珍稀植物红椿种群生命表与生存分析
2017-10-21张汉菊程德华徐晓婷
汪 洋,黄 芳,张汉菊,程德华,徐晓婷
(1.湖北生态工程职业技术学院,湖北 武汉 430200;2.湘潭第一中学,湖南 湘潭 411100)
珍稀植物红椿种群生命表与生存分析
汪 洋1,黄 芳1,张汉菊1,程德华2,徐晓婷1
(1.湖北生态工程职业技术学院,湖北 武汉 430200;2.湘潭第一中学,湖南 湘潭 411100)
通过对恩施盛家坝红椿群落的样地调查和数据统计,编制红椿种群生命表进行生存分析,探究种群数量动态变化。结果表明:红椿种群动态指数Vpi=27.84%>0,种群属于增长型;V’pi=1.74%>0,种群仍稳定,抗风险能力弱。种群生命表和生存曲线显示数量动态变化趋势符合Deevey-Ⅲ型。死亡率出现2个高峰,1~2龄级为0.250;第3~4龄级为0.364;龄级间最大损失度在第3龄级,为0.452。生存率函数最大值在1龄级,为0.75,呈单调下降趋势;累计死亡率在第8龄级,最大值为1,呈单调增加趋势。死亡密度函数f(t)波动出现3个高峰,在第1和第3龄级,分别为0.063和0.063;在6和7两个龄级均为0,第8龄级上升到最高,为0.078;危险率函数λ(t)在第3龄级时最高,达到0.111;到6和7两个龄级为0,第8级达到峰值0.5。在幼苗和中树阶段受限于环境筛和干扰损失较大,也是其濒危主要因素。红椿种群成树过渡阶段稳定,逐渐进入新一轮竞争。
红椿;种群;生命表;生存分析
红椿(ToonaciliataRoem.),别名红楝子,楝科香椿属,半常绿或落叶乔木,国家Ⅱ级重点保护野生植物,是我国热带、亚热带地区的珍贵速生用材树种。其木材为上等家具用材,素有“中国桃花心木”之称[1,2]。天然状况下,红椿主要以散生为主, 较大成片群落很少,且天然更新能力较差[3]。目前对红椿种质资源、遗传结构、植物生理、育种造林、化学及药理等方面进行了较为广泛的研究[4]。有关红椿天然林和人工林的种群生态学研究涉及较少。
种群动态是一定时间和空间范围内种群的大小和数量变化规律[5,6]。种群年龄结构反映了种群在特定时间世代重叠的特征,是分析探索种群动态的有效方法,被广泛应用于种群动态研究中[6,7]。生命表和存活曲线对于研究种群数量的统计及其动态特征是必要工具。生命表所反映的重要参数有助于分析红椿种群生命过程及动态特征,对种群生态学研究和生态保护有着理论与现实意义。
