机制。社会或生物意义刺激如情绪信息,个体对其有着特殊的敏感性,具有情绪意义的刺激会占用更多的注意资源(黄宇霞, 罗跃嘉, 2009),个体对其反应也会加强(Lang, 1995)。尤其是负性情绪信息,个体给予更多注意被认为是一种适应环境的表现(彭晓哲, 周晓林, 2005)。近来,有研究者指出,如果IOR是一种提高搜索效率的进化机制,它的效应量就受到生物学意义刺激的调节(Silvert &Funes, 2016)。本文通过梳理已有的文献,发现情绪刺激对IOR主要有两方面的影响。
首先,特殊被试群体是影响IOR的重要因素。国内和国外学者都发现,高低特质焦虑个体(Fox, Russo, Bowles, & Dutton, 2001)、高低状态焦虑个体(Pérez-Dueñas, Acosta, & Lupiáñez, 2014)及抑郁个体(戴琴, 冯正直, 2009)的IOR能力普遍不足,他们对情绪性刺激的敏感度显著高于普通个体,并且注意不容易从情绪刺激上转移,导致IOR效应量显著减小或消失。其次,对于普通个体,情绪信息和IOR的各自加工阶段是重要的影响因素。在经典的空间线索范式中,对外源性线索的注意是一种自下而上的注意,是对感知觉过程的反应(Klein & Dick, 2002;Hopfinger & West,2006;Sapir, Jackson, Butler, Paul, & Abrams,2014)。另一方面,Bernat,Bunce和Shevrin(2001)曾指出,阈上和阈下的视觉情绪刺激加工有着不同的神经通路和反应特征。王敬欣、贾丽萍、白学军和罗跃嘉(2013)采用空间线索范式,要求被试对情绪刺激完成辨别任务,结果在行为和脑电数据中均没有发现IOR和情绪加工的交互作用,而他们采用定位任务的研究却在行为结果中发现了显著的交互作用(王敬欣, 贾丽萍, 张阔,张赛, 2013)。辨别任务更多的是涉及对情绪信息的意识性判断;而定位任务则要求个体的注意始终保持在屏幕的中央,对情绪刺激只做位置的辨别,此时对情绪信息的加工更多地涉及无意识加工。而空间线索范式中的IOR通常发生在感知觉阶段,涉及的是无意识加工。所以,当两种加工都涉及同一通道时,容易出现对注意资源的竞争,产生交互作用(王敬欣, 贾丽萍, 张阔等, 2013;Pan, Wu, Zhang, & Ou, 2017)。邓晓红、张德玄、黄诗雪、袁雯和周晓林(2010)采用阈上和阈下情绪线索的研究也发现了相同的结果。在非空间IOR和情绪加工的研究中,当研究者要求个体进行情绪辨别任务时,发现情绪加工不会对IOR效应量产生显著影响(Poliakoff et al., 2016)。但也有一些研究结果不一致,如Taylor和Klein(2005,2008)采用拼凑面孔在不同的实验任务中发现IOR并不总是稳定;而采用面孔简图的研究却发现IOR不受影响,表现出对生物学刺激的“盲目性”(王丽丽, 罗跃嘉, 郭亚桥, 张庆林, 2010)。需要注意的是,这些新异的刺激材料由于物理结构的特殊性,对吸引注意均会存在不同程度的影响。
