基于不同尺度和模型的小秦岭谱系结构研究
2017-10-25毕帅帅王雪颖韩军旺贾宏汝叶永忠袁志良
牛 帅, 毕帅帅, 王雪颖, 韩军旺, 贾宏汝, 叶永忠, 袁志良
(1.河南农业大学生命科学学院,河南 郑州 450002; 2.河南小秦岭国家级自然保护区管理局,河南 灵宝 472500; 3.河南财政金融学院,河南 郑州 451464)
基于不同尺度和模型的小秦岭谱系结构研究
牛 帅1, 毕帅帅1, 王雪颖1, 韩军旺2, 贾宏汝3, 叶永忠1, 袁志良1
(1.河南农业大学生命科学学院,河南 郑州 450002; 2.河南小秦岭国家级自然保护区管理局,河南 灵宝 472500; 3.河南财政金融学院,河南 郑州 451464)
为探索研究尺度、零模型与群落谱系结构的关系,在小秦岭自然保护区建立1 hm2样地,运用约束型、非约束型零模型分别从10、20、25 m 3个研究尺度上分析群落谱系结构,结果表明:(1)非约束型模型下,谱系结构聚集;约束型模型下,10、20 m尺度上,谱系聚集,25 m尺度上谱系分散。对比发现,随研究尺度增大,谱系聚集程度降低,趋于分散,可能是由于环境异质性对物种分布的影响程度降低,密度制约作用增强所致。(2)空间因子对谱系结构的影响远远大于地形因子,随研究尺度增加,空间因子及地形因子的解释量逐渐增大。地形的不同营造了不同的空间差异,二者密切相关。因此,空间因子是影响谱系结构形成的直接因素,而地形因子是间接因素。
谱系结构;地形因子;空间因子;净谱系亲缘关系指数;最近种间亲缘关系指数
“群落谱系结构”法是用来研究群落中物种组成和动态变化,追溯群落构建的历史,探究群落形成原因的一种手段,是近代生物生态学专家热衷使用的一种新方法[1]。2000年WEBB等[2]首个提出了谱系树(phylogenetics tree)的理念,并将其运用到群落生态学的研究中,为近代生物生态学的研究开辟了新方向。黄建雄等[3]进行了相关的谱系结构研究后提出,群落内现有物种组成是进化过程和生态过程共同作用的结果,而分析物种间亲缘关系可以反映现有群落形成的历史过程。谱系聚集、谱系发散以及谱系随机是谱系结构研究的普遍结果,经过大量的研究发现,植物群落的谱系结构往往呈现出聚集或发散的格局[4-5]。生态学家围绕植物群落的谱系结构的分布展开了深入研究,先后提出了几种理论来解释植物群落的共存机制[6-9]。WEBB等[10]和KRAFT等[11]提出的生态位理论、中性理论、密度制约假说成为目前最具权威的理论。生态位理论认为生境过滤是维持群落共存的主要原因,筛选出生态特征相似的物种共存,使亲缘关系较近的物种的系统发育结构表现为相对聚集;中性理论认为群落内的物种具有相同的竞争能力、出生率和死亡率,因此群落的物种往往呈随机分布;密度制约是维持群落物种共存的一个重要因素,密度制约同种或亲缘关系相近的物种之间相互排斥,因此近缘种之间的密度制约能在一定程度上解释群落发散的系统发育结构[12-16]。小秦岭国家级自然保护区位于河南省西部,属典型温带落叶阔叶林,优越的物种资源使该区域成为了研究物种共存机制的优质样地[17]。本研究以该区1 hm2落叶阔叶林森林动态样地的木本植物数据为基础,研究了不同尺度上的群落谱系结构,分析在不同零模型和不同研究尺度下,样地的谱系结构产生的变化和影响温带落叶阔叶林物种生物多样性的生存机制。基于第一个问题的研究结果,本研究又比较了空间因子和地形因子对小秦岭落叶阔叶谱系结构的影响,探讨影响温带落叶阔叶林的非生物因素,为温带落叶阔叶林的保护和管理提供科学依据。