1 研究地概况
研究地位于恩施市境内星斗山保护区边沿的盛家坝乡,马鹿河流域。地处中亚热带与北亚热带的过渡带,属亚热带大陆性季风气候。气候变化较大,山地气候明显。由于区内各地海拔差异,高低山之间气候明显不同。区内地形复杂,制约着光、热、水的再分配。该区年均温14.9 ℃,年降雨量达1 300 mm以上[8, 9]。5—9月间的降水量占全年的71%,年内季节分配不均匀。土壤为质地疏松的山地黄壤[8],样地土壤pH为6.5。土壤自然含水率较高,有机质及氮、磷和钾含量相对较高。
马鹿河样地存在一定的人为干扰,为常绿落叶阔叶天然次生林。林地乔灌草丰富度较高,结构完整,藤本植物层及其丰富,且大型藤本已进入乔木层的上层。除红椿外,主要乔木包含:鳞木稠李(Padusbuergeriana)、竹叶楠(Arundinagraminifolia)、黑壳楠(Linderamegaphylla)等。灌木层主要有棠叶悬钩子(Rubusmalifolius)、臭牡丹(Clerodendrumbungei)、砚壳花椒(Zanthoxylumdissitum)等。主要草本有团叶鳞始蕨(Lindsaeaorbiculata)、透茎冷水花(Pileapumila)、楼梯草(Elatostemainvolucratum)等。
2 调查研究方法
2.1 野外调查方法
马鹿河流域的红椿种群较为完整。设立样地完全覆盖全部存活植株,面积为20 m×20 m。在每块标准样地内采用相邻格子法(格子大小为5 m×5 m),共设置16个样方。对胸径≤2.5 cm的植株,测定地径、高度;对大于胸径≥2.5 cm的活立木,记录胸径、冠幅、枝下高等指标。同时记录环境因子。

表1 马鹿河红椿群落环境特征
2.2 龄级划分方法及动态分析
红椿是国家二级保护植物,不可能截取圆盘进行解析植株胸径与年龄来进行模型拟合。红椿年龄与胸径之间相关性较高[10,11]。通过红椿平均胸径与连年生长过程分析,结合样地内植株年龄最大为37年(生长锥测得),胸径36 cm,采用“空间替代时间法”,即将林木依胸径大小分级,以立木级结构代替种群年龄结构分析种群动态[12,13,14]。根据该种群测量径级,将该种群划分为8个龄级。