综上,对情绪信息的敏感性是人类进化过程中形成的一种预警机制,这种机制有利于人类趋利避害,更好地生存与发展。研究表明,人脑在加工的早期阶段就能对情绪刺激做出反应,这种自下而上的知觉分析会引起意识的或无意识的情绪反应(van Honk, Tuiten, de Haan, vann de Hout, &Stam, 2001)。而返回抑制现象的很多研究结果显示IOR也是一种自下而上的反射性行为,其内在机制为大脑对注意过的地方的知觉抑制(Klein &MacInnes, 1999; Stoyanova et al., 2007;王敬欣, 贾丽萍, 白学军等, 2013)。根据这一理论,如果在先前注意过的地方出现情绪性刺激,那么大脑对该位置的知觉抑制将会和情绪的优先加工之间产生竞争,从而发生相互影响。因此,本研究对经典的线索靶子范式进行适当改进,在目标判断阶段单侧呈现情绪图片,被试完成内隐情绪判断任务(图片位置判断),以此考察情绪的内隐加工是否会对返回抑制效应产生影响。研究材料采用真人情绪面孔,相较于面孔简图、拼凑面孔或非面孔(Taylor & Klein, 2005, 2008)人脸表情能传达出更多生物和社会信息(梦向丽, 王萌华, 王玉平,2006),研究的生态效度更好。
2 实验方法
2.1 被试
有偿招募在校大学生13名(5男,8女),平均年龄22岁。被试均为右利手,视力或矫正视力正常,无身心疾病,且之前均未参与过类似实验。实验开始前先签署知情同意书,实验结束后给予一定的实验报酬。
2.2 实验材料
图片来源于中国情感图片系统(CFAPS)(龚栩, 黄宇霞, 王妍, 罗跃嘉, 2011),采用正性、负性和中性面孔图片。正性和负性各4张,中性8张,其中正性选用高兴面孔负性选用悲伤面孔,中性选用平静面孔1,男女各占50%。用图像软件PhotoShop 6.0将各图片大小、亮度、对比度调成一致,图片大小为260×300像素。
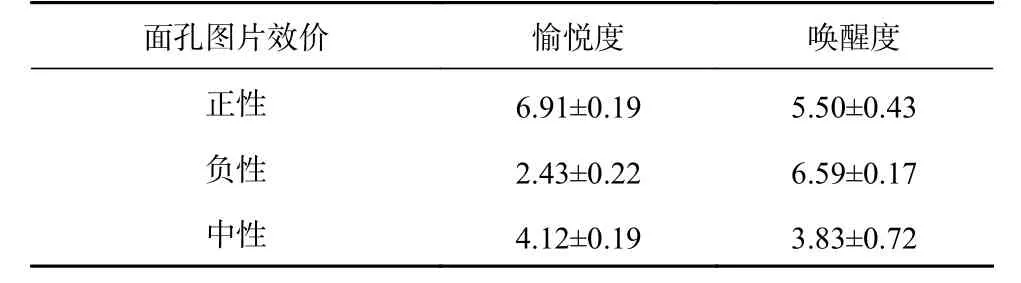
表 1 不同效价面孔的愉悦度和唤醒度(M±SD)
2.3 实验设计和实验程序
采用2×2的被试内设计,自变量为线索有效性(有效线索/无效线索)和目标情绪效价(情绪性/中性),因变量为行为反应时和ERPs的波幅。
实验在隔音的房间中进行,实验开始前先向被试解释脑电的基本原理及签署知情同意书,消除被试的紧张心理。然后让被试坐在电脑面前,给被试带电极帽并开始打导电膏。阻抗降至5 kΩ以下,让被试调整到舒适坐姿,正式开始实验。
