番茄根系对控释氮素的响应及其理想构型
2017-10-19杨俊刚廖上强孙焱鑫李艳梅邹国元
杨俊刚廖上强孙焱鑫李艳梅邹国元
番茄根系对控释氮素的响应及其理想构型
杨俊刚1,2廖上强1,2孙焱鑫1,2李艳梅1,2邹国元1,2
〔1北京市农林科学院植物营养与资源研究所,农业部都市农业(北方)重点实验室,北京 100097;2北京市缓控释肥料工程技术研究中心,北京 100097〕
我国设施蔬菜生产中氮肥环境污染问题十分突出,实现水肥资源高效利用与产量共同提升具有重要意义。根系吸收效率低下是阻碍设施番茄高产高效的一个重要原因。本文结合根系构型的最新研究进展,论述了生产中番茄根系的分布特点及其在控释氮素供应条件下的根系形态响应,分析了控释肥料和常规化肥一次性施肥形成的时空异质性养分供应对根系生长与分布的影响,提出利用控释尿素与普通肥料共同调控番茄根系形成表层密集主根深入的理想根系构型。在此基础上通过地上部群体结构调节构建根层连续体,可以在蔬菜集约化生产中进一步促进植株根系对氮素的吸收,减少氮素淋洗损失,进而在氮肥减施条件下实现番茄高产。
根构型;控释尿素;氮素吸收;硝态氮淋洗
我国是世界上最大的设施蔬菜生产国,种植面积近400万hm2;但是生产中管理粗放、水肥过量投入的现象十分普遍,造成严重的资源浪费和环境污染问题(Shi et al.,2009;喻景权和周杰,2016)。如何以更少的水肥资源和劳动力投入生产出更多的优质农产品,实现环境保护与资源高效利用相协调成为当前迫切需要解决的问题。氮磷化肥减量施用能够显著降低设施蔬菜体系的氮素淋洗损失,进而减轻施肥对环境的污染(Vazquea et al.,2005;Zhu et al.,2005;Ren et al.,2010),但是蔬菜增产并不明显,有的甚至出现减产(Song et al.,2009;Min et al.,2012;Zhao et al.,2012),限制了减肥技术的推广应用。设施蔬菜生产中,追求产量以提高收入始终是菜农的首要目标,因此化肥减量后氮素的合理分配和高产高效就成为一个难点。改善根土互作环境、提升根系吸收的生物学潜力可能是解决这一难题的根本途径。因此,如何提高作物根系的养分、水分吸收效率,尤其是如何调控根系构型来提升茄果类蔬菜的产量显得十分重要。
根系是作物吸收养分的主要器官,其构型变化是增加养分吸收的重要生物学潜力。Hammer等(2009)研究表明,美国现代玉米品种根构型的改变促进了养分和水分的高效吸收,提升了玉米持续增产的能力;栗岩峰等(2006)研究发现,直径小于1 mm的根系占比与番茄产量显著相关;米国华等(2010)认为,深根型玉米品种是协调高产与氮素淋洗控制的重要育种方向。这些研究结果均表明根构型在作物高产高效中的重要作用,但是设施栽培条件下番茄根构型的变化与养分、水分互作的研究还不深入。以往的研究多集中在水氮供应对根系数量特征与产量的影响上,很少考虑高投入体系中氮素淋洗的问题,对于新型缓(控)释肥料与高效吸收根构型的研究还很少,因此深入研究番茄根系与控释氮素的互作效应,阐明根系形态与生理塑性响应以及调控途径,构建高效吸收根系构型,对于促进集约化番茄生产可持续发展具有重要意义。
1 设施番茄氮素损失与根系分布特点
设施蔬菜生产中氮素损失十分严重。梁静等(2015)分析了79个设施番茄施肥试验,结果表明我国菜农传统管理下的氮肥利用率还不到10%,大量氮素以硝态氮形式发生淋洗,损失率高达19.7%~32.8%(Kraft & Stites,2003;Song et al.,2009;Min et al.,2012),较高的淋洗损失也被认为是过量施氮而不能增产的根本原因(Zotarelli et al.,2009a)。菜地养分大量损失与作物根系吸收效率低以及分布不合理有直接关系。过量施肥形成的高氮浓度抑制植物体内生长素(IAA)的合成和向根系运输,根系感受到低IAA信号则会抑制侧根原基形成以及侧根伸长,导致根系表面积减小、侧根密度降低,根系吸收效率下降(Walch-liu et al.,2001;Tian et al.,2008)。大水大肥会增加中后期番茄根系在深层土壤中的分布,这可能与大量硝态氮淋洗到下层有很大关系,但这种分布没有引起氮素吸收增加和硝态氮淋洗损失减少(Zotarelli et al.,2009a,2009b)。Jackson和Bloom(1990)认为番茄根系深入下层与养分富集的土壤表层在空间上错位是导致其氮素利用率低下的重要原因。除此之外,在番茄栽培中采用的稀植和对称栽培法,虽然有利于通风透光促进个体生长,但往往会产生根系重叠竞争区和少根间隙区,这种不合理的根系分布也会降低氮素的吸收,使之随灌水发生淋洗。笔者在基质栽培条件下发现,采用常规施肥的番茄主根和粗根多,养分吸收活力强的细根少,侧根增生受到抑制(图1-A);而施用控释肥的番茄侧根发达,细根布满表层,有利于增加养分的吸收(图1-B)。这也进一步表明番茄根系吸收效率亟待提升。