即幼苗级Ⅰ(DBH<2.5 cm);从第2龄级开始,以胸径5 cm为步长增加一级,即幼树级Ⅱ(2.5 cm≤DBH<7.5 cm),Ⅲ(7.5 cm≤DBH<12.5 cm);Ⅳ(12.5 cm ≤DBH<17.5 cm);中树级Ⅴ(17.5 cm≤DBH<22.5 cm),Ⅵ(22.5 cm≤DBH<27.5 cm);大树级Ⅶ(27.5 cm≤DBH<32.5 cm),Ⅷ(DBH≥32.5 cm)[15]。图1表示不同龄级存活株数所占全部植株的百分比。
种群年龄结构的动态指数采用陈晓德的数学模型进行推导,从而对红椿种群动态进行定量描述[16, 17],Vn,Vpi,Vpi′取正、负、零值的意义分别反映种群或相邻年龄级个体数量的增长、衰退、稳定的动态关系。
2.3 编制生命表
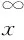
2.4 生存函数
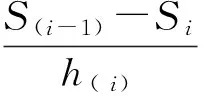
根据上述4个生存函数的估算值,绘制红椿种群的生存曲线、累计死亡率曲线、死亡密度曲线和危险率曲线[13]。
3 结果与分析
3.1 龄级结构与动态
根据龄级划分结果,以年龄级为横坐标,以各龄级存活红椿数量与种群全部存活数量百分比数为纵坐标,绘制红椿种群龄级结构分布图。
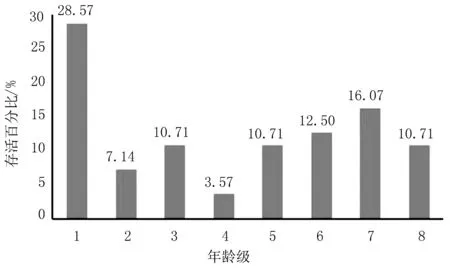
图1 红椿种群龄级与存活百分比分布Fig.1 Age groups and survival percentage of Toona ciliata population
由图1可知,红椿幼苗在1龄级阶段占比达28.57%,第7龄级占16.07%,最小占比反应在第4龄级上,只有3.57%。种群龄级分布不规则,出现2次波谷,分别在第2、第4龄级上。3、5、6级表现稳定。
红椿种群Vn分析表明,第2~8龄级个体数量变化动态指数分别为:75.00%, -33.33%,66.67%,-66.67%,-14.27%,-22.22%,33.33%。对照图1,该数据表明第2~3级,4~7龄级动态表现为负增长,3~4级、7~8级表现为正增长。种群龄级变化指数Vpi=27.84%,表明种群为增长型种群[15,25];Vpi’=1.74%>0,说明种群仍稳定;P=0.062 5,抗风险能力较弱。
3.2 生命表与存活曲线