实验采用经典的线索靶子范式,线索和靶子之间的SOA在1000-1100 ms内变化(Prime &Ward, 2006)。实验程序如图1,每次测试开始前,屏幕中央会出现三个同样大小的矩形框,中间矩形框中有一个“+”,实验指导语要求被试始终注意中间矩形框中的“+”或圆环。750-850 ms后,左边或右边的矩形框中会出现一个圆环作为线索提示,持续200 ms。300 ms后,中间矩形框中的“+”会被圆环所替代,200 ms会又变回“+”。间隔300-400 ms后,左边或者右边的矩形框中出现目标图片持续 200 ms,当图片一出现,要求被试准确且快速地判别图片出现的位置。如果出现在左边,按“Z”键,出现在右边按“M”键。当提示线索和目标出现在同侧,则为有效线索,出现在异侧则为无效线索。实验由练习实验和正式实验组成,练习实验有32个trial,当被试的正确率达到80%以上时方可进行正式实验。正式实验分为两个block,每个block有96个trial,两个block之间有2分钟的休息时间。正式实验中,有效线索和无效线索各占50%,正性和负性面孔作为情绪性面孔和中性面孔在有效/无效线索中各占50%,男女比例各占一半,且全部随机出现。实验过程中要求被试尽量不要有皱眉、吞咽等动作,被试可以自由眨眼,但要求始终注视中心注视点。被试距电脑屏幕约65cm,采用黑色背景,其中注视点大小为0.5°(水平)×0.5°(垂直)视角,圆环大小为2°(水平)×2°(垂直)视角,方框大小为2.34°(水平)×3.36°(垂直)视角,三个方框的视角范围约为±5°。
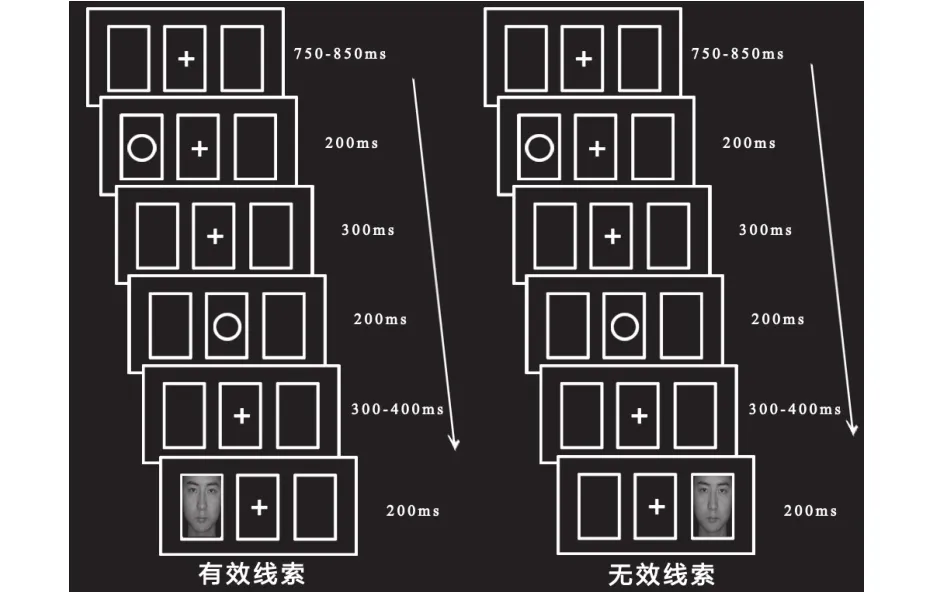
图 1 实验流程图
2.4 脑电数据采集与分析
2.4.1数据采集
采用E-Prime软件编程并记录被试的行为数据。EEG数据采集仪器为美国Neuroscan公司脑电记录系统Scan4.5,采用64导电极帽,头皮电阻降至5 kΩ以下时,开始记录。以左侧乳突作为参考电极,离线分析时转换为双侧乳突的平均为参考。采样频率为500 HZ/导,同时记录被试的水平眼电和垂直眼电,AC=0.05-100 Hz,进行连续采样。
2.4.2数据分析
本研究主要分析ERPs的早期成分。根据文献和总平均图,选取8个电极位置记录的ERP波形用于统计分析,分别为Po5、Po6、O1、O2、Poz、Oz、P7、P8;确定时间窗为P1:80 ms-140 ms,N1:140 ms-180 ms,N170:180-240 ms,其中P1和N1统计Po5、Po6、O1、O2、Poz、Oz这六个点,N170统计P7、P8两个点。先将记录的数据导入Curry7中,首先进行基线校准,type类型选择“Constant”,然后去除眼动伪迹和其他伪迹对脑电的影响,垂直眼电时域选择-200-200 ms,波幅大于±100μV视为伪迹;以图片刺激前200 ms为基线,分析时间窗为-200-800 ms进行分段和叠加平均,并合并行为数据,剔除错误反应对应的脑电,可得到不同水平下的脑电波形;最后将参考电极转换为M1和M2平均为参考,并进行低通30 Hz (24 dB/oct)的数字滤波。将得到的脑电进行峰值探测,并导出数据。