1 材料与方法
1.1 研究区概况
小秦岭国家级自然保护区位于秦晋豫3省交界处,地处河南省灵宝市西部,地理坐标为34°23′~34°31′N,110°23′~110°44′E[15-16],东西长31 km,南北宽12 km,全区面积15 160 hm2。森林覆盖率81.2%,其主峰老鸦岔垴海拔2 413.8 m,为河南省最高峰。本区位于暖温带大陆性气候区,属季风型半干旱气候,年均气温11.2~14.2 ℃,极端最低温度-17 ℃,极端最高温度42.7 ℃,无霜期170~215 d,年降水量620 mm。保护区内土壤类型从南到北分布格局明显,依次为山地棕壤、山地黄棕壤和山地褐土3种[18-20]。小秦岭自然保护区植被类型丰富,主要由针叶林、阔叶林、针阔叶混交林、竹林、灌丛、灌草丛和草甸组成。该区域横跨东西,地处暖温带向亚热带的过渡区,植被带谱有些不同,南坡带有亚热带植被特色,北坡则具华北暖温带植物区系特征。
1.2 样地设置
2012年,在保护区中选取远离生活区和路径,群落保护较好的区域,参照BCI样地的技术规范,建立了一个100 m×100 m的样地(图1)。用全站仪将样地分成25个20 m×20 m的子样方,再将每个20 m×20 m的子样方划分为16个5 m×5 m的小样方,对每个样方中胸径(diameter at breast height,DBH)≥1 cm的个体进行每木调查,记录其种类、胸径、坐标等,定位并挂牌(表1)。使用GPS测量每个样方的海拔,地质罗盘测量坡度和坡向。所选样地海拔合适,生物多样性丰富,生境异质性强,地形复杂,包含了山坡、山顶、山谷、水流等多种生境,对于调查小秦岭自然保护区的植被特征具有重要意义。
1.3 数据分析
1.3.1 谱系树的构建及指数的选择 基于ZANNE等[21]系统树作图方法,收集小秦岭100 m×100 m样地物种有关信息,建立数据库,将数据库中物种名录及科属等信息输入Phylomatic软件,以被认可的被子植物分类系统APG Ш为基础数据,识别输入的物种名录信息,在线直接输出系统发育进化树。最后应用Phylocom软件校正样地各被子植物物种的谱系系统发育长度,输出谱系树[10],见图2所示。图2中,末端节点表示现存物种;种间节点表示分化事件,或其后代最近的共同祖先;枝是连接相邻2个节点的线,同一祖先衍生出来的物种由各分枝相连。
为直观表达小秦岭温带落叶阔叶林群落谱系结构,选择净谱系亲缘关系指数(net relatedness index,NRI)和最近种间亲缘关系指数(net nearest taxa index,NTI)来表示。根据各个物种在系统进化树上的位置,首先利用R语言中的picante程序包计算最近谱系距离(mean nearest taxon distance,MNTD)及平均谱系距离(mean phylogenetic distance,MPD),然后根据实际观察值和相关零模型的模拟值的差异,利用picante软件包计算出谱系指数NRI和NTI的数值,判断响应研究尺度下群落谱系结构的分布状态。计算公式如下:
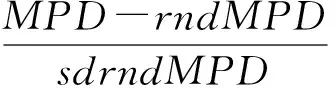
(1)

(2)
(3)
(4)