图1 不同养分分布方式下番茄根系纵向剖面图
2 减少氮素淋洗、提高氮素吸收的高效根构型
植物根构型是指根系在空间上的分布结构以及侧根的数量和长度(Hodge et al.,2009)。养分的高效利用不仅依靠植物根系数量的增加,而且与根系的三维构型、侧根密度以及空间分布存在密切关系(Williamson et al.,2001;Garnett et al.,2009)。研究表明,形成理想的或者最适的根构型是进一步提高养分有效性的一条重要途径(米国华 等,2010),因此许多研究者致力于根构型的研究来提高作物产量和水肥利用效率(Lynch,1995;King et al.,2003;Zotarelli et al.,2009a)。米国华等(2010)提出一种深根型玉米理想构型,认为增加深层土壤中的根系分布,有可能减少氮素向深层的淋失,从而同步实现玉米高产与氮素高效利用,这一观点与King等(2003)的研究结论相似。但也有研究认为,在作物生育早期增加土壤表层根系密度,可以减少中后期氮素的大量淋洗,并提出表层根系密集、主根深入的理想根构型观点(Dunbabin et al.,2003)。与增加侧根数量不同,在“steep-deepcheap”的玉米理想根构型中,Lynch(2013)提出增加侧根长度来提高根长密度,促进水分养分吸收效率提高的设想。由此可见,调控侧根形态变化在增强根系吸收能力上具有重要的作用。
根系构型的改变往往受基因表达以及环境因子的影响。最近几年,应用数量性状基因定位(QTL)方法,寻找控制加深根系或促进细根密度增加、侧根增生等的基因是当前的研究热点(Wachsman et al.,2015)。例如Uga等(2013)在水稻中克隆到1个控制深根的QTL微位点基因Drol,将该基因导入到浅根型水稻中可以显著提高根系在深层土壤中的分布,并表现出氮素高效吸收的特性。根系构型的改变往往由数十个或上百个基因控制,准确找出一系列的控制基因面临的挑战非常大。目前基于磷素有效性的QTL报道较多,但在氮有效性基因调控上的研究相对较少。除了上述水稻深根基因成功应用外,迄今在基因水平上的研究几乎没有新的应用(Li et al.,2016)。基因水平上的根系调控研究有助于理解养分高效吸收的分子与生理机理,最终用于分子育种,但通过转基因育种改变根构型来提升养分吸收效率,尚有许多未知需要长期的探索。
环境养分对根构型的影响以及植物生理响应的发现也为根系构型的调控提供了理论支撑。设施生产中过量施肥形成的高氮环境会对根系生长和分泌物产生抑制和损害,进而降低养分利用效率(Teng et al.,2013;Deng et al.,2014)。在养分供应强度和供应形态上的调控往往会改变根系对养分的活化和吸收。例如在田间条件下,土壤硝态氮浓度与玉米根长存在显著的相关性(Shen et al.,2013);局部供应磷肥和氨态氮可以促进玉米根系增生和根际酸化,进而在石灰性土壤上提高养分的吸收量(Jing et al.,2010,2012;Ma et al.,2013)。近年来,基于根层养分调控的土壤—作物系统综合管理技术,使得我国三大粮食作物产量得到大幅度提高,同时大幅度减少了单位籽粒产量的活性氮及温室气体排放量,在大区域范围内实现了减少投入、增加作物产量的可能性(Chen et al.,2014)。
基于前人的研究,要在高产的前提下实现养分高效,考虑根系分布及构型非常必要,尤其是在个体根构型的基础上,进一步明确高产群体根层高效养分利用机制与调控更为重要。因为当存在新的养分竞争者时,根系吸收特点会发生明显的改变(Cahill et al.,2010),出现适应性塑性反应(王鹏 等,2012),因此对一定群体密度的根层结构研究显然更有意义。目前有关番茄理想根构型与群体生产能力的研究还相对滞后,在设施生产条件下,番茄灵活的畦垄栽培使得群体根系分布形成根层连续体成为可能,明确其高效结构与调控途径将为控制氮素损失并进一步挖掘高产潜力提供理论与技术支持。