3 实验结果
3.1 行为数据结果
首先剔除反应时小于100 ms和大于1000 ms的极端数据以及正负三个标准差之外的数据和错误反应数据,剔除正确率低于80%被试的数据,然后各种条件下的反应时进行重复测量方差分析。
两因素重复测量方差分析结果显示,线索类型主效应显著,F(1, 12)=54.74,p<0.001,η2=0.82,有效线索反应时(363.52 ms)显著长于无效线索反应时(335.23 ms);情绪效价主效应不显著,F(1, 12)=4.41,p>0.05;交互作用不显著,F(1, 12)=0.54,p>0.05。
3.2 脑电数据结果
根据脑电结果分析,表明:P1成分的线索类型、电极点、情绪效价主效应不显著,各个交互作用也不显著。N1成分的线索类型主效应显著,F(1, 11)=29.11,p<0.001,η2=0.73有效线索上的波幅(–4.12μV)显著大于无效线索上的波幅(–2.69μV);情绪效价的主效应显著,F (1,11)=8.05,p<0.05,η2=0.42,情绪性图片的波幅(–4.16μV)显著大于中性图片的波幅(–2.64μV);电极点主效应显著,F(5, 55)=9.30,p<0.001,η2=0.81,Po5、Po6波幅显著大于Poz、O1、Oz显著大于O2波幅;线索类型和情绪效价交互作用显著,F(1, 11)=7.85,p<0.05,η2=0.42,进一步简单效应检验发现,有效线索条件下情绪性图片(–5.24μV)和中性图片(–2.99μV)诱发的ERP波幅存在显著差异,F(1, 11)=8.42,p<0.05;对于其他交互作用不做分析。N170的线索类型主效应显著,F(1,11)=17.14,p<0.05,无效线索上的波幅显著大于有效线索上的波幅;情绪效价、电极点主效应不显著,各个交互作用均不显著。
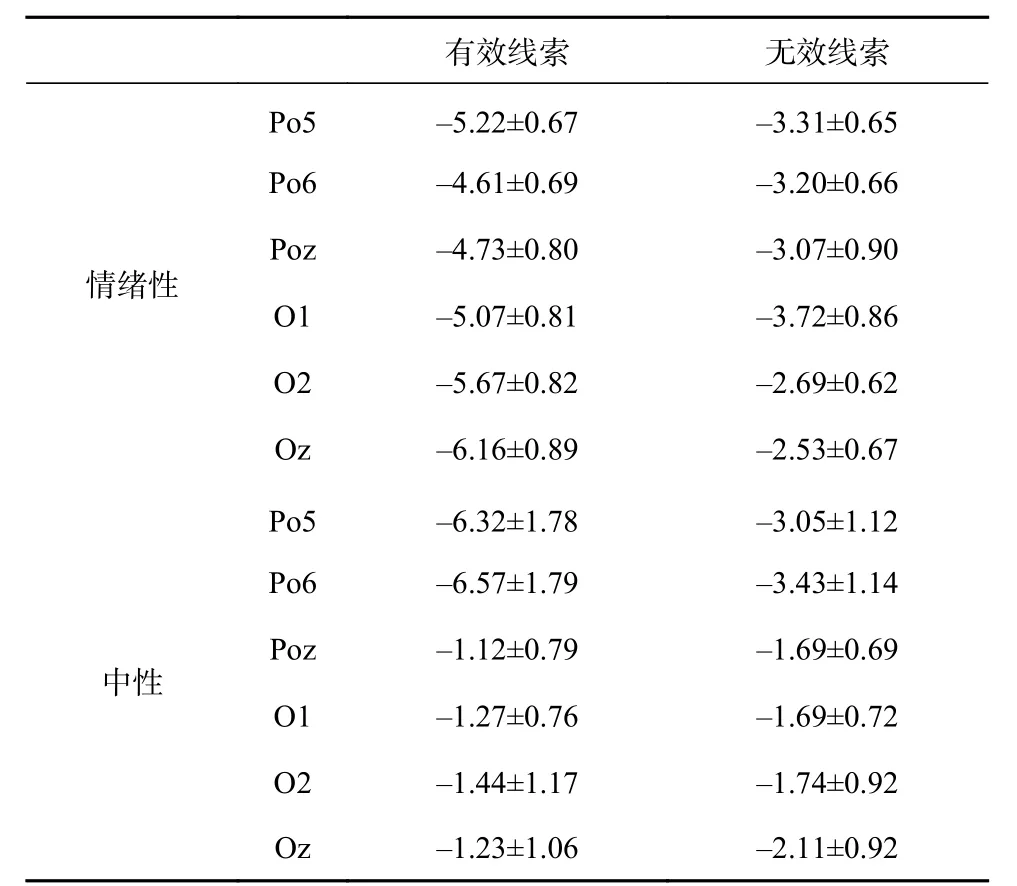
表 2 N1脑电数据波幅(M±SD μV)

表 3 N170脑电数据波幅(M±SD μV)
4 讨论
本研究采用真人情绪面孔,探讨了内隐情绪加工对返回抑制的影响。结果发现,在行为数据和脑电数据上都出现了显著的返回抑制效应,实验结果提示我们返回抑制来源于早期的知觉抑制,内隐情绪加工对返回抑制效应具有调节作用。
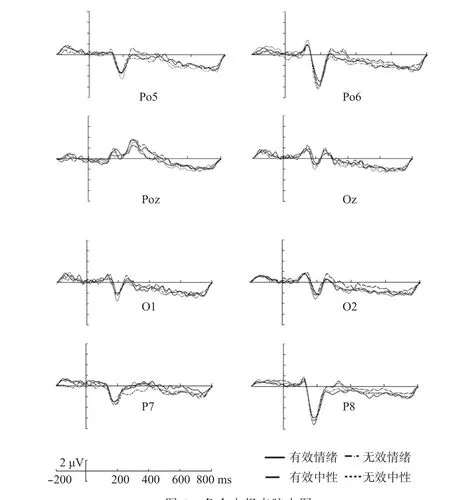
图 2 各个电极点脑电图