年龄结构、静态生命表、种群动态预测都是基于实际数据,能够反映种群的基本属性,结果也符合种群的自然动态规律[5,20]。实际编表中,利用“空间推时间”,对于许多长寿命多年生种群有意义。实际调查数据不可能完全满足编制特定生命时间表的3个假设,会出现死亡率为负的情况。S.D. Wretten(1980)等认为“生命表分析中产生一些负的dx值,这与数学假设技术不符,但仍能提供有用的生态学记录,即表明种群并非静止不动,而是在迅速发展或衰落之中”[21]。匀滑技术(smooth out)[12]是种群研究学者们常用的方法。由于马鹿河红椿种群的第2到第7龄级出现了死亡率为负的现象。因此本文借鉴S.D. Wretten的技术,对ax进行了匀滑处理,用处理后的ax’编制生命表。
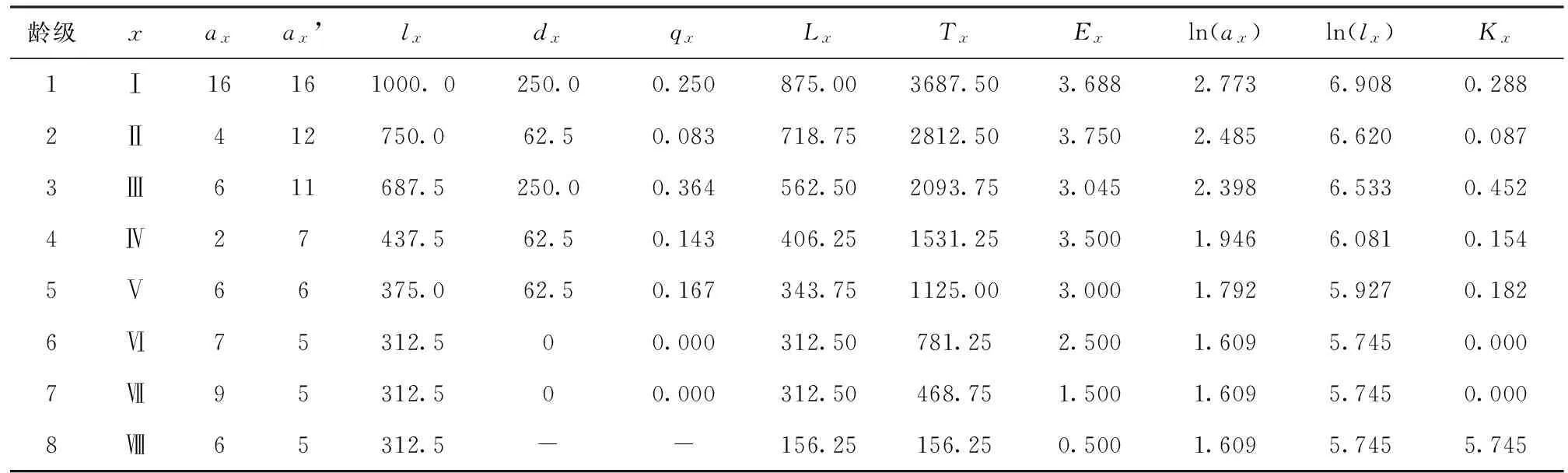
表2 马鹿河红椿种群静态生命表
由表2可知,红椿种群的维持依赖于高龄级个体。第1~2龄级死亡率最高,表明幼苗到幼树、3~4龄级受到环境影响限制。种群发展过程中,在经过2个波动的高死亡率后,形成数量较为稳定的高龄级种群。根据红椿的生理特性,幼苗有较强的耐阴能力,一旦进入第2~3龄级,强阳性的特性成为抑制种群的因子。随着种群的发展,必然出现中间龄级缺失的结构,种群逐步趋向老龄化。
以不同龄级存活量的对数值为纵坐标,以龄级为横坐标作存活曲线 (图2)。存活曲线是反映种群个体在各年龄级的存活状况曲线,借助于存活个体数量来描述特定年龄死亡率[22]。 种群存活曲线从第3龄级后存活数呈明显下降趋势,环境筛的选择强度较大,死亡率高达36.4%,第6~8龄级趋于为0,群落已经摆脱环境筛的作用,进入稳定状态。
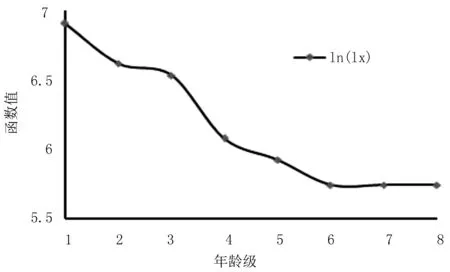
图2 红椿种群成活曲线Fig.2 Survival curve of Toona ciliata population
Hett和Loucks在检验估算的存活状况时,分别用指数函数方程式Nx=N0e-bx和幂函数Nx=N0x-b方程式对Deevey II型曲线和Deevey Ⅲ型曲线进行检验[23,24,25,26]。观察发现,马鹿河红椿种群存活曲线介于Deevey Ⅱ型和Deevey Ⅲ型之间。采用Deevey型曲线对红椿种群存活曲线类型进行检验,并进行拟合得到:
Nx=15.523·e-0.168xR2=0.880F=44.155
Nx=16.496·x-0.615R2=0.977F=252.834