3 控释肥对根构型的调节及番茄高产高效的作用
肥料不仅是养分供应源,而且具有调节植物根系分布和生长的重要作用(Shen et al.,2013)。养分的异质性分布会改变根系的形态塑性和生理塑性,进而增加根系分布和地上部生物量(Hodge,2004;Hendriks et al.,2015)。Hodge 等(2009)总结了前人的大量研究,认为侧根增生有利于形成吸收活力强的细根,进而提升根系吸收效率,但局部氮素供应过高或过低都会抑制侧根增生。与大田作物相比,蔬菜根系对养分的捕获能力和塑性响应研究还不深入,关于番茄根系吸收效率提升途径还不清楚。设施菜田过量施用普通氮肥会在苗期形成较高的氮素浓度不利于侧根增生,而后频繁大量的灌水加速了氮素淋洗,使施肥形成的养分斑块很快在空间上发生变化,供氮强度和空间位置对根系的影响没有持久性。如果养分移动速度超过根系增生的速度,根系的局部增生会出现不良反应,无法达到预期的调控目标(Lynch,2013)。笔者认为控释肥可以改变这一状况。控释氮肥改变了氮素的供应规律,大幅度降低了前期供氮强度,而施入土壤后始终固定在起始位置,在供氮强度和空间上更有利于对根系的调控。因此,从这个意义上讲,控释肥完全可以作为一种全新的根系调节手段,在时空维度上形成异质性养分供应,进而对根系构型产生影响。前人研究指出,控释肥在接触施肥时(控释肥条施或穴施,覆土3~5 cm后直接播种或移栽秧苗)可以提高水稻的氮素吸收(Tian & Saigusa,2002);但在玉米上的研究结果表明,接触施用控释尿素虽然增加了表层土壤的根长密度,但肥料离种子太近致使部分根系出现黄化烧根现象(杨俊刚 等,2010)。Tian和Saigusa(2005)发现将控释肥装袋竖直埋入距番茄植株一侧20 cm时,肥料袋周围细根大量增生,根长密度显著增加。由此可见,控释肥在根系调节中的重要作用,尤其是促进细根比例的增加值得深究。施用控释肥虽然引起了根系构型上的改变,但增产效果并不明显,因此需要寻找一种理想的根系构型,增加养分吸收并实现增产。
朱晋宇等(2015)认为番茄根系横向扩张与纵向深入均衡发育可以增加氮素吸收,但根系仅仅是分布较深而横向半径减小则氮素吸收也会下降。栗岩峰等(2006)研究表明,番茄细根比例的增加与产量呈正相关关系。根系直径越细活性越高,单位根长的吸收效率越高(李春俭,2001;李洪波 等,2013),直径小于0.5 mm的根系呼吸速率是粗根的2.4~3.4倍(Pregitzer et al.,1998),说明调控表层细根对于提升吸氮数量极其重要。控释尿素颗粒进入土壤后其空间位置固定,养分释放常常可以持续2~3个月,采用适当的施肥方法完全可以决定氮素的分布空间,进而影响细根增生的空间位置。杨俊刚等(2010)将控释肥均匀施在0~20 cm土壤中,不同土层番茄根长密度的变化结果表明,随施氮量降低0~15 cm土层根长密度下降了4%~31%,但在15~30 cm土层控释肥处理根长密度不降反升,比其他2个处理增加8.0%~60.0%。这一结果与栗岩峰等(2006)在番茄根系分布上的结果相似,随施氮量下降根长密度减少,调控水氮施用顺序可以增加下层(20~40 cm)细根的密度。然而在这些研究中,增加下层根长密度并没有提高吸氮量,根系吸收效率几乎没有改变,说明细根在下层增生对番茄来说并不是理想的选择。Kristensen和Thorup-Kristensen(2007)研究了几种蔬菜根系对氮素垂直分布的响应,结果表明与深层硝态氮含量增加相比,浅层硝态氮含量增加更能促进根系对氮素的吸收。与玉米深根吸氮不同(米国华 等,2010),番茄根系90%以上分布于0~40 cm土层内(Oliveira & Calado,1996; 栗 岩 峰 等,2006;Zotarelli et al.,2009a),下扎深度比玉米根浅了许多(Marschner,2012),深根型可能是玉米的理想根构型,但并不适用于番茄。因此,通过增加深层根系来阻控番茄体系中硝态氮淋洗的可能性很小。已有研究发现,0~15 cm土层根长密度增加可能与土壤剖面硝态氮淋洗损失有关,且伴随番茄产量的显著增加(Zotarelli et al.,2009a)。虽然还没有证据表明上层根系增多是增产的直接原因,但这些结果给了我们一个启示,增加上层土壤根系分布密度和横向扩张能力可能是提升根系吸收效率的一条有效途径。为此,笔者采用新的施肥方法,将具有“S”型释放特征的控释尿素与其他肥料分开施用,专门施在土壤浅表层(0~10 cm),改变了肥料颗粒在土壤中的分布位置,结果促进了番茄根系在表层土壤中大量增生,直径小于1.0 mm细根比例显著增加(图2-A、B)。与普通肥料相比,控释肥促进细根增生的特性为进一步研究番茄根系生物学潜力提供了新的可能,也为构建高产番茄理想根构型提供了新的思路。