首先,脑电结果在P1上没有差异。Hopfinger和Mangun(2001)在使用探测光点位置的定位任务中发现,在短SOA时发现了增强的P1效应,而长SOA(566-766 ms)时虽然出现了返回抑制效应,但P1效应消失了,他们认为这种反射性注意只暂时对视觉加工过程起促进作用。McDonald,Ward和Kiehl(1999)发现在定位任务中,返回抑制效应的出现伴随着P1成分的减弱。研究者认为这是由于实验任务而产生的差异,Prime和Ward(2004)发现有效提示下波幅明显减小,并认为这种效应可能与辨别任务有关;先前的研究发现N1成分与辨别过程有关(Vogel & Luck,2000),而N1成分的注意加强只有在辨别任务中才能体现(Mangun & Hillyard, 1991)。近来一项ERP研究中发现,有效线索与无效线索条件下早期的成分N1和P1上差异不显著,而随后的一个负性成分(Nd),差异波激活了额部脑区,研究者认为知觉抑制和反应抑制均对返回抑制产生了影响(王丽丽, 郭亚桥, 邱江, 张庆林, 2010)。我们的研究发现,有效线索波幅在N1上显著大于无效线索,而在N170上有效线索波幅显著小于无效线索波幅。作为“视觉搜索加速器”的返回抑制通过对早期知觉进行抑制,使个体的注意更快速的转向未搜索过的位置,从而提高了视觉搜索效率。所以,我们认为这种效应是通过对早期知觉抑制的结果。这一结果和先前结果类似,早期的知觉成分P1和N1在有效线索上诱发出了更小的波幅(Prime & Ward, 2004; Prime & Jolicœur, 2009)。返回抑制受到早期注意的影响,由于实验任务的不同,P1和N1的研究结果也有所不同。罗琬华、曾敏和李凌(2003)的研究发现,当要求被试完成觉察任务时,有效线索下P1值更小,N1值更大了。在最近的一项ERP研究中也得到了一致的结果,当要求被试对目标位置的情绪刺激进行效价判断任务时,与无效线索相比,有效线索上的P1波幅更小,N1波幅更大(王敬欣, 贾丽萍, 白学军等,2013)。这两项结果一方面说明了返回抑制来自于早期的注意抑制,另一方面说明N1成分并不一定是存在于辨别任务中。我们认为N1和N170的这种变化与早期知觉加工过程相关,返回抑制来源于早期的注意抑制,并不是来源于反应抑制。所以本研究为知觉抑制说提供了证据。
其次,我们在早期成分N1上发现了情绪效价主效应及其与返回抑制效应的交互作用。情绪信息能吸引个体的注意,具有情绪意义的刺激会占用更多的认知资源,从而引起注意偏向(Lang,1995)。在内隐情绪加工中,不需要对情绪图片进行效价判断,所以情绪效价对个体的影响不会受到意识的影响。在我们的研究中发现,N1中情绪效价主效应显著,情绪性图片比中性图片波幅更大,说明情绪信息在早期的知觉阶段就能吸引个体注意,在前注意阶段个体就受到了情绪信息的影响。