2组方程的检验结果显示,幂函数方程的F值和R2值都大于指数函数,说明红椿种群存活曲线更趋近于Deevey Ⅲ型。表示幼苗的死亡率高,之后的死亡率逐渐走低继而稳定。
3.3 死亡率与损失度分析
种群死亡率曲线反映种群死亡率的动态变化过程[18,19]。
图3表明,红椿种群死亡率曲线与亏损率曲线基本一致,死亡率与损失度可以很好地相互解释。从l~6龄级,2条曲线波动差异不大,但损失度在各个龄级中均略高于死亡率。死亡率的变化说明:第一次死亡高峰期出现于第1龄级幼苗向第2龄级幼树的生长过程中,高达25%,只有75%幼苗可以进入更高龄级。由于生境条件和干扰因子的存在,在植物的生活史中,种子萌发、幼苗的生长和生存被认为是植物最脆弱的阶段[27,28],幼苗生长和竞争能力都不强。红椿种群第3~4龄级阶段只有63.6%进入成树。表明此时可能环境筛的选择强度更高,可能人为干扰造成,或两个因素综合作用。总体而言,进入成树期,只要穿越环境筛,种群即进入平衡。马鹿河种群第4龄级数量仅占种群总数量的3.57%,造成种群中龄级谷值最低。因此,红椿需强光照的生理特征成为红椿种群濒危的最主要因素。
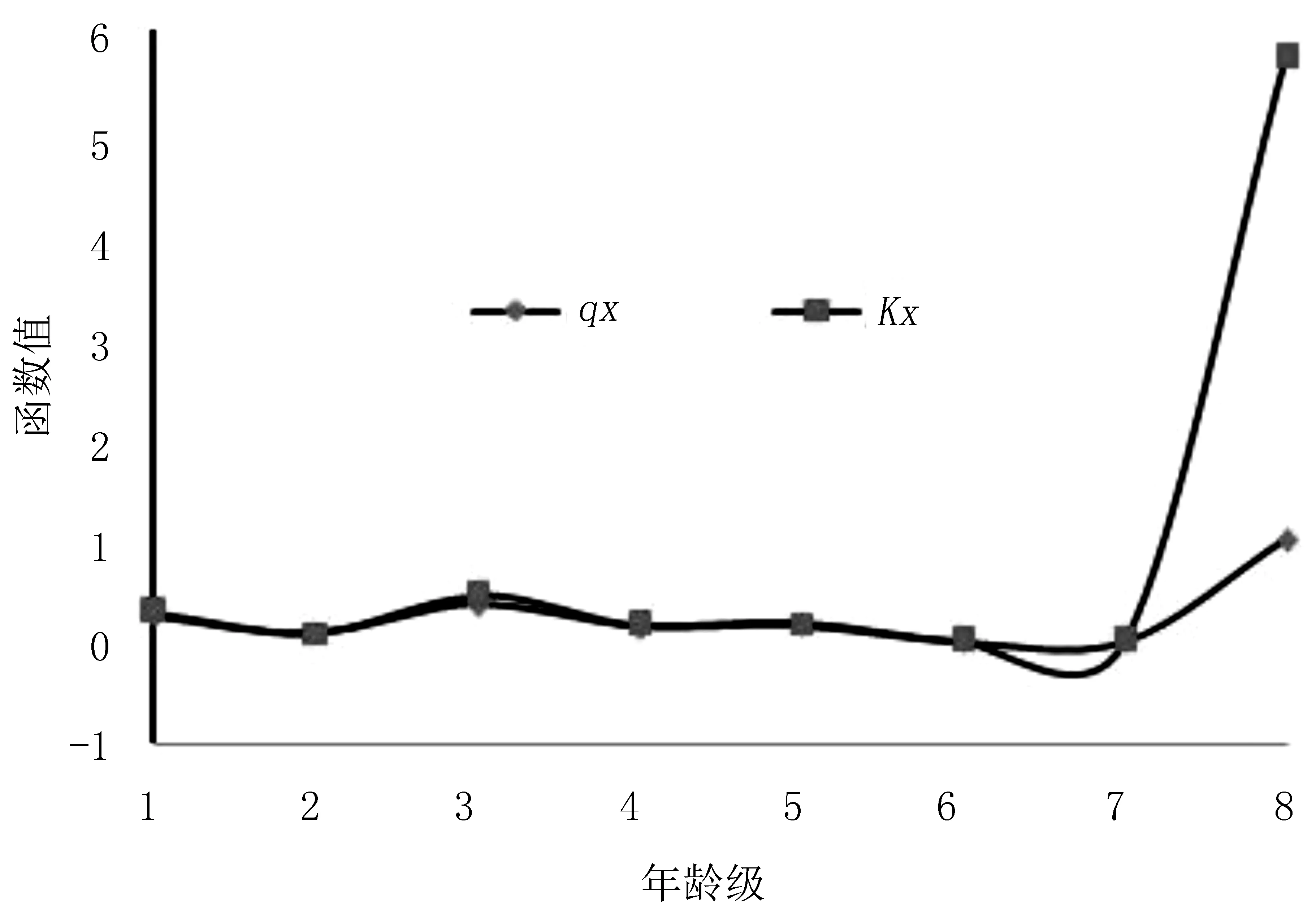
图3 红椿种群死亡率与损失度曲线Fig.3 Death rate and vanishing value of Toona ciliata population
3.4 生存分析
生存分析函数估算值见表3,红椿种群生存分析函数曲线见图4所示。
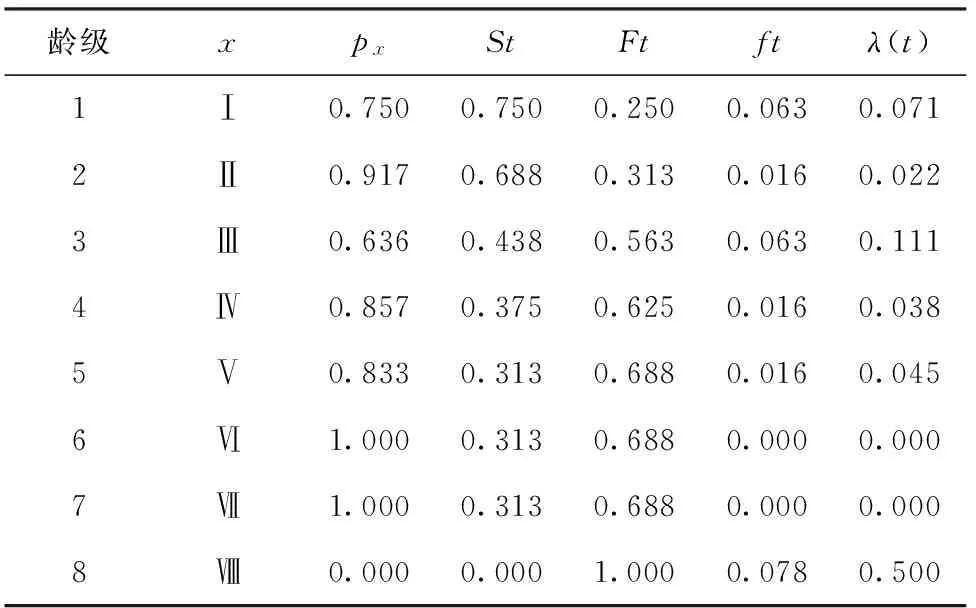
表3 生存分析函数估算值
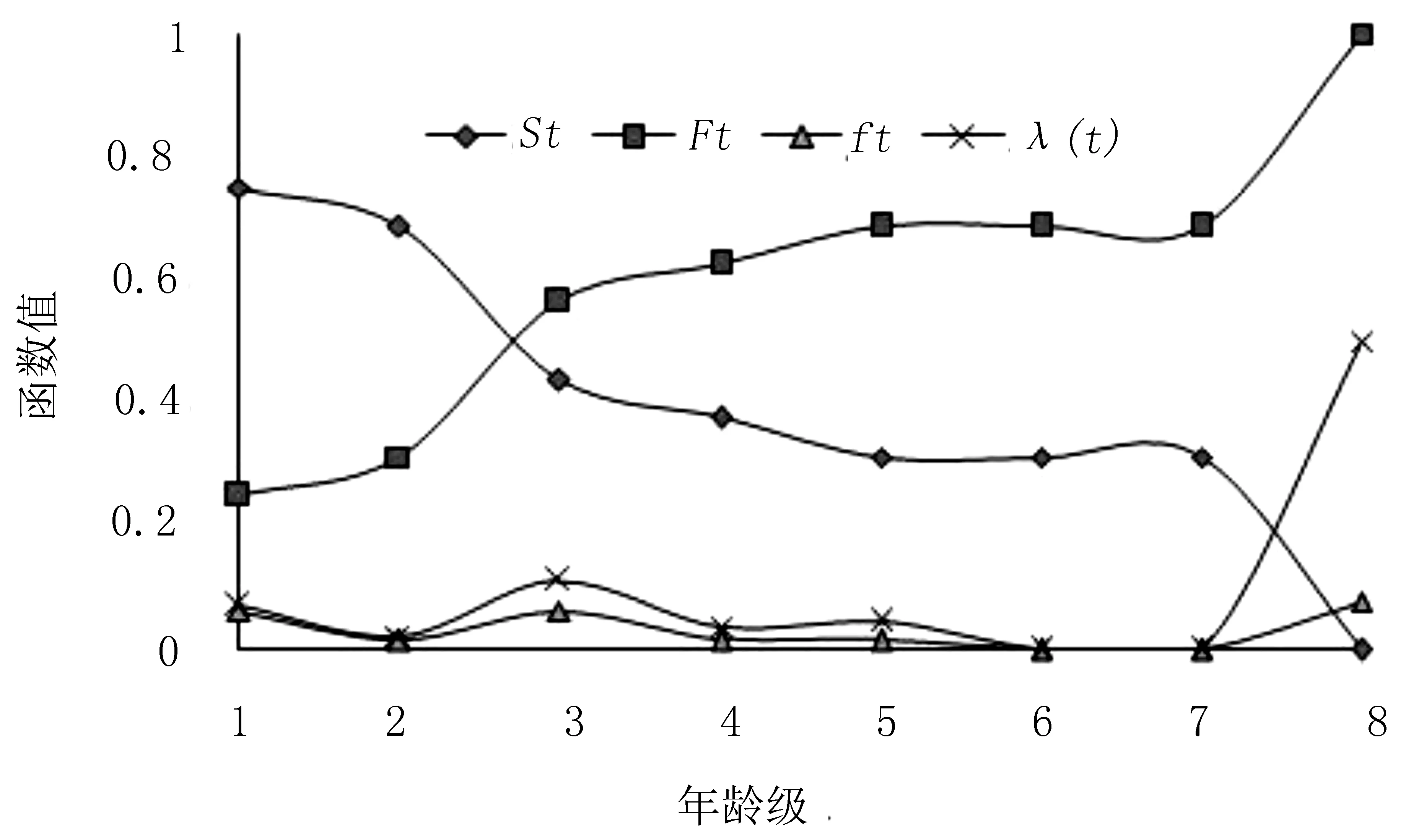
图4 红椿种群生存分析函数曲线Fig.4 Survival analysis functions of Toona ciliata population
由图4可知,红椿种群的生存率函数St、累积死亡率函数Ft互补。St呈单调下降趋势,最高出现在幼苗期,达到0.75;下降趋势在第2向第3龄级发育最明显,与生命表中qx在相应龄级的变化一致。Ft累计死亡率呈单调增加趋势,在4龄级后逐渐趋稳。 2个函数前期的变化幅度大于后期,随着龄级增加,种群存活率S(t)下降,趋向0,累计死亡率F(t)上升,趋向1。