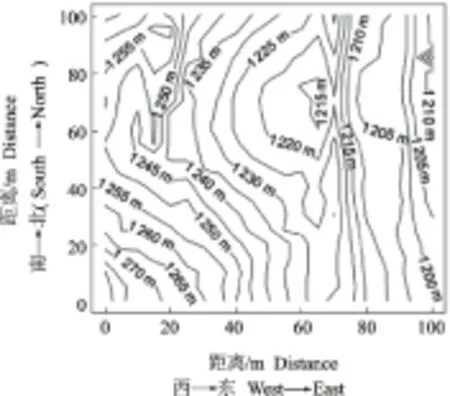
图1 样地等高线Fig.1 Contour map of the sample plot

表1 小秦岭自然保护区样方内物种Table 1 The species list in Xiaoqinling nature reserve sample plot

图2 小秦岭保护区域样方内有支长的植物系统进化树Fig.2 Phylogenetic tree with branch lengths for the sample plants in the dry-hot valley of the Xiaoqinling
1.3.2 分析方法 基于野外样方调查数据,将样地划分为10 m×10 m、20 m×20 m、25 m×25 m 3个尺度来研究群落系统发育结构与尺度的关系。分别采用KEMBEL等[20]提出的2种零假设模型,即约束型和非约束型,计算出指数NRI和NTI,用以直观地表示各研究尺度上群落物种的谱系结构分布状态,根据群落物种的系统发育结构随研究尺度的动态变化情况,探讨影响群落物种组成的原因。运用Wilcoxon rank-sum非参数检验方法,将观察值和零模型下模拟值之差的绝对值之和作为其检验统计量,比较其成对分布是否对称,分析不同尺度上NRI和NTI的差异是否有统计学意义。
探讨非生物因素(地形因子和空间因子)对小秦岭落叶阔叶林谱系结构影响时,首先将环境因子作为自变量,使用前面所选的简化模型,筛选出决定系数R2最大,即对谱系结构有显著影响的4个因素(平均海拔、凹凸度、坡向和坡度)作为地形因子变量,然后通过主轴邻距法(principal coordinates of neighbor matrices,PCNM),在3个尺度(10 m×10 m、20 m×20 m、25 m×25 m)上分别产生69、18和11个PCNM变量,挑选出显著的PCNM变量作为空间因子变量用于谱系结构分析。最后,用vegan程序包对各尺度的样方中植物群落的谱系结构进行方差分解。方差分解的结果分为4部分:(1)纯地形因子解释的部分;(2)纯空间因子解释的部分;(3)地形因子和空间因子共同解释的部分。
2 结果与分析
2.1 群落的谱系结构
群落谱系结构见图3。从图3可看出,基于不同尺度和不同零模型的小秦岭森林净相关指数(NRI)和平均最近邻体指数(NTI)的数值分析,不同尺度和不同零模型的谱系结构都存在NRI和NTI小于0或大于0的样方,非约束性模型所示,NRI和NTI数值大于0的样方数量大于数值小于0的样方数量,谱系结构聚集的样方数比分散的样方数多,随着研究尺度的增大,整个样方的排布逐渐趋于以0为轴的正态分布,说明谱系结构的聚集程度减弱。通过比较不同尺度下的约束性模型可以看出,NRI和NTI数值的分布较为均匀,但随着尺度的增大,NRI和NTI数值大于0的样方逐渐减少,说明谱系结构的聚集程度减弱,25 m× 25 m开始表现为分散。
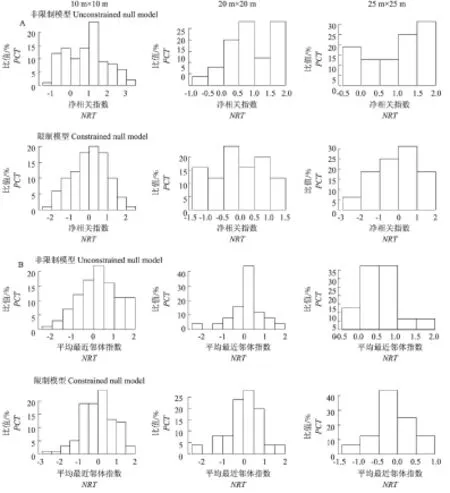
A代表净相关指数(NRI),B代表平均最近邻体指数(NTI)。
The letter A represents net relatedness index (NRI) and the letter B represents nearest taxon index (NTI).