“S”型控释肥(曹兵 等,2010,2012)在作根长物生长前期养分释放较慢,而在后期供应不断增强,与作物生长曲线趋于同步,对于根系合理分布和侧根增生也有明显促进作用,有利于进一步增加氮素的吸收形成高产。最近的研究表明,低氮条件下玉米主根生长增强而侧根增生受到抑制,正常供氮时则促进侧根生长(Gao et al.,2015);番茄苗期高氮供应不利于根系深入分布(朱晋宇 等,2015),可见适宜的供氮是形成高效根构型的基础。与习惯施肥相比,采用控释肥与常规氮肥混配一次性施用可以降低生长前期氮素供应浓度和数量,有利于主根下扎,而后在生长中后期随控释氮释放增加则会促进侧根增生,有利于构建主根深入表层侧根密集的结构,为促进根土高效互作和降低施肥成本提供了一条新途径。
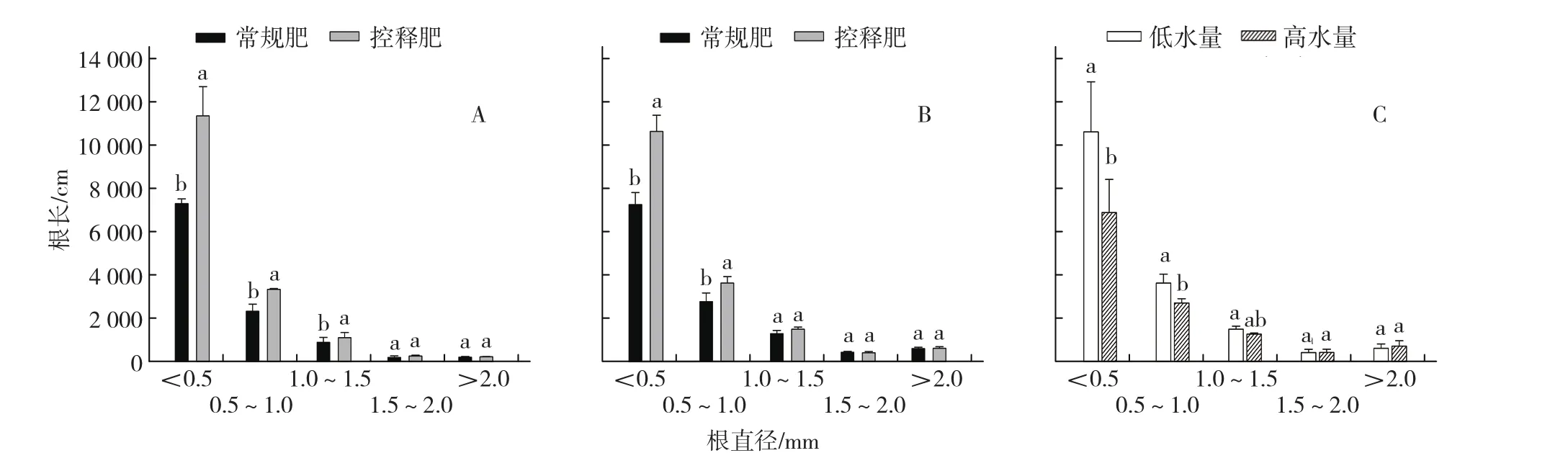
图2 不同施肥方式及灌水量对番茄根长、直径分级的影响
控释氮素需借助水分才能释放,土壤水分也是影响根系生长的重要因素,控释肥与水分互作对根系的调控也就十分重要。一般情况下,减少灌溉量有利于根系增加吸收面积和向深层生长。研究表明,与高水量相比,低水量条件下控释肥处理的玉米根系干质量增加77%,土壤深层分布比例增加,普通尿素也有同样的趋势,但变幅较小(邵国庆,2008)。草莓根系活力在适度节水(减量25%)和中等供应控释肥强度(120~160 kg·hm-2)下达到最大(王翠玲,2010)。节水28%~42%的交替灌溉与施用控释肥的组合可以显著增加水稻产量,其中根系生物量的增加为增产打下了良好的基础(Tuong et al.,2002)。可见,在低水量条件下施用控释肥可以促进根系的生长和活力提高。笔者前期的试验也说明了这一点,低水量下,与常规施氮相比,控释肥可以促进细根增生,而在高水量条件下控释肥处理的细根比例有所降低(图2-C)。因此,节水条件下更加有利于控释肥对根系的调控。然而在极度干旱胁迫下会产生控释氮释放延迟或作物生长期推迟的现象,可能造成氮素供应与作物需求无法同步,影响根系的响应和分布(Ye et al.,2013)。与露地栽培相比,设施生产中灌溉条件便利,出现极度干旱的情况非常少,但要注意避免长期缺水。
减少硝态氮淋洗是实现作物高产与氮肥高效的另一重要环节(米国华 等,2010)。施用控释尿素可以显著减少硝态氮淋洗损失,由于蔬菜根系分布相对较浅,控释肥不用深施即可形成一个与根系空间一致的稳定的养分分布,促进高效吸收的同时抑制硝态氮淋洗(Dunbabin et al.,2003;Garnett et al.,2009)。不仅如此,笔者在控释肥浅表层(5~15 cm)集中施用的基础上同时改进栽培技术,采用“内三角”定植法(小高畦垄上双行种植,幼苗按照一定距离在垄内侧交叉定植,3株呈三角形分布的一种改进的栽培方法)调控番茄群体结构,实现产量提升50%以上(杨俊刚 等,2012,2014;李艳梅 等,2014),突破了前几年番茄产量的低水平徘徊,证明了施用控释肥的高产潜力。根据不同来源养分的供氮特点采用一次性施肥,控释尿素可以为作物花后氮素吸收高峰提供养分,普通肥料则主要在生长前期供应,两种肥料分时协同供应能有效减少氮素的淋洗损失,满足高产作物养分需求与损失控制协调。
4 番茄高效吸收氮素的理想根构型
在分析前人研究结果和总结控释肥养分调控技术的基础上,笔者提出一种番茄理想根构型的假设:即表层根系密集且横向扩张能力强,易于在株间形成连续分布以减少硝态氮的快速淋洗,部分根系深入下层进一步增加水肥的吸收数量(图3)。笔者试图通过应用控释肥将氮素供应在时间和空间上创造出与速效氮肥不同的养分供应环境,来调控和提升根系的吸收效率。设施生产中,以控释肥为核心综合速效氮磷养分,在土壤0~15 cm和15~30 cm上下两层形成时间和空间上的氮磷异质性养分分布。控释肥主要分布在0~15 cm土层且集中在幼苗正下方,通过时空异质性养分供应促进表层细根增生,扩大横向分布;普通氮磷肥和有机肥则分布在0~30 cm土壤中,利用氮磷养分对根系的调节作用促进番茄主根健壮和深入分布,最终形成养分水分高效吸收的理想根构型。施肥可分为两步,首先将有机肥和常规氮磷钾肥撒施深翻,第二步表施控释尿素并与土壤轻轻混拌,然后起高畦将控释肥颗粒翻埋在高畦中底部;接着在高畦上双行错位定植秧苗,形成三角布局,在水平方向减少根系内侧重叠,扩大外侧分布,使株、行两个方向的竞争最小、协同效应最大,最终形成上层交叉密集连续分布,并且深入下层的根土水高效互作综合体,以支撑高产群体,实现养分高效吸收与高产同步。