死亡密度函数f(t)波动出现3个高峰,在第1和第3龄级,分别为0.063和0.063;在6和7 2个龄级均为0,第8龄级再次上升到最高,为0.078。这可能是由于种群数量下降到一定程度,种群的密度下降导致死亡密度下降,种群稳定后进入新一轮竞争造成。危险率函数λ(t)在第3龄级时出现一个高点,为0.111,说明了种群中期发育阶段的高死亡率。λ(t)随着龄级增大,到6和7 2个龄级为0,表明种群逐渐稳定;第8龄级上升到最高值0.5,预计与种群新竞争有关。4个生存函数估算结果表明,红椿种群具有前后期锐减,中期平稳的特点。这与种群生命表数据分析吻合。
4 讨论与结论
种群的龄级结构、生命表、存活曲线和死亡率曲线是物种种群生物学特性与环境因素共同作用的结果[29]。红椿种群数量动态变化趋势表明,随着时间的推移,种群稳定缓慢增长,是一种两端高中间低的稳定,对抗外界风险概率只有0.025,风险能力较低。各龄级数量显示,幼苗(1龄级)个体数量较多,中间龄级出现幼苗幼树(2~4龄级)的连续损失现象,成树龄级个体数量稳定。排除人为干扰,种群在幼苗和进入幼树阶段完成自疏,使得中间龄级植株丧失较多,进入成树的数量减少,因而种群老化问题日趋严重。种群生命表与存活曲线分析表明,红椿整体上存活曲线属于Deevey-Ⅲ型。死亡率与损失度曲线与种内竞争产生自疏现象基本吻合。4个生存函数估算结果与种群生命表、存活曲线、死亡率曲线和损失度曲线分析结果一致。
濒危物种除人为干扰外,存在自身生理特点和对生境条件高敏感性,独特的生理特点正是环境选择的结果。红椿种群低龄级的高死亡率,也验证了1和4 2个龄级阶段是个体数量变化最大的时期。红椿为样地群落的优势种,但红椿有依赖扰动更新的特性,即靠林窗机遇更新。使得红椿在由不同时空特性的同生群斑块组成中表现出较差连续性。其原因如下:
1) 红椿种子结实率高,随风传播。存在环境仅为岩层地貌,土层较薄,难以进入土壤种子库,导致大量的种子不能发育成幼体。红椿具有很强的萌蘖能力,林下幼株中有一定数量根蘖苗。因此,种子和根系萌蘖繁殖是红椿种群的主要更新方式。
2) 红椿幼树比较耐荫,随着生长,植株对光照需求逐渐加强。在天然林中,幼树必须通过光照环境筛的严格筛选,才能进入主林层。由于天然林内缺乏人为的“积极干扰”,大部分幼树在光照环境筛的选择中逐渐死亡。