图3基于不同尺度和不同零模型的小秦岭森林净相关指数(NRI)和平均最近邻体指数(NTI)
Fig.3TreecommunityphylogeneticstructureinquadratsatthreespatialscaleswithintheforestdynamicsplotinXiaoqinlingnaturereserve
从表2和表3可以看出,不同零模型的采用对群落谱系结构的影响差异非常显著,不同研究尺度的分析对群落谱系结构差异也十分明显。具体分析,采用非约束性模型比较,随着尺度的增大,净相关指数NRI和最近邻体指数NTI逐渐减小,NRI和NTI的数值在3个不同的研究尺度上都大于0,谱系结构表现为聚集,而且随着研究尺度的增大NRI和NTI的数值减小,说明谱系结构聚集程度减弱。采用约束性模型比较,谱系结构在10 m×10 m、20 m×20 m的研究尺度上,显示为聚集减弱的表现,在25 m×25 m的研究尺度上,NRI和NTI的数值小于0,谱系结构表现为分散。
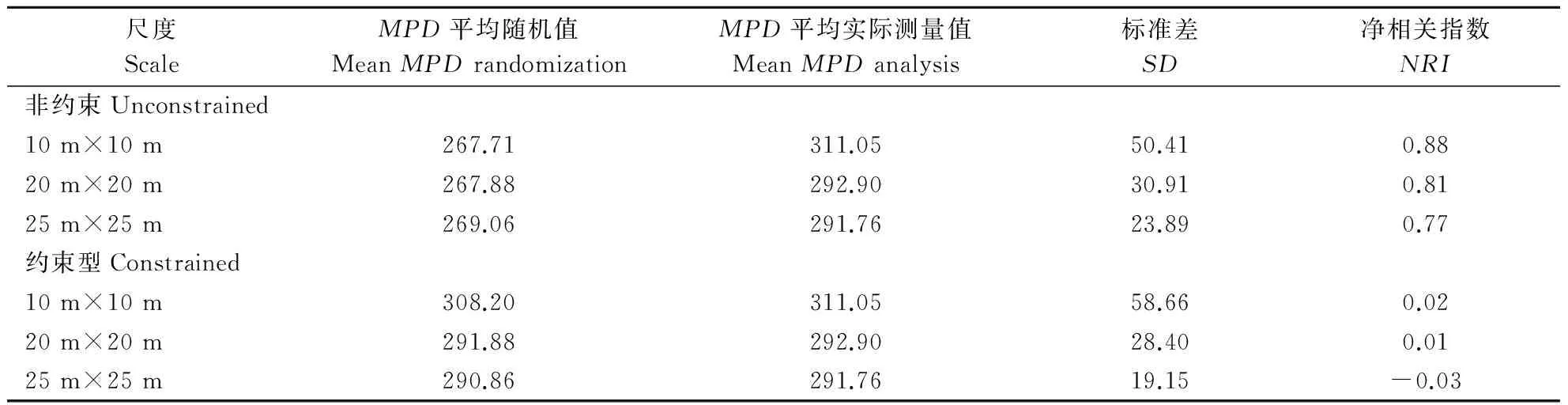
表2 不同零模型和不同尺度下小秦岭森林平均成对分类系统发育距离(MPD)和净相关指数(NRI)Table 2 Mean pairwise phylogenetic distance (MPD) and net relatedness index (NRI) with different null models and different spatial scales in a 1 hm2 deciduous broad-leaved forest plot in Xiaoqinling nature reserve
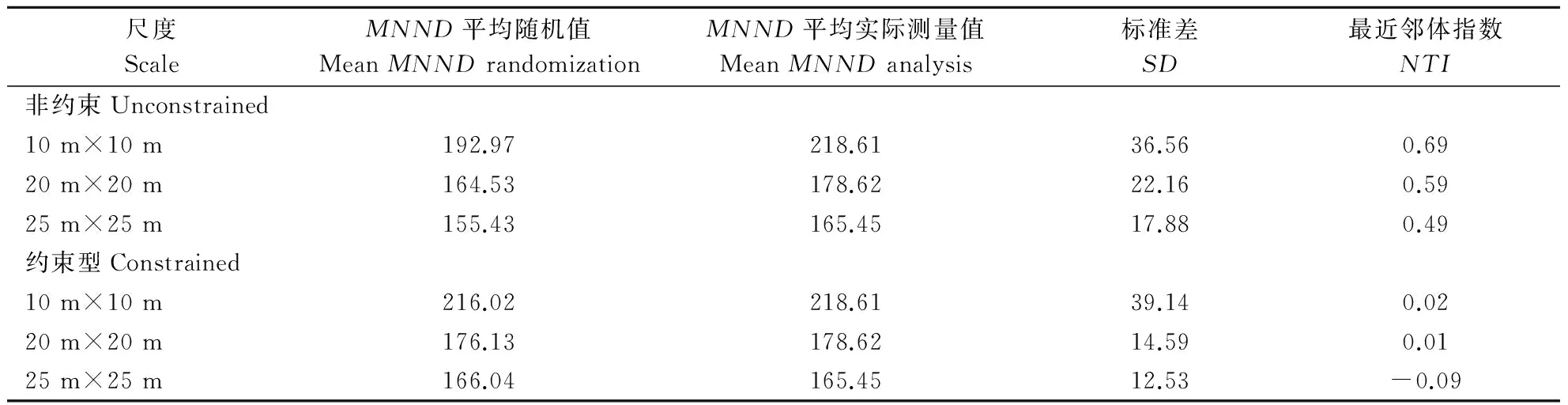
表3 不同零模型和不同尺度下小秦岭森林平均最近系统发育距离(MNTD)和最近邻体指数(NTI)Table 3 Mean nearest taxon distance (MNTD) and nearest taxon index (NTI) with different null models and different spatial scales in a 1 hm2 deciduous broad-leaved forest plot in Xiaoqinling nature reserve
2.2 空间因子和地形因子对群落谱系结构的影响
空间因子和地形因子对样地木本植物群落谱系结构的影响见表4。使用方差分解分析空间变量和地形因子对谱系指数的影响,残差结果均大于0.1,因此该2种因子对谱系结构影响不显著。地形因子单独影响,地形因子和空间因子共同影响对谱系结构的解释量均小于10%,相比之下,空间因子对于谱系结构产生的解释量较大。随着研究尺度的增加,纯地形因子以及2种因子共同对谱系结构的影响逐渐增大。因此,地形因子不是影响研究区域谱系结构分布的主要因素。空间因子对谱系结构的解释量较大,且地形因子随研究尺度的增加而增大,说明该区域谱系分布的区域性明显。
3 讨论
KEMBEL等[22-23]研究发现,群落的系统发育结构与研究尺度之间有一定的相关性,即随着群落空间尺度的增大,谱系结构会表现出明显的变化。SWENSON等[24]进一步详细研究了空间尺度和谱系结构的关系,结果表明: 小空间尺度上, 谱系趋向于发散, 大的空间尺度上,谱系就开始趋向聚集。本研究中,采用非约束性模型比较,NRI和NTI的数值在3个不同的研究尺度上都大于0,谱系结构表现为聚集,由此推断生境过滤是小秦岭物种群落组成的重要原因。然而采用约束性模型比较时,两种群落谱系亲缘指数呈现不同的结果:在10 m×10 m、20 m×20 m的研究尺度上,NRI和NTI的数值大于0,且随尺度的增加数值减小,谱系结构表现为聚集,因此生境过滤是该尺度上植物群落构成的重要因素,但并不能完全解释小秦岭研究区域植物群落形成的问题;在25 m×25 m的研究尺度上,NRI和NTI的数值小于0,谱系结构表现为分散,这可能是由于谱系近的物种发生了竞争,即密度制约对该研究尺度下植物群落的分布产生了重要影响。