图3 番茄理想根构型构建模式
根构型是根系在发育过程中不断与环境因子互作形成的,养分供应对根系发育的影响从苗期就已开始。以番茄为例,不同供氮强度对番茄苗期根系三维空间分布与氮素吸收具有显著差异,4 mmol·L-1氮素供应下番茄幼苗根系三维构型半径窄而深度深,而20 mmol·L-1氮供应下根系三维构型半径宽而深度浅,降低供氮强度有利于根系氮素吸收效率的提高,因此建议苗期适当降低施氮量促进根系深入土壤来提高氮素吸收(朱晋宇 等,2015)。这一研究结果与本文提出的前期促进根系下扎的假设是一致的。植株苗期根系发育速度快,随着灌水和作物生长根系构型很快发生变化,当作物由营养生长转入生殖生长一段时间以后,根系生长基本停止(Ye et al.,2013),根系构型基本形成,作物进入大量吸收养分和产量建成阶段。这一阶段也是水肥大量供应时期,此时的根构型是提高水氮吸收率的关键时期,因此本文将这一时期建成的根构型作为理想构型的调控目标。调控过程则需要从苗期开始,适当降低水肥供应强度以保证根系下扎深度。
5 展望
植物根系对不同种类养分的吸收方式并不相同,例如对氮素的吸收主要以质流为主,而对于磷钾的吸收主要扩散为主(Marschner,2012)。质流与土壤水分运移和蒸腾相伴,而扩散主要依靠根系与土壤的接触范围和体积。氮素是最容易移动和损失的养分,所以理想根构型的假设重点考虑了对氮素的吸收,如深根型分布吸收淋洗到下层的硝态氮。理想的根构型不仅能解决氮素的吸收还应提高磷钾的吸收,尤其是需钾较多的果菜类蔬菜。磷钾的移动性差,容易在上层土壤中累积,多年来蔬菜生产富集的磷素已经变成重大的环境风险。本文提出的表层密集型理想构型,不仅考虑了植株氮素吸收效率的提升,也有利于增加对磷钾的吸收。研究表明,通过对氮磷分布的调控,不仅可以显著增加玉米总根长、根干质量和根质量,而且可以显著增加对氮磷的吸收(王昕 等,2013),说明根系不但在形态上做出塑性响应,而且通过生理塑性响应提高了吸收效率,这也进一步支持了笔者的假设。根系构型改变是一个复杂的过程,除基因和土壤环境之外,定植密度、个体大小、农艺措施以及天气等均会产生影响,本文虽然提出一个番茄理想根构型模式,但尚需进一步的数据支持以及田间试验的验证。根系构型的改变是否能提升养分吸收效率,进而实现增产,是判断理想构型的一个重要标准。目前这一方面的进展比较缓慢,但随着生物和基因技术的快速发展,相信通过遗传改良改变根构型的技术会有新的突破;同时,在养分与根系生理方面通过在适宜的时期和位置提供合理的养分和水分条件,利用细根对养分信号的响应,延长作物中后期细根更替的周期,也会提升根系吸收效率,进而转化为作物产量的增加。
未来农业的可持续发展必须在继续增产的同时,还要实现节约资源、保护环境的目标。为实现高产高效的模式,尚需进一步开展的工作可能有:① 系统阐明高投入条件下控释肥促进细根增生的机制及调控途径。根构型研究多集中在自然生态系统或低投入体系中,在集约化设施生产中的研究还不多,进一步阐明控释肥时空异质性氮素供应强度、供应形态和动态对番茄根系形态的调控及氮、磷、钾养分吸收效率的提升作用;加强养分调控器官发育的遗传与分子机制研究,揭示其养分高效与高产的协同关系。② 在应用层面,以个体高效根构型为基础形成的根层连续体对番茄高产群体的支撑作用,不同种植密度、水氮供应对氮素淋洗的阻控及高产能力等需要系统研究。第一次农业绿色革命通过品种矮化显著提高了稻麦等作物在高水高肥条件下的产量。现代农业中,除了发展高产高效品种外,高密栽培将是一项重要的种植模式,其体系中根系竞争与分布调控将变得十分重要。因此,进一步加强密植体系中番茄理想根构型的吸收效率与形态变化的研究十分必要。未来应用作物品种高产高效遗传改良上的技术成果,与精准的田间养分管理技术结合,可以最大限度地发挥根系的生物学潜力,同时实现高产、安全、资源高效和环境友好的目标。
曹兵,徐秋明,刘宝存,杨俊刚,许俊香,李亚星,范京松,郝赟,倪小会,谷佳林,衣文平,杨宜斌.2010.一种S型控释尿素及其制造方法.中国:CN101302131B.