3)红椿的天然更新强弱与林分类型有关。林窗能够为红椿的种子萌发和幼苗生长提供较好的条件,促进其生长发展。但当林窗的更新完成后,再次对幼苗进入幼树期、幼树进入成树造成极大的抑制作用。并且在成树阶段稳定期后,种群会进入新一阶段的竞争。
基于上述特点,在具体保护管理措施上采用不同措施:人工辅助育苗,提高种子向幼苗的转化率,适当疏伐以改善群落光环境,促进无性幼苗的生长,控制非自然干扰,形成龄级合理的种群结构。
[1] 中国树木志编委会.中国主要树种造林技术[M].北京:中国林业出版社,1981.
[2] 郑万钧.中国树木志(第1卷) [M].北京:中国林业出版社,1983: 797.
[3] 马献良.红椿播种育苗初步研究[J].安徽林业科技,2005,127(4):7-8.
[4] 汪 洋,冉勇军,冷艳芝,等.红椿和毛红椿遗传与生理生态相关研究[J].安徽农业科学,2014,42(20):6910-6912.
[5] 刘贵峰,臧润国,成克武,等.天山云杉的种群动态[J].应用与环境生物学报,2011,17(5):632-637.
[6] 刘金福,洪伟,吴则焰,等. 孑遗植物水松(Glyptostrobuspensilis)种群生命表和谱分析[J].武汉植物学研究,2008,26(3):259-263.
[7] 胡尔查,王晓江,刘永洪,等.乌拉山自然保护区油松种群结构与分布格局研究[J].林业科学研究,2011,24(3):363-369.
[8] 陈绍林,张志华,廖于实,等.星斗山自然保护区秃杉原生种群生境现状及保护对策[J].安徽农业科学, 2008,36(11):4624-4625.
[9] 刘 毅,孙云逸,陈世明,等.湖北星斗山国家级自然保护区生物多样性分析与保护[J].湖北林业科技,2007(6):42-45.
[10] 戴其生,张梅林,徐玉伟,等.红楝子人工造林试验初报[J].安徽林业科技,1997,34(1):33-34.
[11] 龙汉利,冯毅.四川盆周山地红椿生长特性研究[J].四川林业科技,2011,32(3):37-41, 68.
[12] 江 洪.云杉种群生态学 [M].北京:中国林业出版社,1992:8-26.
[13] 吕海英,王孝安,李进,等.珍稀植物银砂槐中国分布区的种群结构与动态分析[J].西北植物学报,2014, 34(1):0177-0183.
[14] 康华靖,陈子林,刘鹏,等.大盘山自然保护区香果树种群结构与分布格局[J].生态学报,2007, 27(1):389-396.