表4 空间因子和地形因子对样地木本植物群落谱系结构的影响Table 4 Variance portioning of the relative roles of topography heterogeneity and spatial factor on net relatedness index (NRI) and nearest taxon index (NTI)
KEMBEL等[22]研究了植物群落谱系相关亲缘指数NRI和NTI,发现运用约束性零模型和非约束性零模型两者结合比较,能更好地检测生境过滤构建的群落谱系结构。本研究中,采用非约束性模型比较,随着研究尺度的增大,小秦岭自然保护区的谱系结构呈现聚集度逐渐减小的聚集分布;采用约束性模型比较,随着研究尺度的增大,谱系结构呈现聚集程度逐渐减小,以致在大尺度25 m×25 m上呈分散分布。2种模型比较来看,随着尺度增大,小秦岭自然保护区的谱系结构趋向于分散分布,由此可见,在大尺度上生态位理论的生境过滤理论对该研究区域的物种群落分布影响较小,密度制约成为影响小秦岭落叶阔叶林植物群落构成的主导因素。
袁志良等[25]对暖温带-北亚热带过渡区宝天曼自然保护区1 hm2固定检测样地物种相关性分析中指出纯地形因子对物种分布的影响不显著。而在本研究中,地形因子对研究区域群落谱系的解释量很低,对谱系结构的形成不显著,但随着研究尺度的增加,地形因子的解释量却在增大,这可能是样地面积太小,地形结构变化不大,未能显示地形因子对谱系结构的影响。空间因子对于谱系结构产生的影响远远大于地形因子的影响,随着研究尺度的增加,空间因子的解释量以及地形因子的解释量都在增大。而空间因子和地形因子密切相关,地形的不同营造了不同的空间差异,因此,空间因子是影响谱系结构形成的直接因素,而地形因子是间接因素。
研究某一区域植物群落的谱系结构不仅能推测出该地区群落结构形成的历史原因,还可能通过现状推测以后的发展方向。本研究探讨了研究尺度的变化对谱系结构所产生的影响,推测出:不同研究尺度下,影响植物群落结构形成的生存机制也不同。但是通过现状推测植物群落以后的发展方向并证实仍需长时间的野外动态调查,建立大数据数据库进行综合分析。在探索非生物因素(地形因子和空间因子)对物种分布的影响时,地形因子和空间因子经过相关性分析,对物种分布的影响不显著,但随着研究尺度的增大,地形因子的解释量在增大,如果研究尺度足够大,地形因子的影响将会呈现什么样的状况,仍需进一步研究。
[1] 牛红玉,王峥峰,练琚愉,等. 群落构建研究的新进展:进化和生态相结合的群落谱系结构研究[J]. 生物多样性, 2011, 19(3):275-283.
[2] WEBB C O, ACKERLY D D, MCPEEK M A, et al. Phylogenies and community ecology[J]. Annual Review of Ecology and Systematics, 2003, 33(1):475.
[3] 黄建雄,郑凤英,米湘成. 不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响[J]. 植物生态学报, 2010, 34(3):309-315.