曹兵,杨俊刚,许俊香,倪小会,徐秋明,刘宝存.2012.一种包膜复合肥及其制备方法.中国:CN101462907B.
李春俭.2001.高等植物的矿质营养.北京:中国农业大学出版社:46,356.
李洪波,薛慕瑶,林雅茹,申建波.2013.土壤养分空间异质性与根系觅食作用:从个体到群落.植物营养与肥料学报,19(4):995-1004.
李艳梅,廖上强,薛高峰,陈延华,孙焱鑫.2014.减量灌溉下番茄施控释肥的水氮耦合效应研究.农业环境科学学报,33(1):134-140.
栗岩峰,李久生,饶敏杰.2006.滴灌施肥时水肥顺序对番茄根系分布和产量的影响.农业工程学报,22(7):205-207.
梁静,王丽英,陈清,张卫峰.2015.我国设施番茄氮肥施用量现状及其利用率、产量影响和地力贡献率分析评价.中国蔬菜,(10):16-21.
米国华,陈范骏,吴秋平,赖宁薇,袁力行,张福锁.2010.玉米高效吸收氮素的理想根构型.中国科学:生命科学,40(12):1112-1116.
邵国庆.2008.控释肥和水分调控对玉米氮水利用、产量和品质的影响〔硕士论文〕.泰安:山东农业大学.
王翠玲.2010.水肥耦合对草莓生长、产量品质及水肥利用效率的影响〔硕士论文〕.泰安:山东农业大学.
王鹏,牟溥,李云斌.2012.植物根系养分捕获塑性与根竞争.植物生态学报,36(11):1184-1196.
王昕,唐宏亮,申建波.2013.玉米根系对土壤氮、磷空间异质性分布的响应.植物营养与肥料学报,19(5):1058-1064.
杨俊刚,倪小会,曹兵,许俊香,曹兵,刘宝存.2010.接触施用包膜控释肥对玉米产量、根系分布和土壤残留Nmin的影响.植物营养与肥料学报,16(4):924-930.
杨俊刚,张冬雷,徐凯,倪小会,肖强,曹兵,刘宝存,邹国元.2012.控释肥与普通肥料混施对设施番茄生长和土壤硝态氮残留的影响.中国农业科学,45(18):3782-3791.
杨俊刚,张鹏飞,倪小会,肖强,曹兵,邹国元,刘宝存.2014.施用控释肥对设施番茄NO3--N淋洗、N2O排放及产量与品质的影响.农业环境科学学报,33(9):1849-1857
喻景权,周杰.2016.“十二五”我国设施蔬菜生产和科技进展及其展望.中国蔬菜,(1):18-30.
朱晋宇,惠放,李苗,马韫韬,余宏军,蒋卫杰.2015.氮水平对盆栽沙培番茄苗期根系三维构型与氮素利用的影响.农业工程学报,31(23):131-137.
Cahill J F,McNickle G G,Haag J J,Lamb E G,Nyanumba S M,St Clair C C.2010.Plant integrate information about nutrients and neighbors.Science,328:1657.
Chen X,Cui Z,Fan M,Vitousek P,Zhao M,Ma W,Wang Z,Zhang W,Yan X,Yang J,Deng X,Gao Q,Zhang Q,Guo S,Ren J,Li S,Ye Y,Wang Z,Huang J,Tang Q,Sun Y,Peng X,Zhang J,He M,Zhu Y,Xue J,Wang G,Wu L,An N,Wu L,Ma L,Zhang W,Zhang F.2014.Producing more grain with lower environmental costs.Nature,514:486-489.
Deng Y,Chen K R,Teng W,Zhan A,Tong Y P,Feng G,Cui Z L,Zhang F S,Chen X P.2014.Is the inherent potential of maize roots efficient for soil phosphorus acquisition? PLoS One,9:1-9.Dunbabin V,Diggle A,Rengel Z.2003.Is there an optimal root architecture for nitrate capture in leaching environments? Plant,Cell Environ,26:835-844.