[15] 汪洋,冷艳芝,苏长江,等.恩施红椿天然种群结构及空间分布格局[J].浙江农林大学学报,2016,33(1): 17-25.
[17] LEAK W B.Age distribution in Virgin red spruce and Northern Hard woods [J].Ecology,1975(56):1451-1454.
[18] 洪 伟,王新功,吴承祯,等.濒危植物南方红豆杉种群生命表及谱分析[J].应用生态学报,2004,15(6):1109-1112.
[19] 吴承祯,洪伟,谢金寿.珍稀濒危植物长苞铁杉种群生命表分析[J].应用生态学报,2000,11(3):333-336.
[20] 李 妍,李登武,韩东辰.贺兰山东坡青海云杉种群动态[J].浙江农林大学学报,2014, 31(1):50-56.
[21] WRETTEN S. Field and Laboratory Exercises in Ecology [M]. London:Edward and Arnad Publishers Limited, 1980: 44-63.
[22] 刘 洋.小兴安岭山杨种群生命表及生存分析[J]. 林业科技,2010,35(2):15-18.
[23] 胡喜生,洪 伟,吴承祯,等.木荷天然种群生命表分析[J].广西植物,2007,27(3):469-474.
[24] 万志兵,程国源,丁强强,等.绩溪光皮桦次生林的种群动态[J].东北林业大学学报, 2014,42(1):27-30.
[25] 杜兴乔,刘映良.荔波瘤果茶种群生命表分析[J] .安徽农业科学,2009,37(33): 16613-16615.
[26] 郭 华,王孝安.子午岭马栏林区油松种群结构分析[J]. 陕西师范大学学报(自然科学版), 2012, 40(1): 67-71.
[27] GRUBB PJ.The maintenance of species-richness in plant communities:the importance of the regeneration niche[J].Bio Rev,1977(52):107-145.
[28] 游水生,叶功富,徐俊森,等.福建东山岛海岸带潺槁树种群生命表分析[J] .广西植物,2009,29(1):96-102.
[29] 黄 刚,杜珍珠,孙肖宁,等.准噶尔盆地南缘小叶桦种群生命表与生存分析[J].江苏农业科学,2012,40(9):310-313.
Life Table and Survival Analysis of EndangeredToonaciliatePopulation
WANG Yang1, HUANG Fang1, ZHANG Hanju1, CHENG Dehua2, XU Xiaoting1
(1. Hubei Vocational College of Ecological Engineering, Wuhan 430200, China; 2. Xiangtan No. 1 High School, Xiangtan, Hunan 411100, China)
Based on sample-plot survey and data statistics ofToonaciliatapopulation in Shengjiaba of Enshi City, the life table ofToonaciliatawas made to analyze and research the dynamics of population quantity. The results showed that population dynamics valueVpi=27.84%>0 indicated a growth form of the population andVpi′=1.74%>0 indicated the a stable form of the population with low risk-resistance capacity. The life table and the survival curve of the population proved the tendency of quantity dynamics in accordance with that of Deevey-Ⅲ. The two peaks of death rates were 0.250 between 1 and 2 age classes, and 0.364 between 3 and 4 age classes, and the maximum loss index fell in 3rd age class with the maximum value 0.452. The peak of survival rate function was 0.75 in 1st age class, decreasing monotonically, while the accumulated mortality rate function approached 1 in 8th age class, increasing monotonically. The mortality density functionf(t)reached 3 peaks, which remained 0.063 in 1st age class, 0.063 in 3rd age class, 0 in 6th and 7th classes and 0.078 in 8th age class. The hazard rate functionλ(t) reached 0.111 in 3rd classes, fell to 0 in 6th and 7th classes but climbed to 0.5 in 8th class. The results indicated that the loss ofToonaciliatapopulation from environmental sieve and disturbances was heavy in seedling and sapling stages, which was one of the principal factors forToonaciliatapopulation′s extinction. The adult population stayed temporarily stable, and then involved in a new round of competition.
Toonaciliata; population; life table; survival analysis
10.3969/j.issn.1671-3168.2017.04.004
S718.54;S759.94
A
1671-3168(2017)04-0017-06
2017-05-16.
湖北省科技厅公益研究项目 (40 2012DBA40001);湖北省教育厅科研项目(B20160555).
汪 洋(1968-),男,湖北武汉人,副教授.主要从事生态学与植物种质资源的教学与研究.
徐晓婷.