[4] VAMOSI S M, HEARD S B, VAMOSI J C, et al. Emerging patterns in the comparative analysis of phylogenetic community structure[J]. Molecular Ecology, 2009, 18(4):572-92.
[5] GROFFMAN P M, BAIN D J, BAND L E, et al. Down by the riverside: urban riparian ecology[J]. Frontiers in Ecology & the Environment, 2003, 1(6):315-321.
[6] GRUBB P J. The maintenance of species-richness in plant communities: the importance of the regeneration niche[J]. Biological Reviews, 1977, 52(1):107-145.
[7] TILMAN D. Resource competition and community structure[M]. Princeton: Princeton University Press, 1982:296.
[8] TAYLOR D R, AARSSEN L W, LOEHLE C. On the relationship between r/k selection and environmental carrying capacity: a new habitat templet for plant life history strategies[J]. Oikos, 1990:58(2):239-250.
[9] HUBBELL S P, AHUMADA J A, CONDIT R, et al. Local neighborhood effects on long-term survival of individual trees in a neotropical forest[J]. Ecological Research, 2001, 16(5): 859-875.
[10] WEBB C O, ACKERLY D D, KEMBEL S W. Phylocom: software for the analysis of phylogenetic community structure and trait evolution[J]. Bioinformatics, 2008, 24(18):2098-2100.
[11] KRAFT N J B, COMWELL W K, WEBB C O, et al. Trait evolution, community assembly, and the phylogenetic structure of ecological communities[J]. The American Naturalist, 2007, 170(2):271-283.
[12] 李新辉,刘延虹,刘晔,等. 地理距离及环境差异对云南元江干热河谷植物群落beta多样性的影响[J]. 生物多样性, 2016, 24(4):399-406.
[13] 宋凯,米湘成,贾琪,等. 不同程度人为干扰对古田山森林群落谱系结构的影响[J]. 生物多样性, 2011, 19(2):190-196.