Gao K,Chen F,Yuan L,Zhang F,Mi G.2015.A comprehensive analysis of root morphological changes and nitrogen allocation in maize in response to low nitrogen stress.Plant Cell Environ,38(4):740-750.
Garnett T,Conn V,Kaiser B N.2009.Root based approaches to improving nitrogen use efficiency in plants.Plant Cell Environ,32:1272-1283.
Hammer G L,Dong Z S,McLean G,Doherty A,Messina C,Schussler J,Zinselmeier C,Paszkiewicz S,Cooper M.2009.Can changes in canopy and/or root system architecture explain historical maize yield trends in the U.S. corn belt? Crop Sci,49:299-312.
Hendriks M,Visser E J W,Visschers I G S,Aarts B H J,de Caluwe H,Smit-Tiekstra A E,van der Putten W H,de Kroon H,Mommer L.2015.Root responses of grassland species to spatial heterogeneity of plant-soil feedback.Functional Ecology,29(2):177-186.
Hodge A.2004.The plastic plant:root responses to heterogeneous supplies of nutrients.New Phytologist,162(1):9-24.
Hodge A,Berta G,Doussan C,Merchan F,Crespi M.2009.Plant root growth,architecture and function.Plant Soil,321:153-187.
Jackson L E,Bloom A J.1990.Root distribution in relation to soil nitrogen availability in field-grown tomatoes.Plant Soil,128(2):115-126.
Jing J Y,Rui Y K,Zhang F S,Rengel Z,Shen J B.2010.Localized application of phosphorus and ammonium improves growth of maize seedlings by stimulating root proliferation and rhizosphere acidification.Field Crop Res,119:355-364.
Jing J Y,Zhang F S,Rengel Z,Shen J B.2012.Localized fertilization with P plus N elicits an ammonium-dependent enhancement of maize root growth and nutrient uptake.Field Crop Res,133:176-185.King J,Gay A,Sylvester-Bradley R,Bingham I,Foulkes J,Gregory P,Robinson D.2003.Modelling cereal root systems for water and nitrogen capture:towards an economic optimum.Ann Bot,91:383-390.
Kraft G J,Stites W.2003.Nitrate impacts on groundwater from irrigated-vegetable systems in a humid north-central US sand plain.Agr Ecosyst Environ,100:63-74.
Kristensen H L,Thorup-Kristensen K.2007.Effects of vertical distribution of soil inorganic nitrogen on root growth and subsequent nitrogen uptake by field vegetable crops.Soil Use and Management,23:338-347.
Li X,Zeng R,Liao H.2016.Improving crop nutrient efficiency through root architecture modifications.J Integr Plant Biol,58:193-202.
Lynch J P.1995.Root architecture and plant productivity.Plant Physiol,109:7-13.
Lynch J P.2013.Steep,cheap and deep:an ideotype to optimize water and N acquisition by maize root systems.Annals of Botany,112:347-357.
Ma Q H,Zhang F S,Rengel Z,Shen J B.2013.Localized application of NH4+-N plus P at the seedling and later growth stages enhances nutrient uptake and maize yield by inducing lateral root proliferation.Plant Soil,372:65-80.
Marschner P.2012.Mineral nutrition of higher plants.Third edition.Oxford.UK:Elsevier Ltd Press:317-325.
Min J,Zhang H L,Shi W M.2012.Optimizing nitrogen input to reduce nitrate leaching loss in greenhouse vegetable production.AgrWater Manage,111:53-59.
Oliveira M R G,Calado A M.1996.Tomato root distribution under drip irrigation.J Amer Soc Hort Sci,121(4):644-648.
Pregitzer K S,Laskowski M J,Burton A J,Lessard V C,Zak D R.1998.Variation in sugar maple root respiration with root diameter and soil depth.Tree Physiology,18:665-670.
Ren T,Christie P,Wang J G,Chen Q,Zhang F S.2010.Root zone soil nitrogen management to maintain high tomato yields and minimum nitrogen losses to the environment.Sci Hortic,125:25-33.
Shen J,Li C,Mi G,Li L,Yuan L,Jiang R,Zhang F.2013.Maximizing root/rhizosphere efficiency to improve crop productivity and nutrient use efficiency in intensive agriculture of China.J Exp Bot,64:1181-1192.
Shi W M,Yao J,Yan F.2009.Vegetable cultivation under greenhouse conditions leads to rapid accumulation of nutrients,acidification and salinity of soils and groundwater contamination in South-Eastern China.Nutr Cycling Agroecosyst,83:73-84.
Song X Z,Zhao C X,Wang X L,Li J.2009.Study of nitrate leaching and nitrogen fate under intensive vegetable production pattern in northern China.Comptes Rendus Biology,332(4):385-392.
Teng W,Deng Y,Chen X P,Xu X F,Chen R Y,Lv Y,Zhao Y Y,Zhao X Q,He X,Li B,Tong Y P,Zhang F S,Li Z S.2013.Characterization of root response to phosphorus supply from morphology to gene analysis in field-grown wheat.J Exp Bot,64:1403-1411.
Tian X H,Saigusa M.2002.Merits,utilization,and perspectives of controlled release nitrogen fertilizers.Tohoku Journal of Agricultural Research,52(3-4):39-55.