也有研究发现,同样要求被试完成内隐情绪任务时,在与注意相关的P2成分上,负性刺激比正性刺激的ERP更大(Carretié, Mercado, Tapia, &Hinojosa, 2001)。由此可以认为,对情绪信息的注意偏向可能发生在早期的信息加工过程。关于返回抑制是否受情绪刺激的影响一直是备受关注的问题。在N1上,我们发现情绪效价和返回抑制之间交互作用显著,情绪图片在有效线索上的波幅显著大于中性图片波幅,在无效线索上则相反,说明返回抑制受到情绪效价的调节。Taylor和Therrien(2008)采用非面孔、拼凑面孔和完整的中性面孔,发现不同特性的靶子在返回抑制中产生了差异,说明返回抑制受到了生物性刺激的影响,此时IOR不再是一种“盲目机制”。王敬欣,贾丽萍,白学军等(2013)发现,在定位任务中,中性面孔产生的返回抑制量显著大于情绪性面孔。显然情绪面孔引发个体的注意偏向,这种知觉阶段的注意对返回抑制产生了影响,即目标的情绪性对IOR起了调节作用。返回抑制作为人类进化过程中的一种适应机制,产生在知觉层面,所以和内隐情绪加工产生了冲突,导致返回抑制受到目标情绪性的调节。
综上,研究发现线索有效性在早期ERP成分上发生了变化,所以我们认为返回抑制的产生来源于早期的知觉抑制,是对先前注意过的位置打上抑制标签,结果支持知觉抑制理论。对情绪的内隐加工和返回抑制发生在同一信息加工阶段,有效线索上情绪性目标更能吸引注意,而无效线索上中性目标更能吸引注意。
5 结论
返回抑制效应是一种基于知觉变化的机制;返回抑制会受到生物意义刺激的影响,内隐情绪加工对返回抑制效应具有调节作用。
戴琴, 冯正直.(2009). 抑郁个体对情绪面孔的返回抑制能力不足. 心理学报, 41(12), 1175–1188.
邓晓红, 张德玄, 黄诗雪, 袁雯, 周晓林.(2010). 阈上和阈下不同情绪线索对返回抑制的影响. 心理学报, 42(3), 325–333.
龚栩, 黄宇霞, 王妍, 罗跃嘉.(2011). 中国面孔表情图片系统的修订. 中国心理卫生杂志, 25(1), 40–46.
黄宇霞, 罗跃嘉.(2009). 负性情绪刺激是否总是优先得到加工: ERP研究. 心理学报, 41(9), 822–831.
罗琬华, 曾敏, 李凌.(2003). 关于返回抑制的一项ERP研究. 心理科学,26(3), 562–563.
梦向丽, 王萌华, 王玉平.(2006). 面孔感知过程中的视觉完形负波. 中国康复理论与实践, 12(3), 185–187.
彭晓哲, 周晓林.(2005). 情绪信息与注意偏向. 心理科学进展, 13(4),488–496.
王敬欣, 贾丽萍, 白学军, 罗跃嘉.(2013). 返回抑制过程中情绪面孔加工优先: ERPs研究. 心理学报, 45(1), 1–10.