[14] 卢孟孟,黄小翠,慈秀芹,等. 沿海拔梯度变化的哀牢山亚热带森林群落系统发育结构[J]. 生物多样性, 2014, 22(4):438-448.
[15] 韩军旺,冯佳伟,张旭, 等. 河南小秦岭自然保护区木本植物优势种群的生态位特征[J]. 河南农业大学学报, 2016, 50(1):85-91.
[16] 任思远,王婷,祝燕,等. 暖温带-北亚热带过渡带落叶阔叶林群落不同径级系统发育结构的变化[J]. 生物多样性, 2014, 22(5):574-582.
[17] 贾宏汝,张旭,陈云,等. 小秦岭国家级自然保护区乔木生物量随地形因子变化格局[J]. 河南农业大学学报, 2015, 49(6):777-782.
[18] 韦博良,陈云,许宁,等. 小秦岭国家级自然保护区维管植物群落结构与组成[J]. 河南农业大学学报,2015, 49(3):335-342.
[19] 陈云,王海亮,韩军旺,等. 小秦岭森林群落数量分类、排序及多样性垂直格局[J]. 生态学报, 2014, 34(8):2068-2075.
[20] 王海亮. 河南小秦岭自然保护区天然混交林物种多样性研究[J]. 河南科学, 2013, 31(1):53-57.
[21] ZANNE A E, TANK D C, CORNWELL W K, et al. Three keys to the radiation of angiosperms into freezing environments[J]. Nature, 2014, 506(7486): 89-92.
[22] KEMBEL S W, HUBBELL S P. The phylogenetic structure of a neotropical forest tree community[J]. Ecology, 2006, 87(sp7):86-99.
[23] KEMBEL S W. Disentangling niche and neutral influences on community assembly: assessing the performance of community phylogenetic structure tests[J]. Ecology Letters, 2009, 12(9):949-60.
[24] SWENSON N G, ENQUIST B J, THOMPSON J, et al. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities[J]. Ecology, 2007, 88(7): 1770-1780.
[25] 袁志良,陈云,韦博良,等. 暖温带-北亚热带生态过渡区物种生境相关性分析[J]. 生态学报, 2013, 33(24):7819-7826.
StudyonphylogeneticstructureinXiaoqinlingbasedondifferentscalesandmodels
NUI Shuai1, BI Shuaishuai1, WANG Xueying1, HAN Junwang2, JIA Hongru3, YE Yongzhong1, YUAN Zhiliang1
(1.Collage of Life Science, Henan Agricultural University, Zhengzhou 450002, China;2.Administration Bureau of Xiaoqinling National Nature Reserve of Henan, Lingbao 472500,China; 3.Henan Finance and Taxation College,Zhengzhou 451464,China)
We established 1 hm2sample plot in Xiaoqinling nature reserve, used two statistical models (the constraint model and the non-constraint model) and analyzed the community phylogenetic structure in this region from three different scales 10, 20, 25 m. The results show: (1) under the non-constraint model, communities were phylogenetically clustered at all spatial scales; Using the constraint model, communities were phylogenetically clustered in 10 m, 20 m scale and overdispersed in 25 m research scale. In contrast, community phylogenetic structures tend to disperse with the scale enlargement. Therefore, phylogenetic density dependence becomes the leading factor of the community compositionin the Xiaoqinling deciduous broad-leaved forest. (2) The study results suggest that the impact of space factors on community phylogenetic structure is much greater than that of the terrain factor. With the increasing of the reseach scale, space factors and terrain factors influence gradually increased. Space factors and terrain factors are closely related to different terrain caused by the space difference. Therefore, The space factors are the direct factors affecting the community phylogenetic structure, and terrain factors are the indirect factors.
phylogenetic structure; terrain factor; space factor; net relatedness index; nearest taxa index
2016-12-12
河南省科技攻关项目(132102110133);河南省教育厅科学技术研究重点项目(14A180013)
牛 帅(1988-),女,河南周口人,硕士研究生,主要从事森林生态方面的研究。
袁志良(1976-),男,河南信阳人,副教授,博士。
1000-2340(2017)04-0495-08
S718
A
(责任编辑:朱秀英)