Tian X H,Saigusa M.2005.Response of tomato plants to a new application method of polyolefin-coated fertilizer.Pedosphere,15(4):491-498.
Tian Q Y,Chen F J,Liu J X,Zhang F S,Mi G H.2008.Inhibition of maize root growth by high nitrate supply is correlated to reduced IAA levels in roots.J Plant Physiol,165:942-951.
Tuong T P,Castillo E G,Cabangon R C,Boling A,Singh U.2002.The drought response of lowland rice to crop establishment practices and N-fertilizer sources.Field Crop Res,74(2):243-257.
Uga Y,Sugimoto K,Ogawa S,Rane J,Ishitani M,Hara N,Kitomi Y,Inukai Y,Ono K,Kanno N,Inoue H,Takehisa H,Motoyama R,Nagamura Y,Wu J,Matsumoto T,Takai T,Okuno K,Yano M.2013.Control of root system architecture by deeper rooting increases rice yield under drought conditions.Nat Genet,45:6.Vazquea N,Pardo A,Suso M L,Quemada M.2005.A methodology for measuring drainage and nitrate leaching in unevenly irrigated vegetable crops.Plant Soil,269:297-308.
Wachsman G,Sparks E E,Benfey P N.2015.Genes and networks regulating root anatomy and architecture.New Phytol,208:26-38.Walch-liu P,Neumann G,Engels C.2001.Response of shoot and root growth to supply of different nitrogen forms is not related to carbohydrate and nitrogen status of tobacco plants.J Plant Nutr Soil Sci,164(1):97-103.
Williamson L C,Ribrioux S P,Fitter A H,Leyser H O.2001.Phosphate available regulates root system architecture in Arabidopsis.Plant Physiology,126:875-890.
Ye Y S,Liang X Q,Chen Y X,Liu J,Gu J T,Guo R,Li L.2013.Alternate wetting and drying irrigation and controlled-release nitrogen fertilizer in late-season rice:effects on dry matter accumulation,yield,water and nitrogen use.Field Crop Res,144:212-224.
Zhao Y,Luo J H,Chen X Q,Zhang X J,Zhang W L.2012.Greenhouse tomato-cucumber yield and soil N leaching as affected by reducing N rate and adding manure:a case study in the Yellow River Irrigation Region China.Nutr Cycling Agroecosyst,94(2-3):221-235.
Zhu J H,Li X L,Christie P,Li J L.2005.Environmental implications of low nitrogen use efficiency in excessively fertilizerd hot pepper(Capsicum frutescens L.)cropping systems.Agr Ecosyst Environ,111:70-80.
Zotarelli L,Scholberg J M,Dukes M D.2009a.Tomato yield,biomass accumulation,root distribution and irrigation water use efficiency on a sandy soil,as affected by nitrogen rate and irrigation scheduling.Agr Water Manage,96:23-34.
Zotarelli L,Dukes M D,Scholberg J M S,Muozcarpena R M,Icerman J.2009b.Tomato nitrogen accumulation and fertilizer use efficiency on a sandy soil,as affected by nitrogen rate and irrigation scheduling.Agr Water Manage,96:1247-1258.
Abstract:The environment pollution of nitrogen fertilization in greenhouse vegetable production is a serious problem in China.It is of great significance to realize efficient utilization of water and fertilizer resources and improve yield at the same time.The low nutrient absorptivity of root system is an important reason hinderinghigh-yielding and high-efficiency of greenhouse tomato production.This paper reviewed the latest research progress in root system architecture,discussed the characteristics of tomato root system distribution in tomato production,and the morphological response of tomato root system to controlled-release of nitrogen fertilize,analyzed the effects of combining controlled-release urea and traditional fertilizer on root system growth and distribution,put forward an ideal root system construction by applying CRU and traditional fertilizers to modify tomato root system development with dense root in top-layer and deep rooting.On this basis,the plant population structure above earth was regulated to construct an efficient root-layer continuum.Thus,in intensive tomatoes production,the nitrogen uptake of root system was increased,the leaching losses of nitrogen were avoid,and high tomato yield was obtained even with a reduced nitrogen application rate.
Key words:Root architecture;Controlled-release urea;Nitrogen uptake;Nitrate leaching
Response of Tomato Root System to Controlled Release Nitrogen and Its Ideal Type Construction
YANG Jun-gang1,2,LIAO Shang-qiang1,2,SUN Yan-xin1,2,LI Yan-mei1,2,ZOU Guo-yuan1,2
〔1Institute of Plant Nutrition and Resource,Beijing Academy of Agricultural and Forest Science,Key Laboratory of Urban Agriculture(North),Ministry of Agriculture,Beijing 100097,China;2Beijing Engineering Technology Research Center for Slow,Controlled-release Fertilizer,Beijing 100097,China〕
杨俊刚,副研究员,主要从事植物营养与新型肥料应用研究,E-mail:jungangyang@163.com
2017-06-12;接受日期:2017-07-25
国家重点研发计划项目(2016YFD0201010,2017YFD0800400),北京市重点科技计划项目(D161100005516003),北京市农林科学院科技创新能力建设项目(KJCX20151407),北京市农林科学院青年基金项目(QNJJ201611)