王敬欣, 贾丽萍, 张阔, 张赛.(2013). 不同位置的情绪面孔加工对返回抑制的影响. 心理科学, 36(2), 335–339.
王丽丽, 郭亚桥, 邱江, 张庆林.(2010). 返回抑制的认知神经机制: 来自ERP研究的证据. 心理科学, 33(6), 1422–1425.
王丽丽, 罗跃嘉, 郭亚桥, 张庆林.(2010). 面孔方位对返回抑制的影响. 心理科学, 33(1), 100–103.
张阳, 彭春花, 孙洋, 张明.(2013). 视觉返回抑制的认知机制. 心理科学进展, 21(11), 1913–1926.
Adam, J., O’Donnell, C., & Pratt, J.(2005). Response selection influences inhibition of return. European Journal of Cognitive Psychology, 17(3),319–328.
Bernat, E., Bunce, S., & Shevrin, H.(2001). Event-related brain potentials differentiate positive and negative mood adjectives during both supraliminal and subliminal visual processing. International Journal of Psychophysiology, 42(1), 11–34.
Carretié, L., Mercado, F., Tapia, M., & Hinojosa, J. A.(2001). Emotion,attention, and the 'negativity bias', studied through event-related potentials. International Journal of Psychophysiology, 41(1), 75–85.
Fox, E., Russo, R., Bowles, R., & Dutton, K.(2001). Do threatening stimuli draw or hold visual attention in subclinical anxiety?. Journal of Experimental Psychology: General, 130(4), 681–700.
Hopfinger, J. B., & Mangun, G. R.(2001). Tracking the influence of reflexive attention on sensory and cognitive processing. Cognitive,Affective, & Behavioral Neuroscience, 1(1), 56–65.
Hopfinger, J. B., & West, V. M.(2006). Interactions between endogenous and exogenous attention on cortical visual processing. NeuroImage,31(2), 774–789.
Ivanoff, J., & Klein, R. M.(2001). The presence of a nonresponding effector increases inhibition of return. Psychonomic Bulletin& Review, 8(2),307–314.
Ivanoff, J., & Klein, R. M.(2004). Stimulus-response probability and inhibition of return. Psychonomic Bulletin & Review, 11(3), 542–550.
Ivanoff, J., & Taylor, T. L.(2006). Inhibition of return promotes stop-signal inhibition by delaying responses. Visual Cognition, 13(4), 503–512.
Klein, R. M.(1988). Inhibitory tagging system facilitates visual search.Nature, 334(6181), 430–431.
Klein, R. M., & Dick, B.(2002). Temporal dynamics of reflexive attention shifts: A dual-stream rapid serial visual presentation exploration.Psychological Science, 13(2), 176–179.
Klein, R. M., & MacInnes, W. J.(1999). Inhibition of return is a foraging facilitator in visual search. Psychological Science, 10(4), 346–352.
Krüger, H. M., MacInnes, W. J., & Hunt, A. R.(2014). Perceptual merging contributes to cueing effects. Journal of Vision, 14(7), doi:10.1167/14.7.13.
Lang, P. J.(1995). The emotion probe: Studies of motivation and attention.American Psychologist, 50(5), 372–385.
Mangun, G. R., & Hillyard, S. A.(1991). Modulations of sensory-evoked brain potentials indicate changes in perceptual processing during visual-spatial priming. Journal of Experimental Psychology: Human Perception and Performance, 17(4), 1057–1074.
McDonald, J. J., Ward, L. M., & Kiehl, K. A.(1999). An event-related brain potential study of inhibition of return. Perception & Psychophysics,61(7), 1411–1423.
Pan, F. D., Wu, X. G., Zhang, L., & Ou, Y. H.(2017). Inhibition of return is modulated by negative stimuli: Evidence from subliminal perception.Frontiers in Psychology, 8, 1012.
Pérez-Dueñas, C., Acosta, A., & Lupiáñez, J.(2014). Reduced habituation to angry faces: Increased attentional capture as to override inhibition of return. Psychological Research, 78(2), 196–208.
Poliakoff, E., Latif, A., Maehr, A. M., Marshall, H., Roesser, A., Scurr, T.,…, Lander, K.(2016). An unsuccessful attempt to demonstrate attentional orienting within the purely emotional domain. Emotion,16(1), 6–10.
Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. In H.Bouma & D. Bouwhuis (Eds.), Attention and performance X: Control of language process (pp. 531–556). Hillsdale, NJ: Erbaum.
Prime, D. J., & Jolicæur, P.(2009). On the relationship between occipital cortex activity and inhibition of return. Psychophysiology, 46(6),1278–1287.
Prime, D. J., & Ward, L. M.(2004). Inhibition of return from stimulus to response. Psychological Science, 15(4), 272–276.
Prime, D. J., & Ward, L. M.(2006). Cortical expressions of inhibition of return. Brain Research, 1072(1), 161–174.
Sapir, A., Jackson, K., Butler, J., Paul, M. A., & Abrams, R. A.(2014).Inhibition of return affects contrast sensitivity. The Quarterly Journal of Experimental Psychology, 67(7), 1305–1316.
Sapir, A., Soroker, N., Berger, A., & Henik, A.(1999). Inhibition of return in spatial attention: Direct evidence for collicular generation. Nature Neuroscience, 2(12), 1053–1054.
Silvert, L., & Funes, M. J.(2016). When do fearful faces override inhibition of return?. Acta Psychologica, 163, 124–134.
Stoyanova, R. S., Pratt, J., & Anderson, A. K.(2007). Inhibition of return to social signals of fear. Emotion, 7(1), 49–56.
Taylor, T. L., & Klein, R. M.(1998). On the causes and effects of inhibition of return. Psychonomic Bulletin & Review, 5(4), 625–643.
Taylor, T. L., & Therrien, M. E.(2005). Inhibition of return for faces.Perception & Psychophysics, 67(8), 1414–1422.
Taylor, T. L., & Therrien, M. E.(2008). Inhibition of return for the discrimination of faces. Perception & Psychophysics, 70(2), 279–290.
van Honk, J., Tuiten, A., de Haan, E., vann de Hout, M., & Stam, H.(2001).Attentional biases for angry faces: Relationships to trait anger and anxiety. Cognition and Emotion, 15(3), 279–297.
Vogel, E. K., & Luck, S. J.(2000). The visual N1 component as an index of a discrimination process. Psychophysiology, 37(2), 190–203.
Modulation of Implicit Emotion Process on Inhibition of Return Effect
WU Xiaogang1,2, SHI Liang1, LI Leyuan1, PAN Fada1
(1 School of Education Science, Nantong University, Nantong 226019; 2 Department of Psychology, Research Center for Psychological and Behavioral Sciences, Soochow University, Suzhou 215123)
Inhibition of return is an inhibitory mechanism which refers to the position previously noticed showing slower response.Providing neural, positive and negative facial images as target stimulus, subjects were asked to perform detection task with ERPs technology. It was found that the data on the behavior and EEG record had significant IOR effect. According to EEG results, no effects on the component of P1 were found. However, the effect of IOR were found on N1 and N170. Compared with amplitude of the invalid cues, we found that the amplitude of N1 of the valid cues was larger, while the amplitude of N170 was smaller.Interestingly, there are emotion effect of N1, indicating that emotional face induced more attentional bias than neural face. More surprisingly, we found a significant interaction effect between emotion values and IOR. The simple effect showed that the amplitude of emotional face was larger than neural face. These results indicate that IOR at least partly arises from changes in perceptual process, and to a certain degree, implicit emotion process can modulate IOR.
inhibition of return, implicit emotion processing, Event Related Potentials.
B842
2016–12–02
教育部人文社会科学研究基金项目(12YJC190026),江苏省社会科学基金项目(14SHC006),江苏省教育科学“十二五”规划项目(C-a/2013/01/003)和江苏省2015年度普通高校研究生科研创新项目(KYZZ15_0355)。
潘发达。E-mail:panfada1979@126.com。
注:1正性面孔选用PF1、PF42、PM95和PM99,负性面孔选用NF60、NF64、NM68和NM69,中性面孔选用TF3、TF18、TF66、TF88、TM56、TM74、TM76和TM100。
